基因工程菌Escherichia coli BL21/pET-pel重组果胶酶的纯化及酶学性质
2014-02-08姚晓静付大伟
徐 伟,姚晓静,付大伟
(哈尔滨商业大学食品工程学院,食品科学与工程重点实验室,黑龙江 哈尔滨 150076)
基因工程菌Escherichia coli BL21/pET-pel重组果胶酶的纯化及酶学性质
徐 伟,姚晓静,付大伟
(哈尔滨商业大学食品工程学院,食品科学与工程重点实验室,黑龙江 哈尔滨 150076)
采用超声波破碎法、镍离子亲合层析柱法,纯化重组菌Escherichia coli BL21/pET-pel表达的胞内重组果胶酶,并用十二烷基硫酸钠-聚丙烯酰胺凝胶电泳(sodium dodecyl sulfate-polyacrylamide pelelectrophoresis,SDSPAGE)检测,研究纯化的重组果胶酶酶学性质。结果表明:重组果胶酶纯化倍数为1.71,回收率为88.5%,分子质量约为44 kD。最适pH值为9.0~9.5,在pH 7.0~10.0之间催化性质较稳定;最适作用温度范围为45~55 ℃,在60 ℃下保温30 min还剩余40%的酶活力;在离子浓度为1 mmol/L时,Ca2+和Co2+对该酶有较强的促进作用,而Fe2+则对其有较强的抑制作用。
重组果胶酶;胞内酶;镍离子亲合层析;酶学性质
果胶酶是一类能够催化果胶质降解的复合酶。酸性果胶酶应用在果汁处理、果酒发酵等工业中,可有效促进饮品澄清和提高果汁出汁率[1-2]。碱性果胶酶则可以应用到造纸业、咖啡和茶发酵、香精和植物油萃取、工业废水处理以及纺织等工业中,代替传统脱胶工艺,既可以提高产品质量,又可以减少工业生产对环境的污染,为实现环境友好型工业做出贡献[3-5]。目前研究较多的微生物果胶酶为胞外酶,分离纯化时常先用硫酸铵沉淀,再用等电聚焦或离子交换层析、凝胶过滤、亲和层析等方法进行纯化;而胞内酶则需要利用超声波、酶解、去垢剂、研磨等方法破碎细胞,先释放出酶,再进行纯化[6-7],方法繁琐。通过基因工程方法则可获得带有6-His标签的重组果胶酶,结合亲合层析法可简单有效地将其分离出来[8-9]。本实验室前期从1 株芽孢杆菌中成功扩增出一段碱性果胶裂解酶基因,并在大肠杆菌E.coli BL21中成功表达,得到重组菌E.coli BL21/pET-pel。本实验对工程菌所表达的胞内碱性果胶酶进行纯化,研究pH值、温度对酶活力和稳定性的影响,以及金属离子对酶的激活或抑制作用,进一步了解重组果胶酶的酶学性质,为其在工业生产上的应用提供参考。
1 材料与方法
1.1 菌种、培养基与试剂
重组菌E.coli BL21/pET-pel为本实验前期构建并保存。
LB(luria-bertani)液体培养基:蛋白胨10 g/L,酵母粉5 g/L,葡萄糖10 g/L。
果胶 天津市东丽区天大化学试剂厂;Trisbase、十二烷基硫酸钠(sodium dodecyl sulfate,SDS) 上海榕悦生物科技有限公司;氨苄青霉素(ampicillin,Amp)、异丙基硫代半乳糖苷(isopropylβ-D-thiogalactopyranoside,IPTG) 天津博美科生物技术有限公司;蛋白胨、酵母浸粉 北京奥博星生物技术有限责任公司;中分子质量蛋白Marker 海基生物科技有限公司;N,N,N’,N’-四甲基乙二胺(N,N,N’,N’-tetramethylethylenediamine,TEMED)、丙烯酰胺、甲叉双丙烯酞胺、考马斯亮蓝 北京博奥拓达科技有限公司;镍离子亲和层析柱填料 上海锐谷生物技术有限公司;其他试剂均为分析纯。
