黄瓜过氧化物酶的分离纯化及酶学性质
2014-01-18胡瑞斌王红杨唐云明
胡瑞斌,李 星,王红杨,王 洁,唐云明*
(西南大学生命科学学院,重庆市甘薯工程研究中心,三峡库区生态环境教育部重点实验室,重庆 400715)
黄瓜过氧化物酶的分离纯化及酶学性质
胡瑞斌,李 星,王红杨,王 洁,唐云明*
(西南大学生命科学学院,重庆市甘薯工程研究中心,三峡库区生态环境教育部重点实验室,重庆 400715)
新鲜的黄瓜经匀浆、抽提、硫酸铵沉淀、CM-Sepharose 离子交换层析、Superdex-200凝胶过滤层析后获得电泳纯的过氧化物酶。该酶的比活力、回收率及纯化倍数分别为64 177.67 U/mg、9.58%、61.93。该酶的分子质量为41.15 kD,亚基分子质量为40.21 kD。该酶的最适温度和最适pH值分别为60 ℃和 6。在25~45 ℃及pH 5~9的范围内非常的稳定。在测定条件下测得该酶的Km值为53.79 mmol/L。硫氰化钾对该酶活力基本无影响。尿素、K+、Mn2+、Ca2+、Mg2+、B a2+、Cu2+对该酶都具有激活作用且浓度至50 mmol/L时酶活力分别被激活至112%、127%、113%、128%、139%、199%、348%,然而十二烷基磺酸钠、抗坏血酸和草酸对该酶有强烈的抑制作用;Zn2+、甲醇、乙醇和异丙醇对该酶有一定的抑制作用。
黄瓜;过氧化物酶;分离纯化;性质
过氧化物酶(peroxidase,POD)是由单一肽链与卟啉构成的血红素蛋白。它作为一种氧化还原酶,普遍存在于各种动植物和微生物体内。它具有参与植物体内活性氧的代谢[1]、吲哚-3-乙酸(indole-3-acetic acid,IAA)的降解[2]、木质素的合成[3]等生理功能。该酶在食品检测[4]、酶联免疫分析[5]、生物传感器[6]、污水处理[7]、医疗[8]等方面具有十分重要的应用价值。当今,该酶的商品化生产主要来源于辣根,但其价格比较昂贵。黄瓜作为一种四季常见的绿色蔬菜,分布于我国各地,具有美容、药用等价值并含有多种营养成分[9],且黄瓜廉价易得,黄瓜POD的酶学性质稳定、酶活力较高,目前未见黄瓜POD的分离纯化的研究报道。本实验以黄瓜为材料对POD进行分离纯化并对其酶学性质进行研究,有利于黄瓜的综合开发利用,为该酶提供了新的获取途径,为该酶为进一步研究和应用于工业、环境及生物医学 领域提供参考数据。
1 材料与方法
1.1 材料
以新鲜的黄瓜为提取过氧化物酶的材料,购买于重庆市北碚区菜市场。
1.2 试剂
CM-Sepharose、Superdex-200、凝胶过滤层析分子质量标准品、蛋白质电泳标准品 美国GE Healthcare 公司;考马斯亮兰G-250、R-250 美国Bio-Rad公司;三羟甲基氨基甲烷 香港Farco公司;蓝色葡聚糖-2000、丙烯酰胺、甲叉-双丙烯酰胺 瑞士Fluka公司;其余试剂均为国产分析纯。
1.3 仪器与设备
AKTA prime plus蛋白质纯化系统 美国GE公司;Mill-Q plus超纯水仪 美国Millipore公司;精密电子天平 瑞士Mettler-Toledo公司;UV-2550型分光光度计、蛋白核酸定量仪 日本岛津公司;冷冻干燥仪 德国Uni Equip公司;垂直板电泳槽和电泳仪 美国Bio-Rad公司;GL-21M高速冷冻离心机 湖南湘仪公司;PHS-32W微电路pH计 上海理达仪器厂。
