苦荞在萌发过程中营养物质的变化及其营养评价
2014-01-18周一鸣崔琳琳李保国周小理
周一鸣,崔琳琳,王 宏,李保国,周小理,*,谢 凡
(1.上海理工大学食品科学与工程研究所,上海 200093;2.上海应用技术学院香料香精技术与工程学院,上海 201418;3.上海商学院旅游与食品学院,上海 200235)
苦荞在萌发过程中营养物质的变化及其营养评价
周一鸣1,2,崔琳琳3,王 宏2,李保国1,周小理2,*,谢 凡2
(1.上海理工大学食品科学与工程研究所,上海 200093;2.上海应用技术学院香料香精技术与工程学院,上海 201418;3.上海商学院旅游与食品学院,上海 200235)
通过测定萌发过程中苦荞的蛋白质、脂肪、脂肪酸、氨基酸、总糖、还原糖、α-、β-淀粉酶活性、矿物质元素、VB1和VB2的含量,研究萌发处理对苦荞营养价值的影响。结果表明:萌发过程中,蛋白质和脂肪等大分子物质均呈下降趋势;但可溶性低分子糖类、含氮物质和脂肪酸等小分子物质含量有较大提高,其中还原糖含量上升1.61%~10.63%,总氨基酸含量增加了3.85%,且7 种必需氨基酸含量均呈现上升趋势,赖氨酸增幅最大(30.5%)。与未萌发苦荞相比,VB1、VB2含量变化不大;同时,随着萌发过程的延长,α-、β-淀粉酶活性不断增强。萌发过程中,必需氨基酸/总氨基酸比值和必需氨基酸指数增加,必需氨基酸组成模式更加符合联合国粮食及农业组织/世界卫生组织(FAO/WHO)联合食品标准计划,第一限制氨基酸由赖氨酸转变为蛋氨酸+胱氨酸。
苦荞;萌发;营养成分;营养价值评价
苦荞含有丰富的蛋白质、不饱和脂肪酸、淀粉、矿物质和维生素、黄酮类化合物等物质,其营养素种类丰富,含量均衡[1-2],但其特有的苦味影响了口感。研究[3-5]表明,由于蛋白酶抑制剂、芦丁降解酶等物质的存在,使得苦荞蛋白质和淀粉消化率低,黄酮类化合物在加工过程中迅速分解。适当的萌发处理后,可以提高蛋白质和淀粉的消化率以及某些限制性氨基酸和维生素等营养物质的含量,其化学成分均有所改变,营养价值得以提高,并可形成独特的风味及口感,有的还增加了一些药用功能成分。同时,萌发处理可以降低或消除其中有毒、有害或抗营养物质的含量。淀粉作为苦荞的主要成分,含量一般在60%~70%之间,萌发期间淀粉酶大量产生,且随着淀粉酶活力的逐渐升高,淀粉含量逐渐降低,还原糖含量则逐渐增加[6-9]。同时,在淀粉酶的作用下胚乳淀粉逐步分解,淀粉颗粒的内部结构发生变化。
本实验重点研究了苦荞萌发过程中蛋白质、氨基酸、脂肪、脂肪酸、碳水化合物和黄酮类化合物等主要营养物质以及α-、β-淀粉酶的变化情况,以掌握萌发过程中苦荞营养成分以及α-、β-淀粉酶的变化规律,为萌发苦荞新型食品开发提供理论依据。