PHS-5CpH计 杭州奥立龙仪器有限公司;V-5000紫外可见分光光度计 上海精科仪器销售公司;H1650-W高速离心机 湖南湘仪有限公司;DYCZ-24D双垂直电泳槽、BG-Power 300电泳仪 北京六一仪器厂;DHG-9123A凝胶成像系统 美国UVP公司产品;LDZX 50KB高压蒸汽灭菌锅 上海申安医疗器械厂;SW-CJ-1FD无菌操作台 苏州净化设备有限公司;SY-2-4电热式恒温水浴锅、HNY-100B恒温培养振荡器 天津市欧诺仪器仪表有限公司。
1.3 方法
1.3.1 菌种的活化与扩大培养
取本实验室甘油冷冻保藏的大肠杆菌E.coli BL21/ pET-pel菌液50 μL,接种到含LB培养基的试管中,加入100 mg/mL的Amp 5 μL,在37 ℃、150 r/min的条件下培养16 h,作为种子液备用。
以1%的接种量,将种子液转接到100 mL(250 mL三角瓶)Amp-LB液体培养基中,37 ℃、150 r/min振荡培养,至菌液OD600nm达到0.6~0.8,加入100 μL 500 mmol/L的IPTG,37 ℃、150 r/min再次振荡培养6 h,得到发酵液,冷藏备用。
1.3.2 粗酶液制备
发酵液于12 000 r/min离心5 min,弃去上清液,收集菌体,将菌体重悬于体积为发酵液10%的pH 7.5、20 mmol/L Tris-HCl中,用超声波破碎仪破碎菌体,破碎功率200 W,破碎时间10 min(超声波5 s,间隔10 s),12 000 r/min离心5 min,收集上清液,即为粗酶液。
李树化的这一首《如此温柔》在中国早期钢琴曲中是较为特别的一首,特别在于它的私密性内容和情感。这是他敬献给爱妻的一首“情歌”,既有西方浪漫主义情调,也兼有中国传统的“执子之手,与子偕老”的意愿。虽然是谱写在“圣诞节”期间,但是音乐并没有明显的宗教情绪,而是非常人性化和世俗化的表达。正是由于它的这个特点,所以李树化并没有将其公开发表,而是深深地收藏在自己的手稿中。西方浪漫主义音乐(尤其是钢琴音乐)中大都隐含着人类深沉而私密的情感,高尚的人文主义因素深蕴其中,这正是可贵之处。
1.3.3 重组果胶酶纯化
E.coli BL21/pET-pel表达的重组蛋白带有6-His标签,采用Ni2+亲合层析柱纯化[10-12]。用pH 7.8的20 mmol/L磷酸盐缓冲溶液平衡Ni2+柱,上样3 倍柱体积的粗酶液,用pH 7.8的10 mmol/L咪唑溶液洗涤,去除杂蛋白,再用pH 7.8的200 mmol/L咪唑溶液洗脱,得到目的蛋白,收集流出液冷藏备用。
1.3.4 重组果胶酶的SDS-聚丙烯酰胺凝胶电泳(polyacrylamide pelelectrophoresi,PAGE)分析
蛋白电泳胶为12%的分离胶和5%的浓缩胶,上样液为镍柱纯化的果胶酶液、粗酶液和未加入诱导剂时的E.coli BL21/pET-pel细胞裂解液,以标准分子质量蛋白作为对照,考马斯亮蓝R-250染色。
1.3.5 重组果胶酶酶活力测定
采用3,5-二硝基水杨酸法,即DNS法测定[13-14]。在试管中加入900 μL 2.5 g/L果胶底物,100 μL酶液,于50 ℃水浴10 min,加入1.5 mL DNS终止反应,在100 ℃沸水中加热5 min,冷却后于540 nm波长处测定OD值。用100 ℃加热5 min灭活的酶液作为空白对照组。结果与葡萄糖标准曲线比对,得出酶活力计算公式。规定在上述条件下,每1 min催化果胶生成1 μmol半乳糖醛酸为一个酶活力单位(U);单位质量(mg)的蛋白质所具有的酶活力单位数为比活力(U/mg)。

