黑龙江省水稻品种抗病基因同源序列多态性分析
2014-01-14邹德堂任月坤王敬国刘化龙
邹德堂,任月坤,王敬国,刘化龙
(东北农业大学农学院水稻研究所,哈尔滨 150030)
黑龙江省水稻品种抗病基因同源序列多态性分析
邹德堂,任月坤,王敬国,刘化龙
(东北农业大学农学院水稻研究所,哈尔滨 150030)
根据60份水稻品种对45株稻瘟病菌株的抗性表现,结合RGA-PCR法对黑龙江省60份水稻种质资源进行抗病性分析。结果表明,不同品种间抗病频率变化较大,变幅为0~93.3%。以抗病基因同源序列多态性聚类,取遗传相似系数0.680时,可将供试品种划分为三个类群。取遗传相似系数0.820时。类群Ⅰ可划分3个亚类,类群Ⅱ可划分为7个亚类,类群Ⅲ可划分为2个亚类。以单孢菌株接种的抗性表型聚类,取相似系数0.440时,可将60个品种划分为两个类群。综合利用抗病表型结合RGA分析可以更加全面地反映品种间的抗性遗传背景,为抗病亲本选育和水稻混合间栽控制稻瘟病研究提供理论依据。
稻瘟病;抗病基因;同源序列
稻瘟病是影响水稻生产最重要的流行性病害之一,发病年份一般减产10%~20%,严重时高达40%~50%,部分田地甚致绝产[1]。自20世纪60年代以来,因氮肥施用量增加和品种抗病性减弱,黑龙江省稻瘟病每年都有不同程度发生,种植品种单一化,寄主与病原菌相互作用形成定向选择,加之稻瘟病菌生理小种变异快,易导致品种的抗病性减弱甚至丧失[2]。实践证明,探明品种间抗性来源及差异是控制稻瘟病最经济有效的策略。迄今,在已克隆的14个稻瘟病抗性基因中,按基因的保守结构域进行分类,发现有12个稻瘟病抗性基因属于NBS-LRR类,这类基因编码的产物在氨基端都含有核苷酸结合位点(NBS),而在羧基端则都具有富亮氨酸重复序列(LRR)[3]。根据抗病基因保守结构域设计特异引物或简并引物,通过PCR扩增同源序列DNA片段,即RGA-PCR法,为筛选抗病基因或克隆相关抗病基因提供有效手段,同时也是快速评价种质资源遗传多样性的有效方法[4]。
黑龙江省是中国水稻种植的北限,是寒地水稻的主要种植区域,针对水稻全省划分为4个积温带,种植品种的积温、生育期和主茎叶片数分别大致变化在2 300~2 800℃、130~150 d和10~14片叶之间。黑龙江省拥有较丰富的寒地水稻品种资源,但品种存在亲缘关系近、遗传基础狭窄的致命缺点[5]。因此,明确品种抗性基因组成对于保证黑龙江省水稻安全生产、提高品种稻瘟病抗性均具有重要意义。本研究利用抗病基因同源序列法对黑龙江省部分骨干亲本和主要育种单位的部分育成品种进行抗病遗传基础分析和稻瘟病人工接种,旨在揭示品种间抗性遗传背景差异,为黑龙江省抗病品种合理利用和抗病育种提供理论依据。
1 材料与方法
1.1 材料与菌株