1 材料与方法
1.1 材料与试剂
苦荞(山西黑丰1号) 山西左云县。
盐酸、高纯氮、石油醚、葡萄糖、无水乙醇、氢氧化钠、碘试剂(碘化钾-碘溶液)、丙酮、硫酸、草酸、硫脲、抗坏血酸等(均为分析纯) 国药集团上海化学试剂有限公司;牛血清白蛋白、考马斯亮蓝G-250 西格玛奥德里奇中国。
1.2 仪器与设备
RF-10A XL荧光检测器、UV2600紫外-可见分光光度计、LC-20Ap高效液相色谱仪 岛津企业管理(中国)有限公司;L-8900全自动氨基酸分析仪 日本日立公司;7890A-5975C型GC-MS联用仪 安捷伦科技有限公司;SpectrAA240FS火焰原子吸收光谱仪 美国Varian公司;WX-4000型微波快速消解系统 上海屹尧微波化学技术有限公司。
1.3 方法
1.3.1 苦荞萌发处理
筛选籽粒饱满、成熟度好的苦荞,先用2%次氯酸溶液浸泡10 min,再用0.1% H2O2中浸泡10 min杀菌,清水反复冲洗后再浸泡12 h,室温下催芽24 h播种,播种后每8 h浇水一次(浇水量以苗盘内基质湿润又不浸水为宜),每天用15 W紫外灯照射15 min,保持环境的湿度(80%~90%)与温度(25~30 ℃),播种后连续4 d取样,1/2用于酶活测定,置于-80 ℃备用,1/2除去外壳,冷冻干燥后粉碎机粉碎,80 目筛分,低温密封保存备用。
1.3.2 萌发苦荞营养成分的测定方法
蛋白质:考马斯亮蓝(G-250)比色法[10];粗脂肪:GB/T 14772—2008《粮油检验粮食中粗脂肪含量测定》中索氏抽提法;碳水化合物:DNS比色法[11];维生素:GB/T 7628—2008《谷物中维生素B1测定》,GB/T 7629—2008《谷物中维生素B2测定》;矿物元素:原子吸收法[12];α-、β-淀粉酶活性测定[13-14]。每个样品测定3次。
脂肪酸测定:准确称取萌发苦荞冻干粉2.0 g,索氏抽提10 h,浓缩并常温真空干燥至恒重。用5 mL的正己烷-乙醚(体积比2∶1)溶解样品脂肪酸,加入0.5 mol/L NaOH甲醇溶液3 mL,超声处理10 min,加入蒸馏水10 mL,摇匀,再超声处理1 min,取出移至离心管中,4 000 r/min离心3 min,吸取上层脂肪酸甲酯,作GC-MS分析。分析条件:HP-5MS毛细管柱(0.25 mm×30 m,0.25 μm),初始柱温:80 ℃,以16 ℃/min的速率升至280 ℃,流速为1 mL/min,分流进样,分流比:25∶1,进样温度285 ℃,进样量0.01 μL。MS分析条件:电离方式:EI,离子源温度:225 ℃,电离电压:60 eV。
