极晚熟杏品种‘冬杏’杂交亲和性研究
2014-01-22赵世荣安晓芹刘曼曼王玉蓉
赵世荣,廖 康,安晓芹,孙 琪,刘 娟,刘曼曼,王玉蓉
(新疆农业大学 特色果树研究中心,新疆 乌鲁木齐 830052)
‘冬杏’又名坎及玉吕克,是原产于新疆阿克苏的实生变异杏品种,属于中亚细亚生态群[1],在4月初始花,10月初果实开始陆续成熟,果实发育期为180~195d,为新疆杏的极晚熟特异种质资源。实际生产中由于杏成熟期过于集中,存在市场供应期及工厂加工期短的压力。亟待开展不同成熟期杏品种的选育工作。Kostina[2]发现,中亚品种和欧洲品种杂交,其后代成熟期有趋中变异趋势,Hough[3]发现杏的成熟期属母本多基因控制的数量性状遗传,‘冬杏’的极晚熟特性可用于开展晚熟品种选育工作,从而解决生产中不同成熟期杏品种的合理配置问题。开展杂交选育工作父、母本亲和是前提条件,刘立强、安晓芹等[4-6]研究表明,‘轮台白杏’在与不同品种的正、反交试验中,‘冬杏’ב轮台白杏’杂交组合坐果率仅为4%,表现为杂交不亲和,新疆杏品种间的杂交坐果率在9%~60%之间,因此有必要扩大范围开展杂交亲和性品种筛选工作,目前杂交亲和性鉴定的方法主要有田间授粉实验、花柱离体培养以及花粉原位萌发与花粉管生长荧光观察法。郑洲、冯建荣[7-8]等的研究结果显示,杂交亲和的组合在授粉12h后花粉均已萌发,72h花粉管到达花柱3/4处,并在授粉后96h进入子房。而杂交(自交)不亲和的组合,在72h花粉管到达花柱3/4处后末端膨大,停止生长。根据花粉管最后在花柱中的位置,可以初步确定果梅[9]、梨[10-11]、榛属[12]等的亲和性。经过连续2年的观察,‘冬杏’的自交坐果率较低,生产中需配置授粉树。本研究以‘冬杏’为母本,分别与15个性状良好的不同成熟期的新疆杏品种进行杂交,通过各品种的花粉量、花粉活力以及杂交组合的花粉管生长状况、坐果率来判定杂交亲和性,为选育杏的晚熟品种奠定理论基础。
1 材料与方法
1.1 材 料
试验于2012~2013年在新疆轮台县国家果树资源圃进行。共选取了16个品种,所选各品种的树龄为25 a,树势中庸,栽培管理水平一致。各品种如表1所示:
1.2 方 法
1.2.1 花粉采集及自花、异花授粉
花粉采集:于小蕾期剪取各品种发育充实的花枝,在室温25℃下水培。至大蕾期采集各品种饱满的花蕾,分离出花药,自然散粉后收集于1.5mL离心管中贮存于-20℃冰箱内,用于授粉试验及花粉活力测定。