1.4 方法
1.4.1 粗酶液的提取
取新鲜黄瓜,用双蒸水冲洗干净、擦干,去籽去瓤,称重剪碎,然后按1∶1(m/V)比例加入4℃预冷的0.05 mol/L磷酸盐提取缓冲液(pH7.2)匀浆(12 000 r/min、15 s,间隔5 s)后,放置于4 ℃冰箱静置抽提2 h,4 ℃、12 000 r/min离心60 min,取上清液即得粗酶液。
1.4.2 硫酸铵分级沉淀
向粗酶液中加入硫酸铵粉末至35%饱和度,4 ℃盐析2 h,12 000 r/min离心35 min收集上清液;加入硫酸铵粉末至70%饱和度,4 ℃盐析2 h,6 000 r/min离心35 min,收集沉淀;加入0.05 mol/L磷酸盐提取缓冲液(pH7.2)至沉淀完全溶解,4 ℃用0.05 mol/L pH 7.2磷酸盐提取缓冲液透析过夜即得初酶液。
1.4.3 CM-Sepharose层析
经0.05 mol/L pH 5的醋酸缓冲液平衡层析柱后,初酶液加样10 mL,用0~1 mol/L的NaCl溶液(含0.05 mol/L pH 5的醋酸缓冲液)进行线性梯度洗脱,流速0.5 mL/min,每管收集5 mL;测定各管过氧化物酶液的活性和蛋白含量,收集活性较高的酶液透析并冷冻干燥后保存备用。
1.4.4 Superdex-200层析
经0.05 mol/L pH 5醋酸缓冲液平衡层析柱后,取上述冷冻干燥好的POD用0.05 mol/L pH 5醋酸缓冲液溶解,每次加样4 mL,用0.05 mol/L pH 5醋酸缓冲液洗脱,流速0.3 mL/min,每管收集3 mL;测定各管过氧化氢酶液的活性和蛋白含量;收集活性较高的酶液,4 ℃双蒸水透析脱盐冷冻干燥后,获得过氧化物酶纯品,-20 ℃冰箱保存备用。
1.4.5 POD活力的测定
按陈贻竹等[10]的方法测定。测定体系为3 mL,含2.775 mL 0.05 mol/L磷酸盐缓冲液(pH 7.0)、100 μL 4%愈创木酚和100 μL 1% H2O2。加入25 μL酶液后迅速混匀,在25 ℃立即记录2 min内OD470nm的变化。酶活力单位(U)定义为:在该反应条件下,每分钟发生光密度OD470nm改变0.01所需的酶量为1 个酶活力单位。
1.4.6 POD纯度鉴定与分子质量测定[11]
十二烷基硫酸钠-聚丙烯酰胺凝胶电泳(sodium dodecyl sulfate polyacrylamide gel electrophoresis,SDSPAGE)对该酶进行纯度鉴定,分离胶为12%,加样量10μL。经SDS-PAGE电泳和Superdex-200凝胶过滤层析分别测定该酶亚基分子质量与全酶分子质量。
Superdex-200凝胶过滤层析柱用醋酸缓冲液平衡过夜,流速0.3mL/min,将蓝色葡聚糖2000上平衡好的该层析柱,其柱床体积为Vt,测定其外水体积(Vo);将已知分子质量的标准蛋白质上样该层析柱,测定每一标准蛋白质的洗脱体积(Ve)并计算各自的有效分配系数(Kav),其计算公式为:Kav=(Ve-Vo)/(Vt-Vo),然后以标准蛋白质的分子质量的对数lgMr作为X轴,Kav为Y轴,制作蛋白质的标准曲线。按以上方法将黄瓜过氧化物酶上该层析柱并测出其Ve和Kav,根据Kav值查标准曲线并计算即得该酶的分子质量。
1.4.7 蛋白质浓度的测定
分别用紫外分光光度法与考马斯亮蓝染料法(Bradford法)[12]测定该酶的蛋白质浓度。