氨基酸测定:准确称取萌发苦荞冻干粉0.3 g于试管中,加入15 mL HCl(浓度6 mol/L)后盖反口塞,真空泵上抽真空,再充N2气,重复3 次后密封,110 ℃烘箱内水解22 h后取出冷却至室温。用氢氧化钠调pH值至2.2后定容至50 mL,过0.22 μm的微孔滤膜,用全自动氨基酸分析仪进行测定。测试条件:分离柱(4.6 mm×60 mm),流速0.4 mL/min,柱温70 ℃。
1.3.3 蛋白质的营养价值指标的计算
1.3.3.1 氨基酸评分(amino acid score,AAS)

1.3.3.2 必需氨基酸(essential amino acids,EAA)与总氨基酸(total amino acids,TAA)之比(E/T)
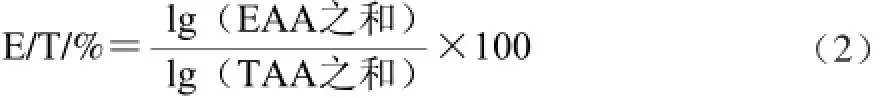
1.3.3.3 必需氨基酸指数(essentia lamino acid index,EAAI)

式中:n为计算中所涉及的必需氨基酸数;αi为样品中各种氨基酸含量/%;Ai为鸡蛋蛋白中相应的必需氨基酸含量/%;i=1,2…,n。
2 结果与分析
2.1 苦荞萌发过程中营养物质的变化
2.1.1 萌发后苦荞蛋白质含量的变化
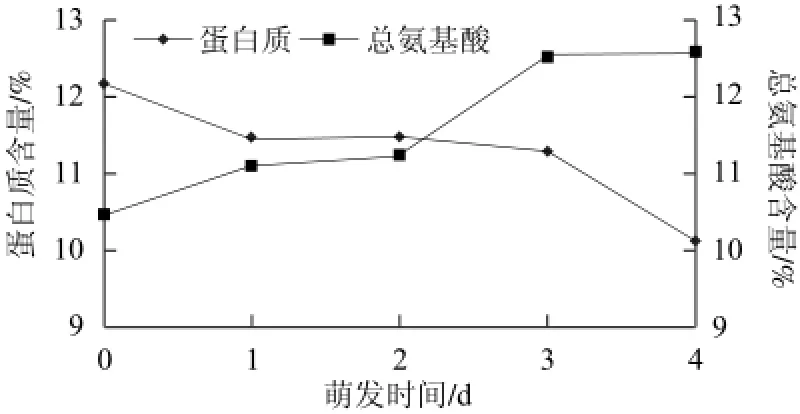
图1 萌发过程中苦荞蛋白质和总氨基酸含量的变化Fig.1 Changes in the contents of protein and total amino acids in tartary buckwheat during germination
由图1可知,苦荞在萌发过程中会表达不同的基因,分化为各种细胞,在整个分化的前期,也就是在没有能力合成蛋白质之前,苦荞内蛋白质储量成为此段时期蛋白的唯一来源,会大量的消耗蛋白质用于分化,与此同时,自身并没有能力大量、稳定地合成蛋白质,表现为随着萌发时间的增加,苦荞蛋白质的含量也随之降低,但在萌发后期,在蛋白酶的作用下苦荞中其他含氮物质被分解为小分子的肽类和氨基酸,这些分解产物分泌到胚芽,合成新的蛋白质,由此蛋白质减低趋势随天数放缓。
同时,与未萌发苦荞相比,萌发后苦荞总氨基酸含量呈显著上升趋势,与蛋白质变化曲线比较,在第2天时,蛋白质的降解及其氨基酸的合成出现平衡。

图2 萌发过程中苦荞粗脂肪的含量变化Fig.2 Changes in fat content in buckwheat during germination
2.1.2 萌发后粗脂肪含量的变化由图2可知,随着苦荞籽粒的萌发,由于初期光合作用的微弱,碳源不足,脂肪类物质被消耗作为能源或是分解成小分子作为其新芽的成分,此时含量呈明显下降趋势,从未萌发的92.5 mg/g下降至萌发4 d的54.6 mg/g,下降了41%。