式中:t为反应时间/min;ODi540nm为平行样光密度值;OD540nm对照样光密度值;k为标准曲线斜率;180为葡萄糖的摩尔质量/(g/mol)。
1.3.6 蛋白含量测定
采用Bradford法,即用考马斯亮蓝G-250试剂,以牛血清白蛋白(albumin from bovine serum,BSA)为标准,测定蛋白含量。
1.3.7 重组果胶酶的酶学性质分析
1.3.7.1 重组果胶酶作用的最适pH值及其pH稳定性分析
在50 ℃下测定重组果胶酶在pH 7.0、8.0、9.0、9.5、10.0、11.0、12.0时的酶活性,确定其最适pH值;将该酶分别在pH 2.0、3.0、4.0、5.0、6.0、7.0、8.0、9.0、10.0、11.0、12.0的条件下37 ℃水浴1 h,在pH 10.0的反应条件下测定酶活性,分析其pH值稳定性。
1.3.7.2 重组果胶酶作用的最适温度和热稳定性分析
在重组果胶酶作用的最适pH值下,分别测定其在30、35、40、45、50、55、60、65、70、75、80 ℃条件下的酶活性;并在50、60、70、80 ℃的温度下将重组果胶酶保温5、10、20、30 min 后,于50 ℃反应10 min测定剩余酶活性,以未保温测得的酶活力定为100%,分析其热稳定性。
1.3.7.3 金属离子对重组果胶蛋白酶作用的影响
在含有1 mmol/L不同金属离子(Mg2+、Ca2+、Fe2+、Fe3+、Co2+、Mn2+、Ni2+、Zn2+、Cu2+、Li+、Al3+)的环境中,测定重组果胶酶的酶活性,将未加金属离子的酶活力定为100%,观察金属离子对酶活力的影响。
2 结果与分析
2.1 重组果胶酶纯化结果
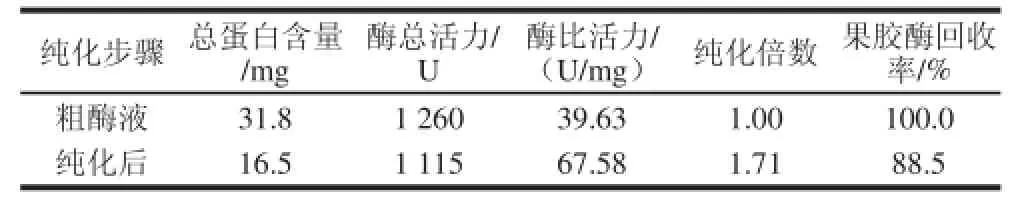
表1 重组果胶酶的纯化结果Table1 Purification of recombinant pectinase
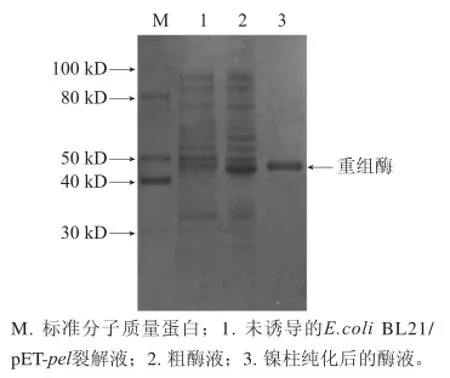
图1 重组果胶酶SDS-PAGE电泳结果Fig. 1 SDS-PAGE of recombinant pectinase
由表1可知,经镍离子亲和层析柱纯化后,酶比活力由39.63 U/mg 提高到67.58 U/mg,纯化1.71倍,回收率达88.5%。SDS-PAGE 分析如图1所示,E.coli BL21/pET-pel裂解液中蛋白条带较多,加入诱导剂后培养6 h的较未加入诱导剂的细胞裂解液在大约44 kD处出现蛋白条带,纯化后蛋白条带单一。与标准蛋白对比可知,该重组酶分子质量约为44 kD,与经Vecter NTI Explorer软件对重组果胶酶氨基酸序列进行预测分子质量相符,可确定所得纯化蛋白为重组果胶酶,且达到电泳纯。
2.2 pH值对重组果胶酶活性的影响

图2 pH值对重组果胶酶活力的影响Fig.2 Effect of pH on the activity of recombinant pectinase

图3 重组果胶酶的pH值稳定性Fig.3 pH stability of recombinant pectinase
酶催化需要其适宜的pH值环境,由图2可知,此重组果胶酶的最适pH值范围为9.0~9.5;由图3可知,在pH 7.0~10.0之间较稳定,剩余相对酶活力可保持在90%以上,而在pH 5.0以下,其稳定性迅速下降,尤其在pH 2.0、3.0时该酶几乎失去活性。