1.4.8 黄瓜POD的酶学性质测定
1.4.8.1 最适温度和温度稳定性
分别测定在不同温度(25~85 ℃,间隔为5 ℃)下该酶的活力(最适温度条件下,测得的酶活力为100%)和分别将酶液放置在不同温度(25~80 ℃,间隔为10 ℃)下,每隔60 min测定酶活力(25 ℃条件下原酶液的酶活力为100%且测定3 次取平均值),计算该酶的相对酶活力,研究其最适温度和温度稳定性。
1.4.8.2 最适pH值和pH值稳定性
分别测定不同pH值(2.2~10.0)下该酶活力以确定最适pH值;分别将酶液放置在不同的pH值(3~9,间隔为1)的缓冲液中,25 ℃每隔60 min测定该酶的活力,二者均以最适条件下测得的酶活力为100%且测定3 次取平均值,计算该酶相对酶活力,研究其最适pH值和pH值稳定性。
1.4.8.3 米氏常数(Km)的测定
配制不同浓度的过氧化氢(10~50 mmol/L,间隔为10 mmol/L)作为底物,在测定条件下计算POD的酶活力且测定3次取平均值,用双倒数作图法(Lineweaver-Burk法)[13]计算该酶的Km值。
1.4.8.4 部分有机溶剂对POD活性的影响
分别把甲醇、乙醇、异丙醇与醋酸缓冲液(0.05mol/L、pH 5)和酶液混匀(终体积分数分别为10%、20%、30%、40%、50%),4 ℃放置30 min,然后在测定条件下确定该酶的活力(以不加有机溶剂时酶活力为100%且测定3 次取平均值)。
1.4.8.5 部分化合物对POD活性的影响
分别将尿素、草酸、硫氰化钾(potassium thiocyanate,KSCN)、十二烷基磺酸钠(sodium dodecyl sulfonate,SDS)、抗坏血酸(ascorbic acid,ASA),配成100 mmol/L母液,以一定比例与醋酸缓冲液(0.05 mol/L、pH 5)和酶液混合,使得终浓度分别达到10、20、30、40、50 mmol/L,4 ℃放置30 min,然后在测定条件下确定该酶的活力(以不加化合物时酶活力为100%且测定3 次取平均值)。
1.4.8.6 部分金属离子对POD活性的影响
分别将部分金属离子配成100 mmol/L母液,以一定比例与醋酸缓冲液(0.05 mol/L、pH 5)和酶液混合使得终浓度分别达到10、20、30、40、50 mmol/L,4 ℃放置30 min,然后在测定酶活力(以不加金属离子时的酶活力为100%且测定3 次取平均值)。
2 结果与分析
2.1 黄瓜POD的分离纯化
黄瓜POD的粗酶液经CM-Sepharose层析,如图1所示,有3 个酶活力峰,推测该酶可能有3 个同工酶,收集其酶活力最高的第2个峰中的各管,经Superdex-200层析,如图2所示,酶活力集中在29~33管,第31管活性最高,收集活性较高的各管,透析冷冻干燥获得该酶纯品,经SDS-PAGE电泳,为单一条带(图3),说明该酶达到了电泳纯。该酶的整体分离纯化结果如表1所示,可知黄瓜POD纯品的纯化倍数为61.93,回收率为9.58%,酶比活力达到64177.67U/mg。

图1 黄瓜POD的CM-Sepharose离子交换层析Fig.1 CM-Sepharose ion-exchange chromatography of POD from cucumber