2.1.3 萌发后维生素含量的变化
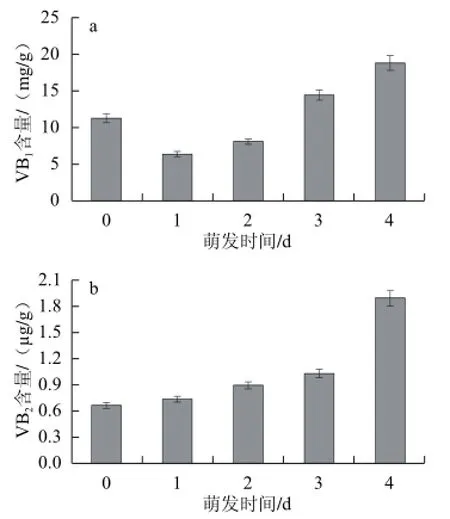
图3 苦荞萌发过程中VVBB1、VVBB2的含量变化Fig.3 Changes in the contents of VB1and VB2in buckwheat during germination
由图3可知,苦荞萌发过程中水溶性维生素含量总体上是随着萌发天数的增加而增加,与未萌发样品比较,VB1、VB2分别提高了67.3%、190.7%。在萌发初期,由于营养供给的消耗都会有一个下降的过程,但随着萌发的继续,作为植株内生长发育所需的辅酶,分化的细胞开始表达维生素,故含量稳步上升。从含量上来看,VB1的含量在苦荞中比例最大。
2.1.4 萌发后α-、β-淀粉酶活性的变化
α-淀粉酶是胚乳淀粉粒水解中的起始酶。在植物种子萌发过程中,随着温湿度的增加,胚芽中合成植物激素赤霉酸从而诱导α-淀粉酶从头开始合成[15]。在苦荞萌发过程中α-淀粉酶活性变化如图4所示,α-淀粉酶是在萌发开始后重新合成的酶,与之前Jones等[15]的研究结果一致,α-淀粉酶不断合成并逐渐积累,萌发初期(0~2 d),α-淀粉酶活性增加迅速。萌发3 d后,α-淀粉酶的合成速度变慢,变化进入一个相对稳定的阶段。β-淀粉酶的合成机制与α-淀粉酶不同,所有的 β-淀粉酶都不是从头合成的[16]。成熟的植物籽粒中有游离态和束缚态的两种形态的β-淀粉酶,其中束缚态的 β-淀粉酶占总β-淀粉酶的 75%,β-淀粉酶是在籽粒成熟期间合成和积累,萌发时没有进一步合成,束缚态β-淀粉酶在籽粒萌发过程中逐渐释放[17]。从图4可以看出,萌发过程中,β-淀粉酶的变化趋势和 α-淀粉酶相似,未萌发的苦荞籽粒中β-淀粉酶具有相对较低的活性,萌发开始后到2 d这个阶段 β-淀粉酶的活性增加相对较快,在2~4 d这个阶段,β-淀粉酶活性继续增加,但变化进入一个相对稳定的阶段。
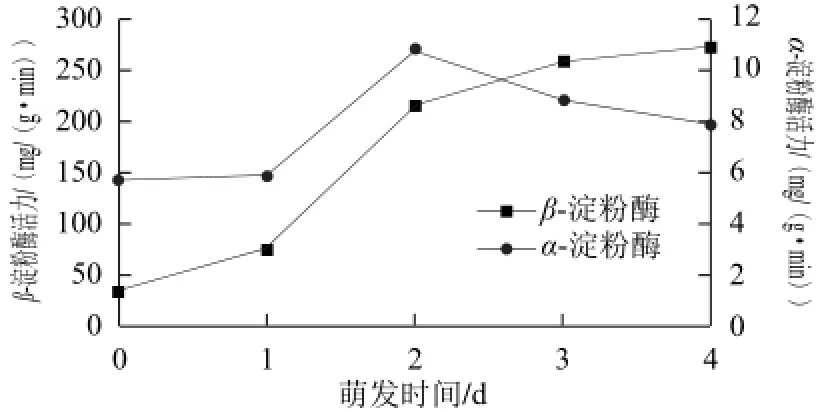
图4 苦荞萌发过程中α--、β-淀粉酶活性的变化Fig.4 Changes in α and β-amylase activities in tartary buckwheat during germination
2.1.5 萌发后碳水化合物含量的变化
苦荞籽粒中大部分糖类是作为营养源的,含量接近90%。由图5可知,在萌发初期(0~1 d)总糖含量增加,说明苦荞籽粒在萌发初期生理过程比较复杂,同时存在蛋白质、脂类和糖类之间的转化。在萌发后期(2~4 d)随着植株生长分化所需,淀粉在淀粉酶的作用下发生降解,生成较小分子的单糖和低聚糖,籽粒萌发的呼吸作用消耗和水溶性糖类的溶解导致总糖含量的减少。