2.3 温度对重组果胶酶活性的影响
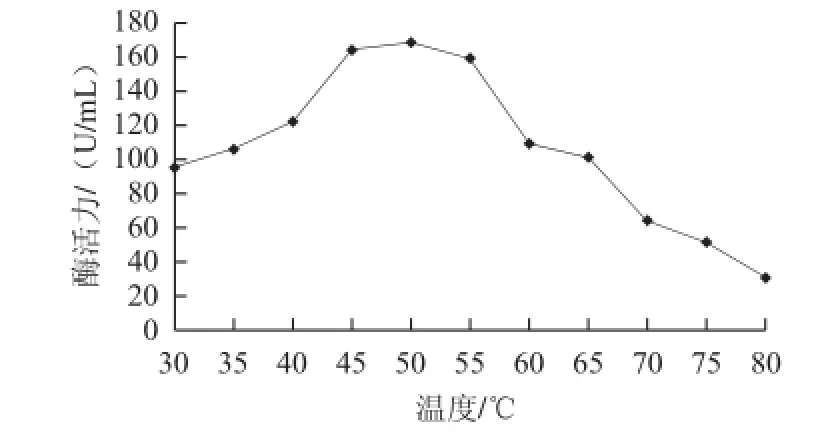
图4 温度对重组果胶酶活力的影响Fig.4 Effect of temperature on the activity of recombinant pectinase
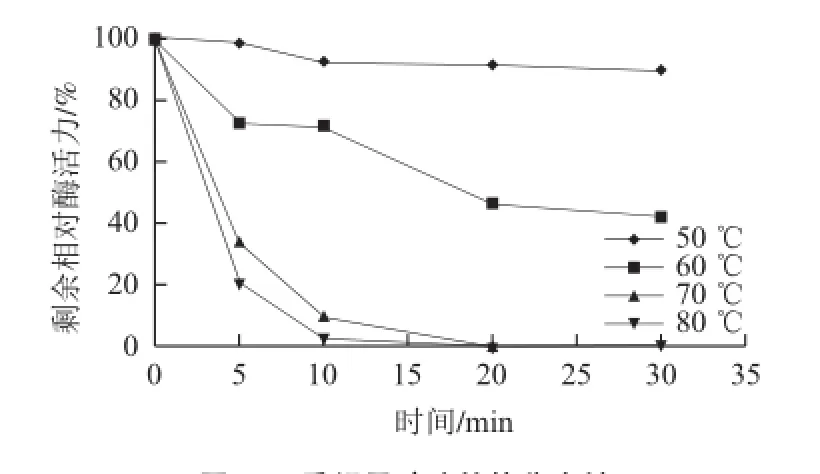
图5 重组果胶酶的热稳定性Fig.5 Thermostability of recombinant pectinase
由图4可知,该酶的最适温度为50 ℃,此时酶活力可达到168.77 U/mL,最适温度范围在45~55 ℃之间,酶活力保持在160 U/mL以上,65 ℃及以上酶活性迅速下降;由图5可知,在50 ℃下保温30 min,重组果胶酶酶活性基本不变,在60 ℃保温30 min酶活力还可保持40%以上,在70、80 ℃保温30 min,酶基本失活。
2.4 金属离子对重组果胶酶活性的影响
由表2可知,在1 mmol/L 离子浓度条件下,Zn2+、Ni2+、Ca2+、Co2+和Al3+均可提高果胶酶的活性,其中Ca2+和Co2+对果胶酶的促进作用较强,分别提高66%和74%,Zn2+、Al3+和Ni2+对果胶酶的促进程度较小,分别提高16%、19%和14%。Fe3+、Fe2+、Mn2+和Cu2+对重组果胶酶活性有不同程度的抑制作用,Fe3+、Mn2+和Cu2+的抑制作用为10%左右,而Fe2+对酶有较强的抑制作用,可使重组果胶酶丧失47%的活力。Mg2+和Li+对果胶酶活性基本没有影响。

表2 金属离子对重组果胶酶活性的影响Table2 Effects of various metal ions on the activity of recombinant pectinase
3 结论与讨论
本实验用镍柱纯化重组菌胞内的果胶酶,得到电泳纯的重组果胶酶,回收率为88%,纯化倍数可达1.71;SDS-PAGE检测表明,重组酶分子质量约为44 kD,与相关文献[15-16]报道的重组果胶酶分子质量大小在38~46 kD之间相符。
已报道的碱性果胶酶最适pH值为8.0~l0.0,最适温度范围在30~80 ℃,多数为50~60 ℃[17],不同来源的果胶酶酶学性质有一定差异。Li Zuming等[18]报道的碱性果胶酶最适反应pH值为10.0,可在pH 8.0~10.0、温度不大于40 ℃的条件下保持稳定;古丽斯玛依·艾拜都拉等[19]研究的碱性果胶酶最适作用pH值为10.0,在pH 9.0~12.0的范围内有较高活性和稳定性;Mei Yanzhen等[20]构建的工程菌表达的果胶酶最适作用pH值为10.0,在pH 9.5~10.5范围内稳定,最适作用温度为80 ℃。本研究中的重组果胶酶最适作用条件与报道的作用范围相吻合,最适反应pH值范围为9.0~9.5,条件温和,在pH 7.0~10.0之间相对稳定,范围较广泛。作用的最适温度范围为45~55 ℃,在50 ℃下保温30 min,可保持90%以上活性,热稳定性良好。