图2 黄瓜POD的Superdex-200凝胶过滤层析Fig.2 Superdex-200 gel filtration chromatography of POD from cucumber
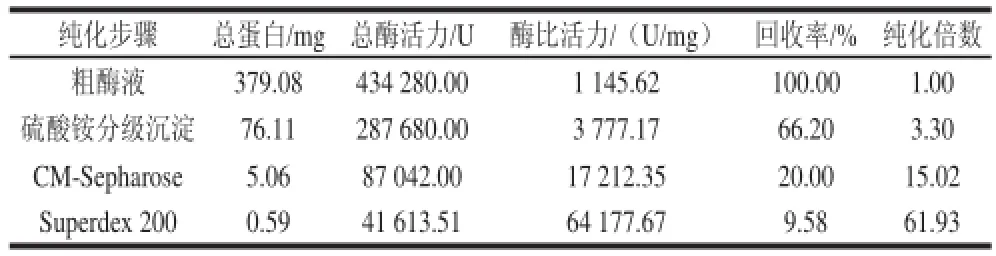
表1 黄瓜POD的分离纯化Table1 Isolation and purification of POD from cucumber
2.2 过氧化物酶的分子质量
经SDS-PAGE后获得电泳纯的单一条带,测得酶的亚基分子质量为40.21 kD(图3),经Superdex-200凝胶过滤层析测得该酶的分子质量为41.15 kD,由此可以判定黄瓜过氧化物酶由单一亚基构成。
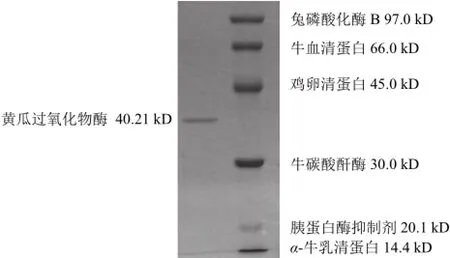
图3 黄瓜POD的SDS-PAGE电泳图谱Fig.3 SDS-PAGE of purified POD from cucumber
2.3 POD的部分酶学性质
2.3.1 最适温度和温度稳定性
在不同温度下测定黄瓜过氧化物酶的活力如图4所示,黄瓜过氧化物酶的最适温度为60 ℃,该酶作用温度范围较广,在30~70 ℃范围内该酶的活力均保持在80%以上,超出此温度,酶活力迅速下降,85 ℃时该酶活力仍保留39%。从图5可知,该酶在25~45 ℃范围内的热稳定性较好,保温5 h酶活力均保持在90%以上;55 ℃保温5 h后酶活力保留49%;高于65 ℃时,酶活力下降迅速;75 ℃保温1 h后,酶蛋白基本完全失活。
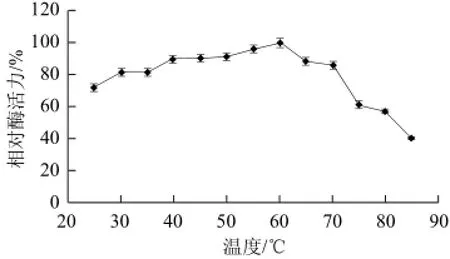
图4 温度对黄瓜POD活力的影响Fig.4 Effect of temperatures on the activity of POD from cucumber
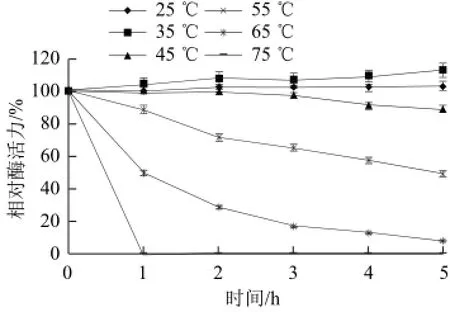
图5 黄瓜POD的热稳定性Fig.5 Thermal stability of POD from cucumber
2.3.2 最适作用pH值和pH值稳定性
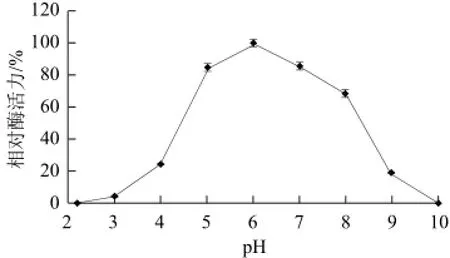
图6 pH值对黄瓜POD活性的影响Fig.6 Effect of pH on the activity of POD from cucumber
如图6所示,黄瓜过氧化物酶的最适pH值为6,在pH 5~7范围内该酶的活力较高,超出此范围酶活力迅速下降。从图7可知,该酶在pH 5~9范围内稳定性较好,保温5 h其活力均保持在85%以上,在pH 3~4保温5 h酶活力还保持在38%以上,表明该酶可以耐受较大范围的pH值,有利于该酶在不同的pH值环境下发挥作用。
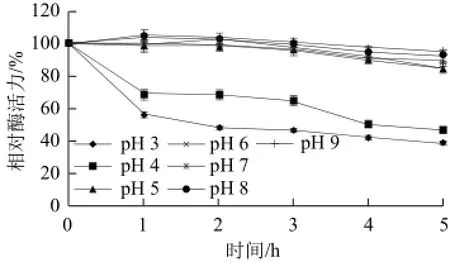
图7 黄瓜POD的pH值稳定性Fig.7 pH stability of POD from cucumber
2.3.3 米氏常数(Km)的测定
Km值的研究结果如图8所示,在测定条件下,以过氧化氢(10~50mmol/L)为底物,测定该酶的活力,采用双倒法作图求得该酶的Km值为53.79mmol/L。

图8 双倒数法测定黄瓜POD的米氏常数Fig.8 Lineweaver-Burk plot for Kmdetermination of POD from cucumber