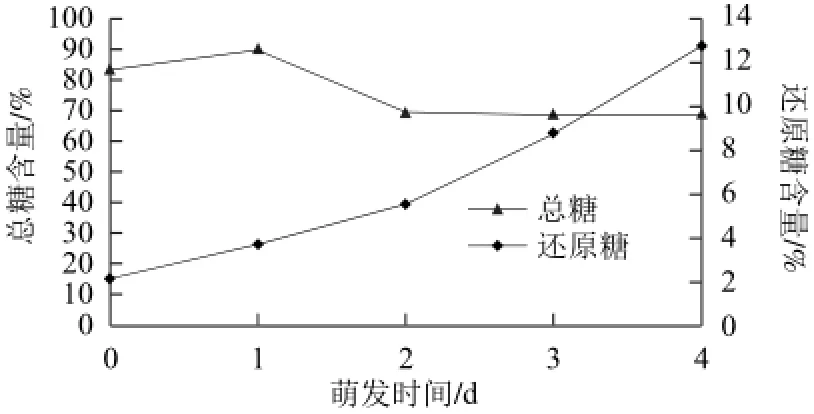
图5 萌发过程中总糖和还原糖的含量变化Fig.5 Changes in the contents of total sugar and reducing sugar during buckwheat germination
还原糖含量随萌发时间的变化曲线如图5所示,苦荞籽粒在萌发过程中随着萌发时间的延长,还原糖的含量逐渐增大。这是由于萌发过程中,苦荞籽粒的淀粉酶活性提高,使难溶的大分子淀粉降解为小分子水溶性糖类,导致还原糖含量增加。
2.1.6 萌发后矿物质元素含量的变化
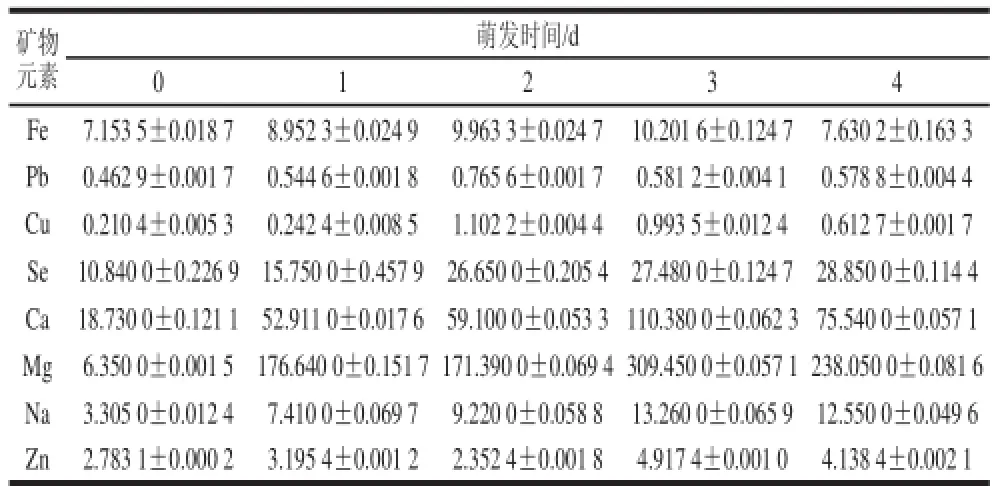
表1 萌发过程中苦荞各矿物元素的含量变化Table 1 Changes in the contents of major mineral elements in tartary buckwheat during germination mg/100 g
由表1可知,随着萌发天数的增加,Mg、Ca的含量波动较大,而Se、Na、Fe、Pb、Zn、Cu这几种矿物元素的含量波动较小。苦荞经萌发处理后其各矿物元素含量均高于未萌发籽粒,其中,Mg是未萌发苦荞籽粒的37.5 倍,其次是Ca、Se、Na,是未萌发苦荞籽粒的5 倍左右,然后是Zn、Pb、Cu是未萌发苦荞籽粒的1~2倍左右,变化不大。
2.1.7 萌发后脂肪酸含量的变化
由表2可知,苦荞脂肪酸的主要成分为棕榈酸、油酸和亚油酸,其中油酸和亚油酸的含量大约占到了相对含量的60%~70%。在苦荞萌发过程中,油酸的增幅最大,亚油酸次之,山嵛酸最小。不同萌发天数时单一脂肪酸的变幅不同。
由图6可知,脂肪酸总量呈现出增大的趋势,不饱和脂肪酸中油酸和亚油酸呈现互为消长的变化。油酸与亚油酸相比,不仅在抗氧化能力方面能力更强外,而且不存在亚油酸、亚麻酸之间的抑制竞争。因此,从不饱和脂肪酸含量和构成上看,萌发后的苦荞具有更好的降低血脂的功效。

表2 萌发过程中苦荞脂肪酸含量变化Table 2 Changes in the content of fatty acids in tartary buckwheat during germination %
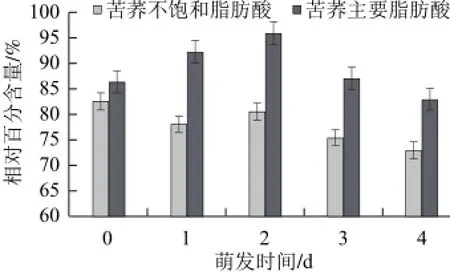
图6 萌发过程中苦荞脂肪酸的含量变化Fig.6 Changes in the contents of unsaturated and major fatty acids in tartary buckwheat during germination