试验材料共60份,包括骨干亲本4份(石狩白毛、上育397、五优稻一号、富士光)[6],各时期主栽品种49份,以及中国的7个稻瘟病鉴别寄主。其中除特特普、珍龙13、四丰43为籼稻,其余57个品种均为粳稻。所有水稻材料由东北农业大学水稻育种实验室提供。供试菌株于2011年秋季采集于黑龙江省11个水稻种植区,分离后获得单孢菌株45个,经鉴定分为6群(A、B、C、D、E、F)7个生理小种。
1.2 稻瘟病生理小种和主栽品种抗病鉴定
将中国7个稻瘟病鉴别寄主以及53个主栽水稻品种分成45个小区播种育苗,出苗后每隔3~4 d喷施营养液,确保稻苗茁壮生长。待稻苗长到三叶一心时分别对各小区进行稻瘟病单孢菌株喷雾接种。10 d后按Mackill等[7]评价标准调查各品种发病情况,将0~3级记为抗病(R),4~5级记为感病(S)。并且计算每个品种的抗病频率。抗病频率(%)=表现R反应的菌株数/总菌株数×100%。
1.3 DNA提取与扩增
取三叶一心时期的水稻幼叶,参照Zheng等方法[8]并稍加修改后提取水稻全基因组DNA。所用引物见表1。所有引物由上海生工生物工程有限公司合成。PCR反应总体积为10.0 μL,包括5 μL ddH2O、1.0 μL 10×PCR Buffer、0.75 μL Mg2+(1.0 mmol·L-1)、0.15 μL dNTPs(0.2 mmol·L-1)、0.1 μL Taq酶(5 U·μL-1)、引物1.5 μL、DNA模板1.5 μL。
反应程序:94℃预变性5 min;95℃变性1 min,45℃退火1 min,72℃延伸2 min,35个循环;72℃完全延伸7 min;4℃保存。PCR反应在Eppendorf 5333型PCR仪中进行,扩增产物通过6%聚丙烯酰胺凝胶电泳进行分离,采用Promega公司的DNA银染系统进行显色。2次重复。
1.4 数据处理
根据水稻品种对单孢菌株的抗感病反应,将抗病(R)记为“1”,感病(S)记为“0”。在荧光灯下读取RGA-PCR扩增产物的变性聚丙烯酞胺凝胶电泳图谱带。根据两次扩增谱带的有无记录,有带的记为“1”,无带的记为“0”。将数据制成Microsoft Excel文件,用于聚类分析和扩增谱带的多态性比较。数据处理采用NTSYSpcVersion2.10e(Applied Biostatistics Inc.)USA软件中的FLEXI(Flexiblebeta method)程序进行聚类分析,构建聚类图谱。
2 结果与分析
2.1 水稻品种抗病性反应
根据60份水稻品种对45个稻瘟病单孢菌株的抗感反应,计算每个品种的抗病频率(见表2)。品种间抗病性差异变化较大,抗病频率在0~93.3%。龙粳14和东农415的抗病频率大于90%,农林20和丽江新团黑谷的抗病频率则为0。高抗品种8个,中抗品种33个,中感品种14个,高感品种5个。

表1 引物序列及来源Table 1 Sequence and origin of RGA primer pairs used in this research