2.3.4 部分有机溶剂对黄瓜POD活性的影响
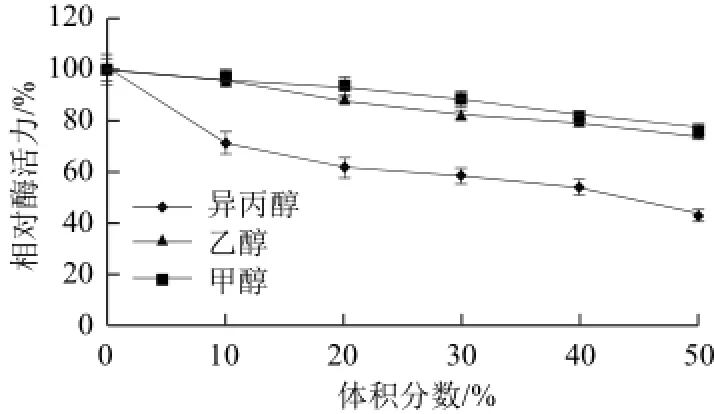
图9 甲醇、乙醇、异丙醇对黄瓜POD活力的影响Fig.9 Effects of methanol, ethanol and isopropyl alcohol on the activity of POD from cucumber
如图9所示,甲醇、乙醇、异丙醇均对该酶有抑制作用,随着三者浓度不断升高,酶活力逐渐受到抑制;异丙醇对该酶抑制作用相对较强,甲醇和乙醇对该酶抑制相对较弱,终浓度达50mmol/L时酶活力均保持在75%以上。
2.3.5 不同化合物对黄瓜POD活性的影响
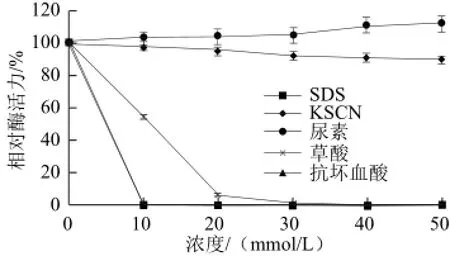
图10 不同化合物对黄瓜POD活力的影响Fig.10 Effects of various compounds on the activity of POD from cucumber
如图10所示,在0~50 mmol/L范围内,随着各自浓度的逐渐增加,KSCN对该酶活力基本无影响,浓度达到50 mmol/L时,酶活力均在90%以上;尿素对该酶有激活作用,且终浓度至50 mmol/L时使酶活力被激活至112%;草酸对该酶抑制作用较强;SDS和抗坏血酸对该酶抑制作用很强,当其浓度达到10 mmol/L时,该酶的活力为0。
2.3.6 不同金属离子对黄瓜POD活性的影响
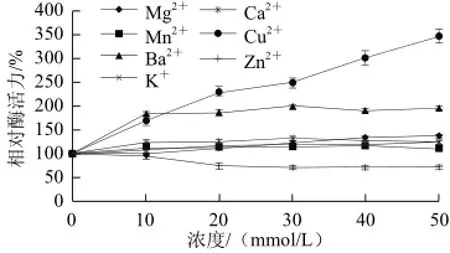
图11 不同金属离子对黄瓜POD活力的影响Fig.11 Effects of metal ions on the activity of POD from cucumber
如图11所示,不同金属离子及同一金属离子不同浓度对该酶活力的影响存在一定差异。Zn2+对该酶有一定抑制作用;K+、Mn2+、Ca2+、Mg2+、Ba2+、Cu2+对该酶有激活作用,且终浓度50mmol/L时该酶的活力分别被激活至127%、113%、128%、139%、199%、348%。
3 讨 论
本实验以黄瓜为材料分离纯化获得电泳纯的过氧化物酶,与其他报道相比较,具有以下特点:一是材料四季常见,成本低廉;二是过氧化物酶含有多种同工酶[14]。粗酶液经CM-Sepharose离子交换层析后检测到3种同工酶,为了获得单一过氧化物酶纯品,选取活力最高的峰作进一步研究,经Superdex-200凝胶过滤层析后仍然得到了较高的回收率(9.58%)、比活力(64177.67U/mg)、纯化倍数(61.93),回收率高于棕榈叶[15](5.8%)、肖黄栌[16](3.3%)和甘薯叶[17](1.59%)来源的POD,比活力和纯化倍数高于来源于椰子[18](12.8U/mg、9.77倍)和辣根[19](26805U/mg、29.3倍)。