表1 杂交所选16个不同成熟期的杏品种Table 1 16 apricot cultivars with different maturation periods in the pollination experiment
花粉量测定:从各品种采集10朵饱满的气球期花蕾,分离出单花花药,每花蕾各取完整的10枚花药置于1.5mL离心管中自然散粉。用悬浮离心法测定其花药的花粉含量[5]。
花粉活力测定:配制0.01%硼酸+10%蔗糖+1%琼脂的培养基,将采集的花粉均匀弹布在倒有培养基的载玻片上,25℃恒温培养4h,用显微镜(Motic BA400 TYPE 102M型)镜检,统计视野中的花粉总数和萌发花粉数,重复5个视野。
花粉萌发率/%=(视野中已萌发花粉数÷视野中总花粉数)×100。
自花授粉:待‘冬杏’花蕾至大蕾期时,去雄授自花花粉,套袋,第一次生理落果结束后统计坐果率。
异花授粉:待‘冬杏’花蕾至大蕾期时,去雄后分别授以15个不同成熟期品种的花粉,套袋,第一次生理落果结束后统计坐果率。
1.2.2 花粉原位萌发及花粉管伸长的荧光显微观察
剪取发育充实且粗细、长短一致的小蕾期‘冬杏’花枝置于室内水培,室温25℃左右,大蕾期去雄,授以自花花粉。剪取‘冬杏’发育充实、粗细和长短相似的大蕾期花枝数枝,室内25℃水培。去雄后分别授以所选15个不同成熟期品种的花粉。各授粉处理后的水培花枝分别于授粉后0.5、1、2、4、6、8、12、24、48、72h和96h时取样,每处理10枚花柱重复3次。样品用FAA固定液(甲醛∶冰醋酸∶70%酒精=5∶5∶90)固定后,带回实验室。
花柱观察前处理参考Kho、杨志、杜纪红等[13-15]的方法,将固定材料经浓度分别为70%、50%、30%的酒精清洗,过渡到蒸馏水中,然后用8mol/L NaOH溶液软化处理,再用0.1%的苯胺蓝染色液染色。经染色处理后的花柱做成压片,在荧光显微镜(Nikon ECLIPSE 80i)下观察(荧光激发采用EX 380~420nm,DM 430nm,BA 450nm),分别观察柱头上花粉的萌发及花柱内花粉管生长状况,并进行显微拍照。统计每一时段有花粉萌发的花柱比率、花粉管到达花柱1/3、1/2与花柱基部的花柱比率。
1.3 数据统计及分析
统计各品种的花粉量、花粉活力、坐果率和每一时段有花粉萌发的花柱比率、花粉管到达花柱1/3、1/2与花柱基部的花柱比率。数据采用Excel和SPSS 19.0进行数据统计和方差分析。