2.1.8 萌发对苦荞氨基酸含量变化及营养价值评价
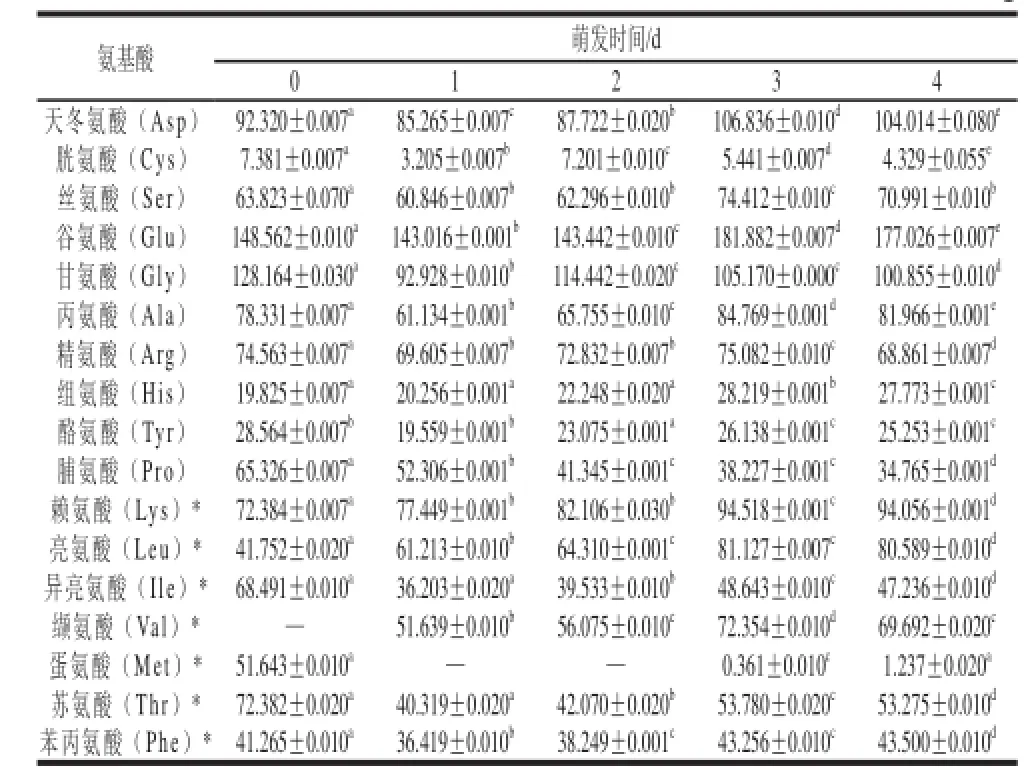
表3 苦荞萌发过程中氨基酸组成分析(x±s,n=3)Table 3 Amino acid profiles of tartary buckwheat during germination (x ±s,n=3) nmol/mg
苦荞籽粒萌发后,由于酶的活性增加,蛋白质等大分子营养物质开始降解,其氨基酸种类虽没有变化,但各种氨基酸含量有较大的变化。苦荞萌发过程中氨基酸变化情况如表3,萌发苦荞中含有17 种氨基酸,其中7 种人体必需氨基酸含量丰富,尤其富含其他谷物中缺乏的赖氨酸(Lys)和豆类中缺乏的蛋氨酸(Met)。不同萌发天数氨基酸含量变化虽存在小幅波动,但随着萌发时间的延长,氨基酸含量变化总体呈现上升趋势,且含量变化均达到显著水平(P<0.05)。在萌发的各个时期,均以谷氨酸(Glu)含量最高,其次为甘氨酸(Gly)、天冬氨酸(Asp),接下来依次为精氨酸(Arg)、丙氨酸(Ala)、丝氨酸(Ser)、亮氨酸(Leu)和缬氨酸(Val),这主要是由于各种酶活力的迅速提高和胚芽乳溶解性的增大。
蛋白质的营养价值主要取决于样品满足人体对氨基酸需要的程度,比较样品中人体必需氨基酸与非必需氨基酸的比值(E/N)、必需氨基酸与总氨基酸的比值(E/T)以及必需氨基酸指数(EAAI),都随着萌发天数的增加呈上升趋势(表4),当萌发至4 d时E/N、E/T和EAAI分别达到最高值52.88%、34.59%和64.1,优于联合国粮食及农业组织/世界卫生组织(FAO/WHO)提出的必需氨基酸模式推荐值(蛋氨酸+胱氨酸和赖氨酸除外),说明萌发后的苦荞营养更高。萌发过程中蛋氨酸+胱氨酸的AAS分最低,说明萌发使苦荞第一限制性氨基酸从赖氨酸转变为蛋氨酸+胱氨酸,EAAI在萌发过程中不断增高,说明萌发苦荞中必需氨基酸的组成因萌发而趋向平衡。
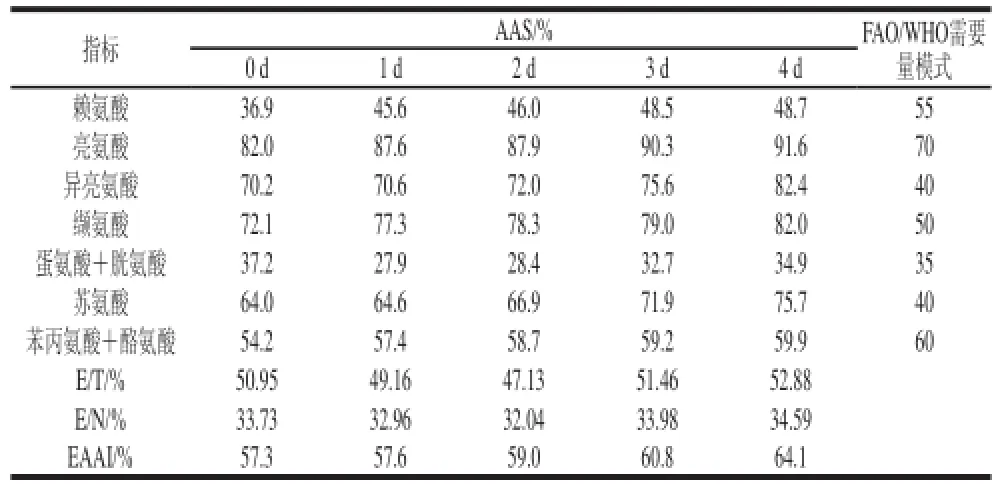
表4 萌发苦荞必需氨基酸组成模式及评分Table 4 Compositional pattern and scoring of essential amino acids in tartary buckwheat during germination
3 结 论