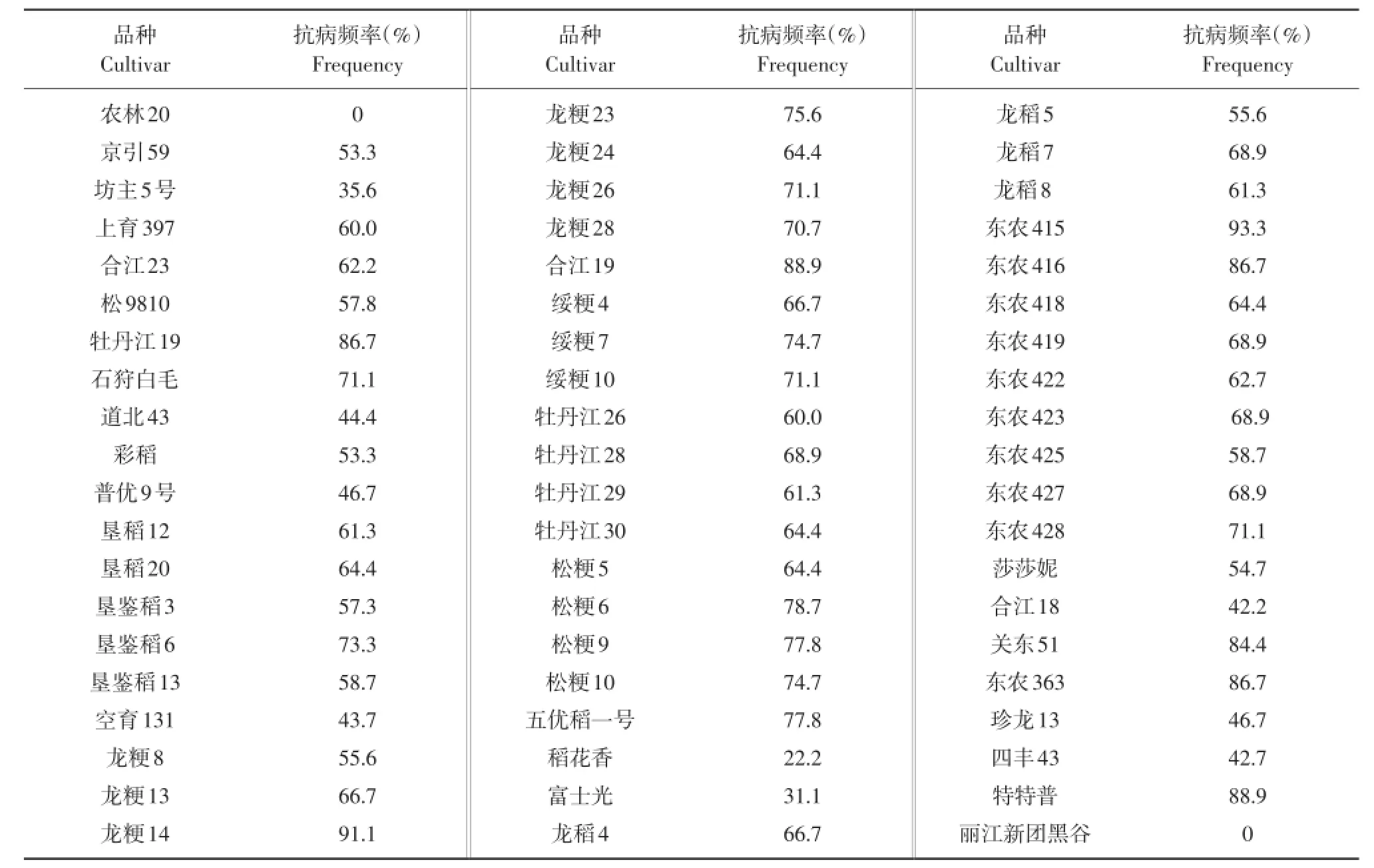
表260 个水稻品种的抗病性Table 2 Resistance to blast of 60 rice varieties
2.2 水稻抗病基因同源序列的多态性
由RGA引物扩增的DNA片段大小在60 bp~ 3 kb,多态性谱带主要集中在500 bp~2 kb。引物多态性信息详见表3。选取6对引物对供试的60个水稻品种共扩增出157条清晰谱带,其中呈多态性带数共58条,占总带数的36.94%。就单个品种而言,不同引物间扩增出的条带数存在差异,引物XLRR扩增出的条带数最多,这可能与XLRR是根据水稻抗病基因保守结构域设计的有关。就引物多态性来讲,其中引物S2/AS1多态性比较丰富,高达82.60%。而引物NLRR多态性则较差,只有8.96%。
2.3 基于RGA相似性对品种进行聚类
大多数RGA与抗病基因紧密连锁,甚至就是抗病基因的一部分。因此,研究RGA在基因组中的分布和演化在一定程度上可以反映R基因的情况。根据6对RGA引物对60个品种扩增的多态性谱带的相似系数,将6对引物单独或组合,利用NTSYS程序进行聚类分析。将所得结果一一与稻瘟病表型聚类相比较,最终选取3对较适合稻瘟病抗性基因同源序列相似性分析的引物(RGA2/RGA5/ RGA6)组合,利用NTSYS程序进行聚类分析,结果见图1。遗传相似系数范围在0.65~1.00,说明供试品种在抗性遗传基础上存在一定差异。当相似系数取0.680时,可将60个品种划分为3个类群。其中Ⅱ为主要类群,包括35个水稻品种。当遗传相似系数取0.820时,类群I又可划分为3个亚类,其中牡丹江19单独为一亚类,这可能与其强抗病性有关。类群II又可划分为5个亚类,各亚类间存在一定的遗传差异。类群Ⅲ又可划分为2个亚类,其中三个籼稻品种珍龙13、四丰43和特特普聚在一个亚类,且与其他粳稻品种遗传距离较大。I、II类群中都有抗病基因同源序列完全相同的品种。如I类群中的上育397和彩稻,Ⅱ类群中的东农423、东农425、东农427、东农428,说明这些供试品种在抗性遗传上几乎无差异。总体来讲,类群I中抗病频率高的品种所占比例较高,石狩白毛、合江19、牡丹江19都是各时期的高抗稻瘟病品种。同一亚类内的品种具有相似的抗病性,如五优稻一号、稻花香、富士光、龙稻4等品种在生产实践中稻瘟病抗性表现均较差。另一方面,抗性基因同源序列多态性分析在一定程度上能够反映品种间的亲缘关系,如类群I中的龙粳14和绥粳7均是合江22的衍生品种。类群II中的牡丹江28、牡丹江29、牡丹江30均为黑龙江省农科院牡丹江分院选育,几乎都含有上育397的血缘。
2.4 基于抗稻瘟病表型聚类分析