2 结果与分析
2.1 不同授粉品种单花药花粉量和花粉活力比较
2.1.1 花粉量比较
测定不同品种的花粉量并进行方差分析,结果如表2所示。16个杏品种的花粉量差异达极显著水平。花粉平均含量为1 413.24粒,有7个品种高于平均值,多数品种在1 000~2 500粒之间。其中以‘特尔湾玉吕克’最高,为2 508.8粒/花药。‘轮台白杏’最低,为405.1粒/花药。
2.1.2 花粉活力比较
测定不同品种的花粉活力并进行了方差分析,结果如表2所示。16个品种花粉活力差异极显著,平均值为51.0%。以‘贾格达玛依桑’最高,达79.8%;‘库车托永’最低,仅为20.1%,多数品种的花粉活力在40%~80%之间,其中有8个品种的花粉活力高于平均值。
2.2 不同父本田间授粉坐果率比较
以‘冬杏’为母本,授以其它所选品种花粉后统计其坐果率,结果如表3所示。不同父本授粉后坐果率差异较大,平均坐果率为22.5%,以‘贾格达玛依桑’为最高,达36.6%,‘旦杏’最低,仅为16.4%。所有杂交组合的坐果率均显著高于自然坐果率(10.7%)。‘冬杏’自交的坐果率最低,仅为1.5%。
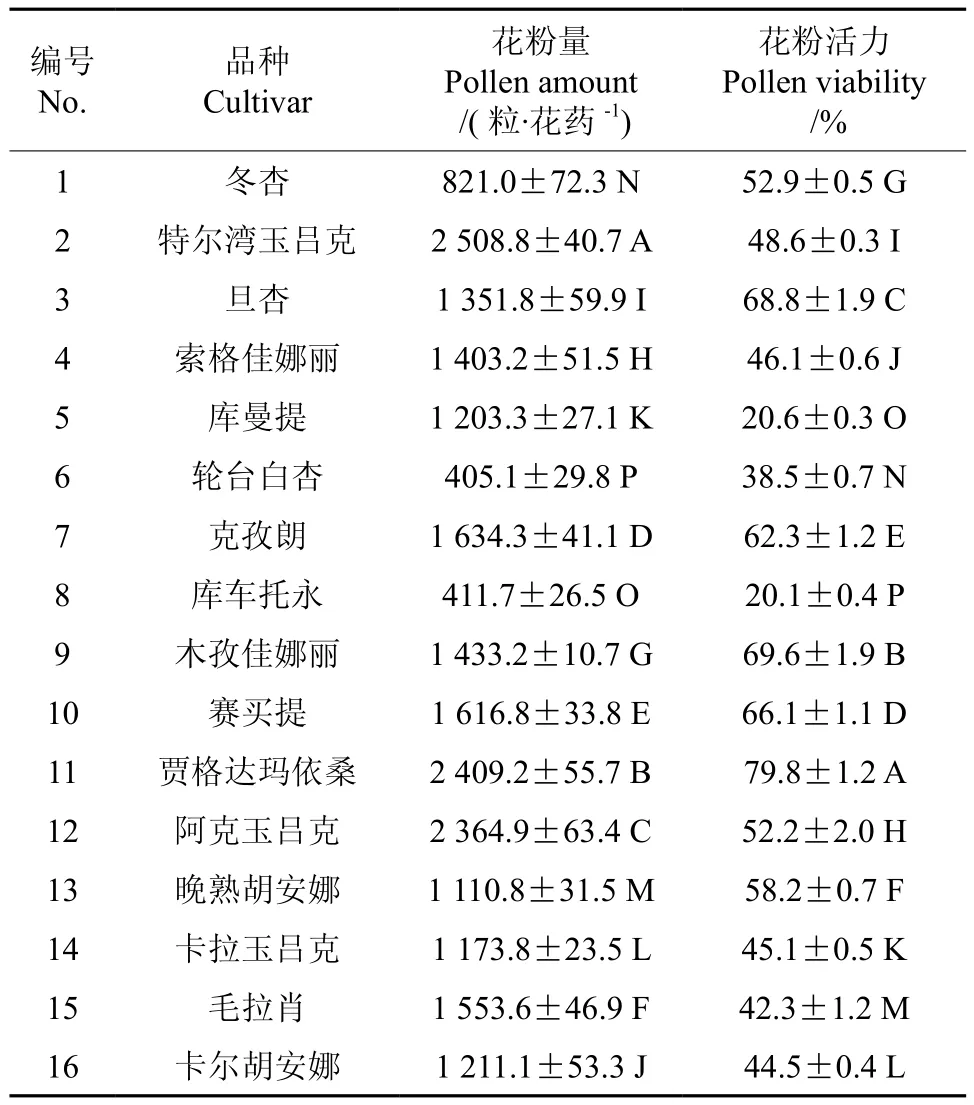
表2 不同杏品种单花花粉量及花粉活力†Table 2 Pollen amount and viability of different apricot cultivars
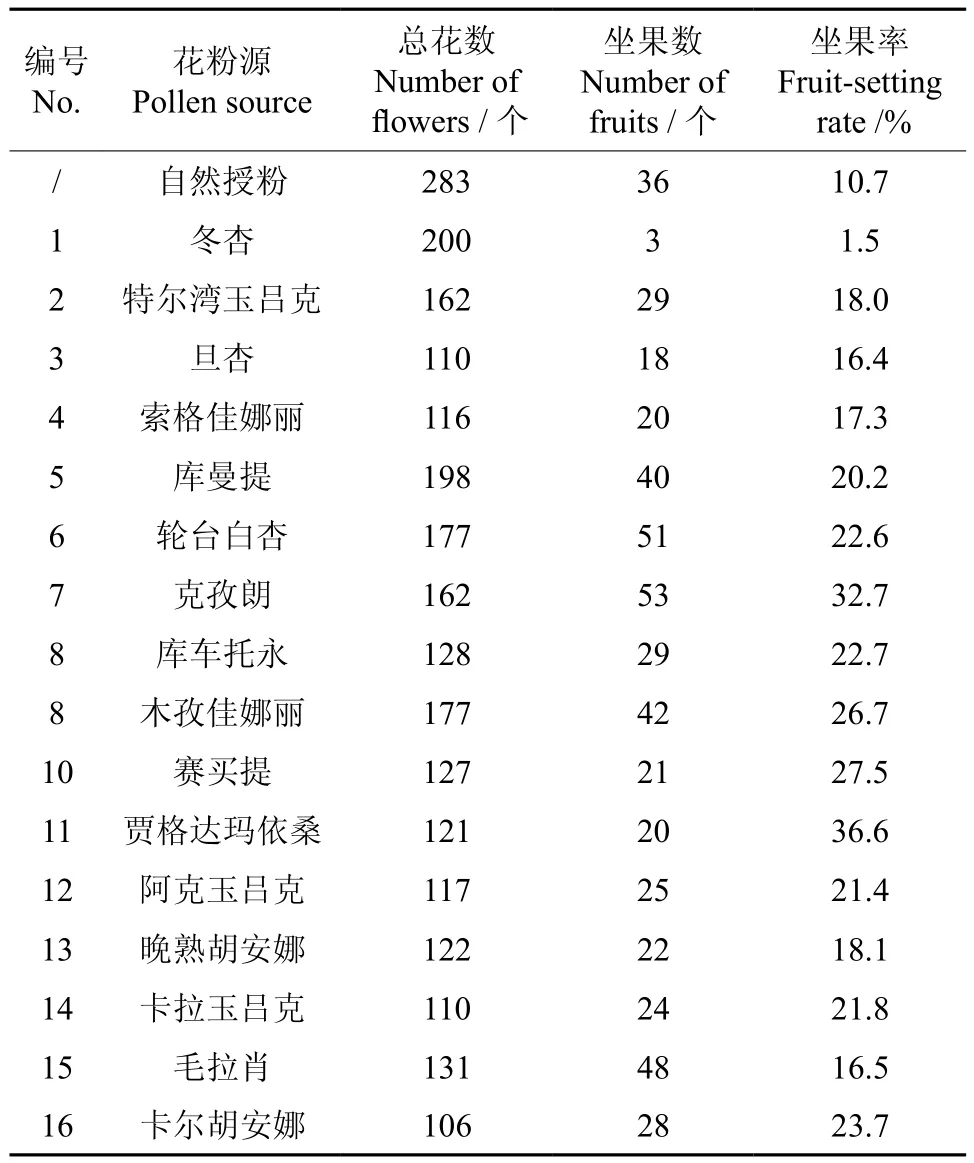
表3 不同父本授粉的坐果率比较Table 3 Comparison of fruit-setting rates after pollinated by different male parents
2.3 花粉原位萌发和花粉管伸长
对各父本花粉在‘冬杏’柱头上不同生长时间花粉管的伸长进程进行观察,统计结果如表4所示,图1显示了不同父本花粉在‘冬杏’柱头上的萌发和花粉管的伸长状况。其中有9个品种花粉授在‘冬杏’柱头上2h后,花粉开始萌发(图1 A)。以‘索格佳娜丽’、‘晚熟胡安娜’花粉在‘冬杏’柱头萌发的花柱最多,达到50%;‘卡拉玉吕克’花粉在授粉后6h才开始萌发,花粉在‘冬杏’柱头萌发的花柱为20%。