苦荞在萌发过程中吸收水分和氧气,酶类活力增强,生理活动变得活跃,蛋白质、脂肪、多糖等营养物质分解成小肽、氨基酸、脂肪酸和单糖等小分子物质,以满足生长发育和生理需求。萌发苦荞的主要营养物质变化结果表明,苦荞籽粒蛋白质、脂肪和碳水化合物都显著降低,而还原糖、维生素(VB1、VB2)矿物质元素、部分非必需氨基酸和全部必需氨基酸都呈现上升趋势,特别是赖氨酸含量高达77.449~94.518 nmol/mg干质量,远高于小麦和其他禾谷类作物,因此苦荞可以作为人类摄取赖氨酸的重要来源。具体变化结果如下:1)苦荞经过萌发处理后,被测定的17 种氨基酸含量有明显增加,人体必需的7 种氨基酸组成合理、配比适宜,优于FAO/WHO提出的必需氨基酸标准模式谱。因此,苦荞籽粒经萌发后其口感得到改善,可以作为一种口感鲜美的天然食品资源。2)苦荞经过萌发处理后,不饱和脂肪酸含量增加,其主要成分亚油酸是人体必需脂肪酸,具有软化血管,稳定血压,降低血清胆固醇和提高高密度脂蛋白含量的功能。3)苦荞经过萌发处理后,α-、β-淀粉酶活性随着萌发天数的增加而增加,同时,内含的还原糖、水溶性维生素也都随着萌发天数的增加而增加。4)苦荞籽粒经萌发处理后,各矿物元素均高于未萌发籽粒中的含量,其中,Mg和Ca含量较高,且变化幅度较大。
总之,通过本实验对苦荞萌发后各营养成分含量变化的测定,可以看出,苦荞经萌发处理后,其总体营养价值得以提高,口味也得以改善,受到人们的普遍欢迎,有助于萌动食品的开发利用。
[1] 郎桂常. 苦荞麦的营养价值及其开发应用[J]. 中国粮油学报, 1996, 11(3): 9-14.
[2] 韩梅. 苦荞麦的氨基酸含量与营养评价[J]. 天然产物研究与开发, 2000, 12(1): 39-41.
[3] 郑艺梅, 何瑞国, 郑琳, 等. 糙米发芽过程中营养成分及植酸含量变化的研究[J]. 中国粮油学报, 2006, 21(5): 1-4.
[4] IKEDA K, ARIOKA K, FUJII S, et al. Effect on buckwheat protein quality of seed germination and changes in trypsin inhibitor content [J]. Cereal Chemistry, 1984, 61: 236-238.
[5] MOSTAFA M, RAHMA E H. Chemical and nutritional changes in soybean during germination[J]. Food Chemistry, 1987, 23: 257-275.
[6] 陈志刚, 顾振新, 王玉萍, 等. 不同粳稻品种的糙米发芽力及其发芽糙米中主要物质含量比较[J]. 中国粮油学报, 2004, 19(5): 1-3.
[7] 张钟, 王丽, 田奎. 发芽对黑糯玉米淀粉特性的影响[J]. 中国粮油学报, 2006, 21(5): 54-57.