不同离子在特定条件下可以作为酶的抑制剂或催化剂。Yuan Peng等[21]报道50 μmol/L的Ca2+可以使重组果胶酶酶活提高1.8 倍,Cr3+、Pb2+可抑制果胶酶活性;Li Xiaoman等[22]研究的重组果胶酶在0.5 mmol/L Ca2+的促进下酶活可提高7 倍,Ba2+可以使其严重失活;刘丽萍等[23]研究的果胶裂解酶在Ca2+浓度为0.75 mmol/L时酶活力最高;张菊等[24]也报道Ca2+为大多数果胶酶的激活剂,Co2+和Fe2+等可抑制果胶酶活性。本研究中,在1 mmol/L离子浓度的条件下,Ca2+和Co2+使重组果胶酶酶活力提高66%~74%,可作为促进剂使用,Zn2+、Al3+和Ni2+对果胶酶的促进作用较小,Fe3+、Fe2+、Mn2+和Cu2+对重组果胶酶活力有不同程度的抑制作用,其中Fe2+对该酶有较强的抑制作用。Co2+对重组果胶酶激活作用未见报道,可能由于此酶的活性中心具有区别于其他果胶酶的基团,与Co2+作用发生了可促进酶催化的变构,从而使酶活力提高。
通过对基因工程菌E.coli BL21/pET-pel重组果胶酶酶学性质分析,该酶作用条件温和,并且在该条件下性质稳定。今后再从基因工程上进一步研究,以提高该碱性果胶酶基因的表达量,以期该酶可以工业化应用。
[1] PRATHYUSHA K, SUNEETHA V. Bacterial pectinases and their potent biotechnological application in fruit processing/juice production industry: a review[J]. Journal of Phytology, 2011, 3(6): 16-19.
[2] 郭育齐, 赵良忠, 尹乐斌, 等. 微生物果胶酶研究及其在食品加工中的应用进展[J]. 邵阳学院学报: 自然科学版, 2013, 10(2): 68-73.
[3] 李祖明, 何立千, 李鸿玉, 等. 碱性果胶酶的应用进展[J]. 食品科技, 2007, 32(8): 1-4.
[4] KASHYAP D R, VOHRA P K, S CHOPRA, et al. Applications of pectinases in the commercial sector: a review[J]. Bioresource Technology, 2001, 77(3): 215-227.
[5] ARUNACHALAM C, ASHA S. Pectinolytic enzyme-a review of new studies[J/OL]. Advanced Biotechnology Journal-Online, 2010, 561: 1-5. http://www.advancedbiotech.in/online%20article%20 Pectinolytic%20Enzyme.pdf.
[6] 贾月, 弓爱君, 邱丽娜, 等. 果胶酶分离纯化及分析方法的研究进展[J].工业微生物, 2005, 35(1): 55-58.
[7] 李祖明, 张洪勋, 白志辉, 等. 微生物果胶酶研究进展[J]. 生物技术通报, 2010, 80(3): 42-49.
[8] 肖静, 路福平, 杨晓杰, 等. Bacillus subtilis碱性果胶酶基因在大肠杆菌中的克隆和表达[J]. 高师理科学刊, 2011, 31(4): 57-59.
[9] 杨欣伟. 黑曲霉EIM-6果胶裂解酶基因的克隆与表达[D]. 福州: 福建师范大学, 2011.
[10] 王水兴, 龚珩, 郭勇, 等. 来自基因工程菌E.coli BL21/pET-DsbAMalQ的麦芽糖转糖基酶的分离纯化及酶学性质研究[J]. 食品科学, 2007, 28(12): 258-262.
[11] 夏海锋, 张显, 金雄华, 等. 镍离子亲和层析介质的制备及其用于组氨酸标记蛋白质的纯化[J]. 江南大学学报: 自然科学版, 2010, 9(6): 685-689.