表4 不同父本花粉在‘冬杏’柱头上原位萌发与花粉管伸长进程Table 4 Processes of pollen germination and pollen tube growth of different male parents in ‘Dongxing’ styles
‘库曼提’、‘轮台白杏’和‘赛买提’花粉在授粉6h后,花粉管最先到达‘冬杏’花柱的1/3处(图1 B)。以‘库曼提’花粉管到达花柱1/3处的花柱最多(30%)。授粉8h大多数花粉的花粉管都到达花柱1/3处,‘索格佳娜丽’‘卡拉玉吕克’在授粉12h后才到达花柱1/3处。
‘木孜佳娜丽’、‘赛买提’、‘贾格达玛依桑’、‘毛拉肖’4个品种花粉管在授粉后12h到达‘冬杏’花柱1/2处(图1 C),花粉管到达花柱1/2处的花柱为10%。‘阿克玉吕克’在授粉后72h后花粉管才延伸至‘冬杏’花柱的1/2处,但到达的花柱较多(70%)。其余品种均在授粉后24~48h才到达。
在授粉24h后‘赛买提’花粉管最先到达‘冬杏’花柱基部(图1 D),并在48h有10%花柱中花粉管到达胚珠(图1 E)。‘阿克玉吕克’花粉管在授粉96h后最迟到达花柱基部,有花粉管到达基部的花柱为20%。其它的在48~72h到达基部。授粉96h后各品种花粉管到达花柱基部的平均比率为70%,其中以‘库曼提’、‘贾格达玛依桑’、‘卡拉玉吕克’、‘卡尔胡安娜’4个品种花粉管到达比率最高(90%),‘阿克玉吕克’最低(20%)。
‘冬杏’花粉在授粉2h后开始萌发,在授粉12h后才到达花柱1/3处,48h后到达花柱的1/2处。花粉管在授粉96h后才到达花柱基部,到达花柱比率为40%(图1 F)。
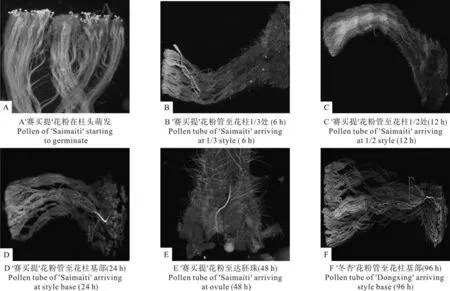
图1 花粉在‘冬杏’柱头上萌发及花粉管伸长进程Fig.1 Processes of pollen germination and pollen tube growth in ‘Dongxing’ styles
3 讨 论
不同成熟期杏品种的花粉量和花粉活力均存在较大的差异。经分析可知,二者之间并不存在明显的相关性。品种间花粉量的差异主要与其自身的遗传特性有关,其次还与树体的营养状况和花芽分化和发育状况有关[16]。在供试的16个杏品种中,除‘库车托永’外,其它品种的花粉活力均在40%以上,均适宜作为授粉品种。实验测得的各品种的花粉活力与安晓芹等[5]的测定结果相近,但‘轮台白杏’、‘克孜朗’等品种的花粉活力均高于刘立强[4]2004年在同一实验地的所测值,且与野生杏相比多数品种的花粉活力较低[6],这可能是栽培型和野生型在遗传特性、所处环境、活力测定培养基等[17-19]方面的因素不同所致。