本实验所提纯的黄瓜POD的作用温度范围较广,在25~85 ℃均有酶活力且在30~70 ℃时该酶的活力均保持在80%以上,其最适温度为60 ℃,与甘薯叶(60 ℃)[17]来源的相同,高于来源于棕榈叶(55 ℃)[15]、乳(55 ℃)[20]、豆壳(30 ℃)[21]、女贞果实(40 ℃)[22]、莲藕(35 ℃)[23]、香草豆(16 ℃)[24]的POD,高于辣根POD (30℃)和经三聚氰酰氯丝固定化的辣根的POD(40 ℃)[25],表明该酶的最适温度较高。该酶在25~45 ℃范围内的热稳定性非常好,保温5 h酶活力均保持在90%以上,55 ℃保温5 h后酶活力保留49%且热稳定性高于经三聚氰酰氯丝固定化的辣根POD[25]。该酶的最适pH值为6,和来源于辣根(pH 6)[25]和豆壳(pH 6)[21]的POD相同,低于女贞果实POD(pH 6.5)[22]、高于莲藕POD(pH 5)[23]、乳POD(pH5~5.5)[20],远高于香草豆POD(pH 3.8)[24]。pH值耐受范围较广,该酶在pH 5~9范围内稳定性较好,保温5 h其活力均保持在85%以上,在pH 3和pH 4保温1 h酶活力还保留50%以上,其稳定性高于棕榈叶[15]和甘薯叶[17]来源的POD。在水果和蔬菜中POD的分子质量一般介于30~54 kD[26]。该酶的亚基分子质量为40.21 kD,与莲藕(41.5 kD)[23]、红薯(42.03 kD)[27]、苦瓜(43 kD)[28]来源的POD相近,低于椰子来源的POD同工酶(~47、9 kD)[18]、木瓜POD(69.4 kD)[29],高于枇杷果肉POD(22.6 kD)[30],可知该酶的分子结构具有一定的物种差异性。该酶的Km值为53.79 mmol/L,高于豆壳POD(0.41 mmol/L)[21],远低于红甜菜POD(98.61 mmol/L)[31]和甘薯叶POD(291 mmol/L)[17],可知该酶的亲和力相对较高。
甲醇、乙醇、异丙醇对该酶均有抑制作用,与甘薯叶POD[17]所报道的结果一致。KSCN对该酶活力基本无影响,而对甘薯叶POD[17]有非常强烈的抑制作用。SDS、草酸、抗坏血酸对该酶都有显著的抑制作用。SDS作为一种变性剂能破坏蛋白酶分子中的氢键和疏水作用,从而使该酶活力下降为零。草酸对该酶的抑制作用可能是由于维持该酶活性中心的构象进而使该酶的活力不断下降直至失活。抗坏血酸作为一种自由基清除剂能够抑制氢过氧化物进而抑制该酶的活性[32]。尿素对该酶有一定的激活作用。不同的金属离子对POD酶活力的影响也不尽相同,随着金属离子浓度的增大,K+、Mn2+、Ca2+、Mg2+、Ba2+、Cu2+对该酶表现出激活的作用,浓度至50mmol/L时使该酶的活力分别激活至127%、113%、128%、139%、199%、348%,然而其中的Ca2+、Cu2+对辣根来源的POD[19]却有一定的抑制作用。Zn2+对该酶有一定抑制作用,而对莲藕POD[23]具有激活作用,说明不同物种来源的POD在性质上存在一定差异,可能是与所处环境相适应的结果。
[1] BOLWELL G P, WOJTASZEK D. Mechanisms for the generation of reactive oxygen species in plant defence-a broad perspective[J]. Physiological and Molecular Plant Pathology, 1997, 51: 347-366.
[2] ZHENG X, van HUYSTEE A B. On tyrosine oxidation by cationic POD[J]. Plant Cell Tissue Organ Culture, 1991, 25: 35-44.