[12] 张培因, 王燕媚, 卫红飞, 等. 金属鳌合亲和层析填料的制备及其在重组六聚组氨酸融合蛋白中的应用[J]. 吉林大学学报: 医学版, 2008, 34(2): 340-342.
[13] QURESHI A S, BHUTTO M A, CHISTI Y, et al. Production of pectinase by Bacillus subtilis EFRL 01 in a date syrup medium[J]. African Journal of Biotechnology, 2012, 11(62): 12563-12570.
[14] XIAO Zhizhuang, BOYD J, GROSSE S, et al. Mining Xanthomonas and Streptomyces genomes for new pectinase-encoding sequences and their heterologous expression in Escherichia coli[J]. Applied Microbiology Biotechnology, 2008, 78(6): 973-981.
[15] 房淑颖. 来源于枯草芽孢杆菌碱性果胶酶基因工程菌的构建与发酵优化[D]. 无锡: 江南大学, 2012.
[16] 张成杰. Bacillus subtilis 168碱性果胶酶基因在毕赤酵母中的优化、表达、特性以及应用的研究[D]. 武汉: 湖北大学, 2012.
[17] PEDROLLI D B, MONTEIRO A C, GOMES E, et al. Pectin and pectinases: production, characterization and industrial application of microbial pectinolytic enzymes[J]. The Open Biotechnology Journal, 2009, 7(3): 9-18.
[18] LI Zuming, BAI Zhihui, ZHANG Baoguo, et al. Purification and characterization of alkaline pectin lyase from a newly isolated Bacillus clausii and its application in elicitation of plant disease resistance[J]. Applied Biochemistry Biotechnology, 2012, 167(8): 2241-2256.
[19] 古丽斯玛依 艾拜都拉, 艾尼江 尔斯满, 木合塔 阿不都克里木, 等.碱性果胶酶产生菌Bacillus claussi XJU-8的分离鉴定及酶学特性分析[J]. 食品科学, 2011, 32(5): 182-186.
[20] MEI Yanzhen, CHEN Yuru, ZHAI Ruying, et al. Cloning, purification and biochemical properties of a thermostable pectinase from Bacillus halodurans M29[J]. Journal of Molecular Catalysis B: Enzymatic, 2013, 94(6): 77-81.
[21] YUAN Peng, MENG Kun, WANG Yaru, et al. Abundance and genetic diversity of microbial polygalacturonase and pectate lyase in the sheep rumen ecosystem[J]. PLoS One, 2012, 7(7): e40940. doi: 10.1371/ journal.pone.0040940.
[22] LI Xiaoman, WANG Huilin, ZHOU Cheng, et al. Cloning, expression and characterization of a pectate lyase from Paenibacillus sp. 0602 in recombinant Escherichia coli[J]. BMC Biotechnology, 2014, 22(10): 14-18.
[23] 刘丽萍. 基因重组果胶酶的构建及酶学性质表征[D]. 长春: 吉林大学, 2013.
[24] 张菊, 薛永常. 细菌果胶酶的研究进展[J]. 生物技术通报, 2011, 91(2): 56-60.
Purification and Characterization of Recombinant Pectinase from Genetically Engineering Strain Escherichia coli BL21/pET-pel
XU Wei, YAO Xiao-jing, FU Da-wei
(Key Laboratory of Food Science and Engineering, School of Food Engineering, Harbin University of Commerce, Harbin 150076, China)
The recombinant pectinase expressed in Escherichia coli BL21/pET-pel was purified by ultrasonic cell disruption and nickel ion affinity chromatography, and detected using SDS-PAGE. Its enzymatic characteristics were analyzed. The results showed that purification fold of the recombinant pectinase was 1.71, and recovery was 88.5%. Molecular weight of the recombinant pectinase was approximately 44 kD. Its optimal temperature and pH were 45-55 ℃ and 9.0-9.5. It was stable at pH 7.0-10.0. When the recombinant pectinase was incubated for 30 min at 60 ℃, the relative activity was approximately 40%. Different metal ions revealed different effects on the recombinant pectinase, which was activated by Ca2+and Co2+but inhibited by Fe2+.
recombinant pectinase; intracellular enzyme; nickel ion affinity chromatography; enzymatic characteristic
Q815
A
1002-6630(2014)23-0245-04
10.7506/spkx1002-6630-201423047
2014-06-24
徐伟(1963—),女,教授,博士,研究方向为微生物发酵工程。E-mail:xuw@hrbcu.edu.cn