在相似生境下,以‘冬杏’为母本的不同父本授粉后坐果率存在一定差异。分析得知,父本花粉量与坐果率之间无明显的相关性,主要是受到父本的花粉活力、花粉与柱头的亲和性等因素的影响。
不同父本花粉在母本‘冬杏’柱头上的萌发与花粉管的伸长速度和所占花柱比率,从一定程度上反映了它们之间的亲和性。从花粉管伸长的荧光显微观察结果可以看出,花粉活力较高的品种在‘冬杏’柱头原位萌发率和花粉管到达花柱基部的比率均较高。个别品种的坐果率高低与柱头荧光观察结果并不完全一致,这可能是花粉管在伸长过程和进入子房后,受到了其它因素的干扰,有待于进一步研究。‘贾格达玛依桑’、‘木孜佳娜丽’、‘赛买提’、‘克孜朗’等坐果率较高的授粉品种,其花粉活力、花粉萌发速度和萌发量、花粉管到达花柱基部的速度和比率均处于中上水平。所以花粉活力及亲和性可作为筛选坐果率较高品种的主要参考因素。
16个杏品种的花粉多数在授粉2h后开始萌发,并于授粉后96h均到达了花柱基部,这与郑洲[7]在红丰、新世纪等华北生态群杏品种上的研究结果一致。‘冬杏’自交坐果率小于6%,表现为自交不亲和[11,20-22],此结果与安晓芹等[5]的研究结果一致。冯建荣、郑洲等[7-8]试验结果显示‘凯特杏’、‘红丰’、‘新世纪’等华北生态群的自交不亲和品种花粉管不能或极少到达花柱基部,但‘冬杏’在授粉96h后仍有40%的花粉管能到达子房基部,这可能是由于实验品种的遗传特性和生境不同所致,有待于进一步开展研究。
4 结 论
16个不同成熟期杏品种的平均花粉含量为1 413.24粒/花药,平均花粉活力为51.0%,平均坐果率为22.5%。‘贾格达玛依桑’、‘木孜佳娜丽’、‘赛买提’、‘克孜朗’等品种与‘冬杏’的杂交坐果率较高,‘毛拉肖’、‘索格佳娜丽’、‘旦杏’等品种与‘冬杏’杂交坐果率相对较低。‘冬杏’自交坐果率小于6%,为自交不亲和类型。花粉多在授粉后2h后开始萌发,授粉96h后到达花柱基部,但是各品种间的差异性较大。花粉活力与花粉萌发和花粉管伸长存在一定的相关。通过坐果率、花粉原位萌发生长结果来看,‘贾格达玛依桑’、‘木孜佳娜丽’、‘赛买提’、‘克孜朗’等品种与‘冬杏’杂交亲和性较高。‘毛拉肖’、‘索格佳娜丽’、‘旦杏’等品种与‘冬杏’杂交亲和性相对较差。
[1] 张加延,张 钊.中国果树志(杏卷)[M].北京:中国林业出版社,2003:283.