[3] CHRISTENSEN J H, BAUW G, WELINDER K G, et al. Purification and characterization of peroxidases correlated with lignification in poplar xylem[J]. Plant Physiology, 1998, 118: 125-135.
[4] 冯东, 李雪梅, 王丙莲, 等. 用辣根过氧化物酶生物传感器测定啤酒中的过氧化氢[J]. 酿酒科技, 2011(12): 37-39
[5] 李宗妍, 曹立民, 林洪, 等. 水产品中恩诺沙星残留的一步法酶联免疫检测研究[J]. 食品科学, 2009, 30(10): 231-235.
[6] 吴萍, 蔡称心, 张卉, 等. 辣根过氧化物酶-凹土纳米复合材料及其制备方法以及基于所述复合材料的生物传感器: 中国, G01N27/26; G01N21/35[P]. 2011-09-07.
[7] 郑琦, 李忠铭. 辣根过氧化物酶处理邻苯二酚废水的研究[J]. 化学与生物工程, 2007, 24(7): 65-67.
[8] KIM D S, JEEON S E, PARK K C. Oxidation of indole-3-acetic acid by horseradish peroxidase induces apoptosis in G361 human melanoma cells[J]. Cell Signal, 2004, 16: 81-88.
[9] 乔宏宇, 朱芳, 粟长兰, 等. 黄瓜主要营养品质性状遗传分析[J]. 东北农业大学学报, 2006, 36(3): 290-293.
[10] 陈贻竹, 王以柔. 荔枝果实过氧化物酶(POD)的研究[G]//中国科学院华南植物研究所集刊. 北京: 科学出版社, 1989.
[11] 李建武, 余瑞元. 生物化学实验原理和方法[M]. 北京: 北京大学出版社, 1994: 207-223.
[12] 李建武, 余瑞元. 生物化学实验原理和方法[M]. 北京: 北京大学出版社, 1994: 171-176.
[13] 王镜岩, 朱圣庚, 徐长法. 生物化学[M]. 北京: 高等教育出版社, 2002: 378-380.
[14] 田国忠, 李怀方, 裘维蕃, 等. 植物过氧化物酶研究进展[J]. 武汉植物学研究, 2001, 19(4): 332-344.
[15] AL-SENAIDY A M, ISMAEL M A. Purification and characterization of membrane-bound peroxidase from date palm leaves (Phoenix dactylifera L.)[J]. Saudi Journal of Biological Sciences, 2011, 18: 293-298.
[16] KUMARA R, SINGH K A, SINGH V K, et al. Biochemical characterization of a peroxidase isolated from Caribbean plant: Euphorbia cotinifolia[J]. Process Biochemistry, 2011, 46: 1350-1357.
[17] 付伟丽, 唐靓婷, 王松, 等. 甘薯叶过氧化物酶的分离纯化及其部分性质研究[J]. 食品科学, 2010, 31(7): 223-227.
[18] BALASUBRAMANIAN M, BOOPATHY R. Purification and characterization of peroxidases from liquidendosperm of Cocos nucifera (L.): biotransformation[J]. Journal of Molecular Catalysis B: Enzymatic, 2013, 90: 33-42.
[19] MOHAMED S A, ABULNAJA K O, ADS A S, et al. Characterisation of an anionic peroxidase from horseradish cv. Balady[J]. Food Chemistry, 2011, 128: 725-730.
[20] 卢蓉蓉, 许时婴, 王璋. 乳过氧化物酶的分离纯化和酶学性质研究[J].食品科学, 2006, 27(2): 100-104.
[21] 赵杨. 豆壳过氧化物酶的分离纯化、酶学性质及固定化研究[D]. 长春: 长春工业大学, 2011: 34-36.
[22] 王瑧, 廖祥儒, 张建国, 等. 女贞果实过氧化物酶的纯化及热稳定性研究[J]. 河北农业大学学报, 2007, 30(5): 23-27.
[23] 阙瑞琦, 张丽丽, 郭小路, 等. 莲藕过氧化物酶的分离纯化及性质研究[J]. 西南大学学报: 自然科学版, 2007, 29(12): 63-67.
[24] OFELIA M N, WALISZEWSKIA K N, OLIARTA R M, et al. Purification and characterization of cell wall-bound peroxidase from vanilla bean[J]. LWT-Food Science and Technology, 2008, 41(8): 1372-1379.