[8] DAUSSANT J, MIYATAS M, ITSUI T, et al. Enzymic mechanism of starch breakdown in germinating rice seeds[J]. Plant Physiology, 1983, 71(1): 88-95.
[9] TAKAHIRO N, SHIGENOBU T, CHIE M E, et al. The physicochemical properties of partially digested starch from sprouted wheat grain[J]. Carbohydrate Polymers, 2004, 56: 271-277.
[10] BRADFORD M M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein dye binging[J]. Analytical Biochemistry, 1976, 72(1): 248-254.
[11] 赵凯, 许鹏举, 谷广烨. 3,5-二硝基水杨酸比色法测定还原糖含量的研究[J]. 食品科学, 2008, 29(8): 534-536.
[12] 王汉屏, 王立志. 木瓜叶芽营养成分含量分析[J]. 食品工业科技, 2007, 28(8): 269-270.
[13] 岳海凤, 郜庆炉, 薛香. 小麦α-淀粉酶活性测定方法比较[J]. 陕西农业科学, 2008, 54(6): 6-7.
[14] 凌腾芳, 林锦山, 刘辉, 等. 一种微量、快速测定植物种子β-淀粉酶活性的方法[J]. 植物学通报, 2006, 23(3): 281-285.
[15] JONES R L, JACOBSEN J V. Regulation of synthesis and transport of secreted proteins in cereal aleurone[J]. International Review of Cytology, 1991, 126: 40-49.
[16] HARDIE G D. Control of carbohydrase formation by gibberellic acid in barley endosperm[J]. Photochemistry, 1975, 14: 1719-1722.
[17] HARA-NISHIMURA I, NISHIMURA M, DAUSSANT J. Conversion of free beta-amylase to bound beta-amylase on starch granules in the barley endosperm during desiccation phase of seed development[J]. Protoplasma, 1986, 134: 149-153.
Nutrient Changes and Nutritional Evaluation of Tartary Buckwheat during Germination
ZHOU Yi-ming1,2, CUI Lin-lin3, WANG Hong2, LI Bao-guo1, ZHOU Xiao-li2,*, XIE Fan2
(1. Institute of Food Science and Engineering, University of Shanghai for Science and Technology, Shanghai 200093, China; 2. School of Perfume and Aroma Technology, Shanghai Institute of Technology, Shanghai 201418, China; 3. Department of Tourism and Food Science, Shanghai Business School, Shanghai 200235, China)
The contents of proteins, crude fat, fatty acids, amino acids, total sugar, reducing sugar, mineral elements, vitamin B1and vitamin B2, and α and β-amylase activities in tartary buckwheat were determined to reveal the changes in nutrients during germination. The results showed that the contents of proteins, fat and total sugar were signifi cantly decreased. Meanwhile, the contents of soluble low molecular sugar, nitrogen-containing substances and fatty acids were greatly increased. The content of reducing sugar was increased from 1.61% to 10.63%, and an increase in total amino acid content of 3.85% was observed. The contents of seven essential amino acids were markedly increased during the fi rst four days of germination, and the lysine content was increased by approximately 30.5% at the end of germination. Non-essential amino acids also displayed an increasing trend. Compared with ungerminated tartary buckwheat, the contents of vitamin B1and B2did not signifi cantly change. Meanwhile, α and β-amylase activities exhibited an increase. During germination, the values of essential to total amino acid ratio and essential amino acid index (EAAI) were increased, the compositional pattern of EAA measured up better to the FAO/WHO standards, and the fi rst limiting amino acid was changed from lysine to methionine and cysteine. In conclusion, the nutritional value is improved in tartary buckwheat during germination.
tartary buckwheat; germination; nutritional components; nutritional value assessment
TS231
A
1002-6630(2014)13-0208-05
10.7506/spkx1002-6630-201413040
2014-04-11
国家自然科学基金面上项目(31371761);“十二五”农村领域国家科技计划课题(2012BAD34B08-11);国家现代农业(燕麦荞麦)产业技术体系建设专项(CARS-08-D-2-5)
周一鸣(1981—),男,讲师,博士研究生,研究方向为食品冷冻冷藏及干燥技术。E-mail:zhouymsit@163.com
*通信作者:周小理(1957—),女,教授,学士,研究方向为食品新资源深度开发与利用。E-mail:zhouxlsit@163.com