[2] Kostina KF.Breeding use of apricot varietal funds[J].Proc State Nikit Bot Sad,1969,40:65-80.
[3] Hough LF,Bailey CH.30 years of apricot breeding in New Jersey[J].Acta Hort,1982,121:207-210.
[4] 刘立强,廖 康,张大海,等.新疆杏品种授粉生物学初探[J].北方园艺,2007,(9):36-38.
[5] 安晓芹,廖 康,孙慧瑛,等.不同杏品种授粉对‘轮台白杏’坐果及果实品质的影响[J].浙江农林大学学报,2013,30(2):187-193.
[6] 刁永强,廖 康,许 正,等.新疆野生杏开花生物学特性及授粉受精初步研究[J].新疆农业科学,2010,47(5):947-951.
[7] 郑 洲,陈学森,冯宝春,等.杏品种授粉生物学研究[J].果树学报,2004,21(4):324-327.
[8] 冯建荣,陈学森,孔 宁,等.杏(Prunus armeniaca)自交不亲和强度及其授粉受精相关特性[J].果树学报,2006,23(5):690-694.
[9] 陶书田,张绍铃,陈迪新,等.果梅花粉原位萌发及花粉管生长特性的研究[J].果树学报,2004,21(4):338-340.
[10] 吴 俊,徐 芹,张绍铃,等.精胺对梨花粉原位萌发及花粉管生长的影响[J].果树学报,2005,22(6):606-609.
[11] 袁德义,张 琳,段经华,等.梨树生殖生物学研究进展[J].经济林研究,2007,25(3):73-80.
[12] 翟学杰,董凤祥,张日清,等.5种榛属植物的亲和性分析[J].中南林业科技大学学报,2009,29(4):26-29.
[13] Kho YO,Bear J.Observing pollen tubes by means of fl uorescence [J].Euphytica,1968,17:298-302.
[14] 杨 志,董晓涛,杨 巍,等.果树花粉生活力应用研究概况[J].北方果树,2006(4):1-3.
[15] 杜纪红,叶正文,苏明申,等.桃花粉离体萌发和花粉管生长特性研究[J].西北植物学报,2011,31(1):0064-0071.
[16] Sedgly M,Griffin A R.Sexual reproduction of tree crops[M].Toronto: Academic Press,1989.
[17] 杜玉虎,张绍玲.梨43个品种花粉生活力及4种测定方法的比较[J].果树学报.2006,23(2):178-181.
[18] 王湘南,陈永忠,王 瑞,等.油茶花粉活力及柱头可授性研究[J].中南林业科技大学学报,2012,32(3):18-22
[19] 王湘南,陈永忠,王 瑞,等.油茶主栽品种的开花授粉习性研究[J].中南林业科技大学学报,2013,33(12):1-5
[20] Audergon J M.Contribution to the study of inheritance of the character self-incompatibility in apricot[J].Acta Hort,1999,488:275-279.
[21] 李 疆.新疆扁桃结实率低的生理与分子机制[J].经济林研究,2011,29(2):1-7.
[22] 郭素娟,吕文君,邹 峰,等.迁西板栗主栽品种授粉组合的优化[J].江西农业大学学报,2013,35(3):437-443.