[25] MOHAMED S A, DARWISH A A, El-SHISHTAWY R M, et al. Immobilization of horseradish peroxidase on activated wool[J]. Process Biochemistry, 2013, 48: 649-655.
[26] VIGYAZOA L V, HAARD N F. Polyphenol oxidase and peroxidase in fruits and vegetables[J]. Critical Reviews in Food Science and Nutrition, 1981, 15: 49-127.
[27] ROMPEL A, ALBERS M, NASERI J I, et al. Purification, cloning and characterization of a novel peroxidase isozyme from sweetpotatoes (Ipomoea batatas)[J]. Biochimica et Biophysica Acta-Proteins and Proteomics, 2007, 1774: 1422-1430.
[28] FATIMA A, HUSAIN Q. A role of glycosyl moieties in the stabilization of bitter gourd (Momordica charantia) peroxidase[J]. International Journal of Biological Macromolecules, 2007, 41: 56-63.
[29] CHEN L C, CHUNG Y C, CHANG C T, et al. Characterisation of an acidic peroxidase from papaya (Carica papaya L. cv Tainung No. 2) latex and its application in the determination of micromolar hydrogen peroxide in milk[J]. Food Chemistry, 2012, 35(4): 2529-2535.
[30] 林建城, 吴智雄, 彭在勤. 枇杷果肉过氧化物酶的分离纯化及其性质研究[J]. 四川农业大学学报, 2007, 25(4): 419-424.
[31] RUDRAPPA T, LAKSHMANAN V, KAUNAIN R, et al. Purification and characterization of an intracellular peroxidase from genetically transformed roots of red beet (Beta vulgaris L.)[J]. Food Chemistry, 2007, 105: 1312-1320.
[32] POPP J L, KALYANARAMAN B, KIRK T K. Lignin peroxidase oxidation of Mn2+in the presence of veratryl alcohol, malonic or oxalic-acid, and oxygen[J]. Biochemistry, 1990, 29 (46): 10475-10480.
Isolation, Purification and Characterization of Peroxidase from Cucumber
HU Rui-bin, LI Xing, WANG Hong-yang, WANG Jie, TANG Yun-ming*
(Key Laboratory of Eco-environments in Three Gorges Reservoir Region, Ministry of Education, Chongqing Sweet-Potato Engineering Research Center, School of Life Sciences, Southwest University, Chongqing 400715, China)
Electrophoresis-purity peroxidase was extracted from fresh cucumber and purified through homogenization, extraction, ammonium sulfate precipitation, CM-Sepharose and Superdex-200 chromatography. The specific activity, recovery and purification fold of the peroxidase were 64 177.67 U/mg, 9.58% and 61.93, respectively. Molecular mass of this purified enzyme was 40.21 kD as determined by sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE), while native gel filtration confirmed a monomer form of 41.15 kD. Besides, the peroxidase, whose optimum temperature and pH were 60 ℃ and 6, respectively, was comparatively stable in the temperature range of 25-45 ℃ and pH range of 5-9. The purified peroxidase showed Kmvalue of 53.79 mmol/L under definite conditions. In addition, its activity was scarcely affected by potassium thiocyanate (KSCN). The peroxidase was found to be activated by urea, K+, Mn2+, Ca2+, Mg2+, Ba2+and Cu2+by 12%, 27%, 13%, 28%, 39%, 99% and 248% at 50 mmol/L, respectively. However, the peroxidase activity was significantly inhibited by SDS, ascorbic acid and oxalic acid. Moreover, Zn2+, methanol, ethanol a nd isopropanol caused partial inhibitory effects on the peroxidase.
cucumber; peroxidase; isolati on and purification; characterization
Q946.5
A
1002-6630(2014)11-0168-06
10.7506/spkx1002-6630-201411034
2013-07-09
重庆市科委重点攻关项目(CSTC2011AB1027)
胡瑞斌(1985 —),男,硕士研究生,主要从事蛋白质与酶工程研究。E-mail:huruibin55@163.com
*通信作者:唐云明(1960 —),男,教授,博士,主要从事蛋白质与酶工程研究。E-mail:tbright@swu.edu.cn