根据水稻品种对45个单孢菌株接种抗感病反应转化而来的数据,利用NTSYS程序进行聚类分析,结果见图2。当相似系数取0.440时,可将60个品种划分为A、B两类。A类只包括农林20、道北43、丽江新团黒谷3个品种,丽江新团黒谷和农林20抗病频率最低表现为全感。B类包括57个水稻品种,抗病频率22.2%~93.3%。当相似系数取0.636时,B类又划分为5个亚类。第ⅰ、ⅱ亚类为主要亚类,ⅰ亚类39个品种的平均抗病频率为56.3%。ⅱ亚类15个品种的平均抗病频率为81.0%。两亚类之间差异明显。
2.5 RGA相似性与水稻抗稻瘟病表型聚类比较
植物抗病性不仅决定于植物的基因型还取决于病原菌的基因型以及外部环境条件,因而品种的感病性是相对的。就水稻抗稻瘟病而言,利用同一稻瘟病单孢菌株接种不同的水稻品种明显地表现出不同的抗感反应。从本研究选用的60个水稻品种来看,用抗感单孢稻瘟病菌表型聚类,倾向于抗谱相似的聚为一类;按RGA相似性聚类,倾向于遗传背景相近的聚为一类。RGA聚类中相同类群的品种抗病性也存在差异,如Ⅰ类群中ⅰ亚类的合江19和合江23,二者抗谱相差26.7%。II类群中ⅰ亚类的龙粳8和龙粳23,二者抗谱相差20%。这种遗传背景相似而抗病频率差异大情况的发生,可能是由于广谱抗性基因存在造成的。对于抗病频率低或是抗病频率高的品种两者类与类之间有较好的对应关系,例如II类群中的ⅴ亚类与B类中的ⅰ亚类,抗病频率低的富士光、稻花香对应较好;B类中的ⅱ亚类和Ⅰ类部分对应较好,如抗病频率高的合江19、牡丹江19、龙粳14对应较好。但总体看类与类之间没有明显的对应关系,不过两者结合起来更能准确反映水稻材料的抗病性遗传背景。
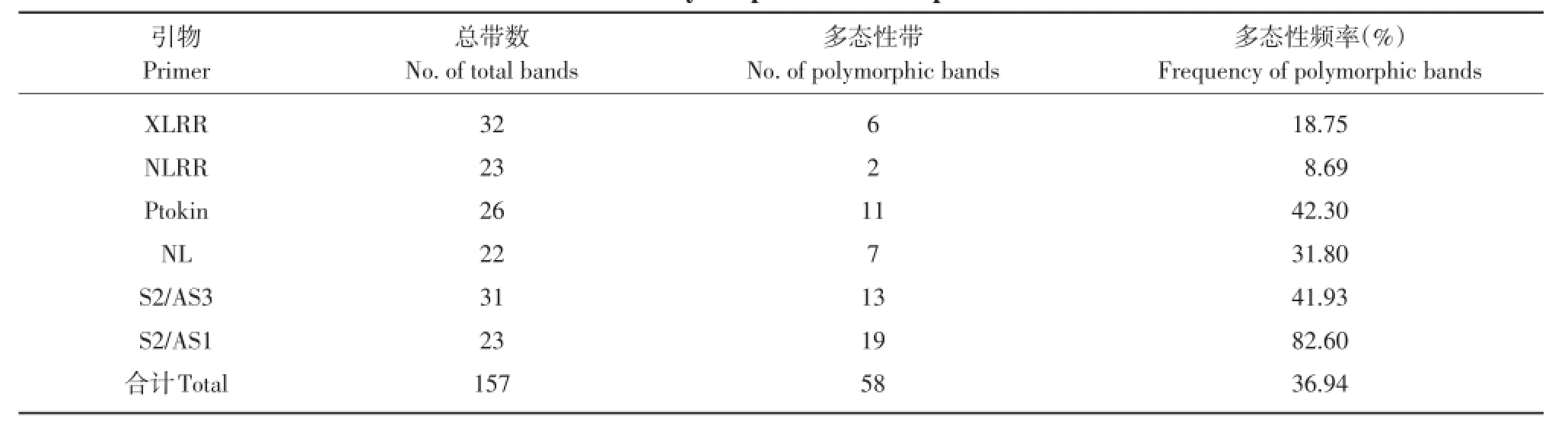
表3 抗病基因同源序列引物多态性Table 3 Polymorphism of RGA primers

图1 供试品种对3对引物的RGA指纹聚类Fig.1 Dendrogram of 60 rice cultivars based on RGA bands generated by three pairs of primers

图2 稻瘟病抗性表型聚类Fig.2 Clustering based on phenotypes of blast resistance
3 讨 论
从水稻抗稻瘟病的表型到抗病基因型,是复杂的生理生化过程,既涉及到稻瘟病菌株与水稻自身的抗性基因的互作,又涉及到稻瘟病菌株与环境之间的互作与选择[10]。这也是同一品种在不同生育期、不同栽培管理措施下、不同生长环境中,抗性会有很大差异的原因。因此抗稻瘟病的表型聚类存在一定误差。由此可见,抗性基因的表达受基因本身、病原菌的无毒基因和环境的影响,会产生R基因与表型不一致现象。
RGA相似性聚类与抗性表型聚类结果相比,对于抗病频率低或抗病频率高的品种两者类与类之间有较好的对应关系,但总体看来类与类之间没有明显的一一对应关系,这与刘二明等研究结果一致[11]。孙雁等研究表明,用RGA多态性聚类得到的类群与其系谱来源基本相符[12]。以往研究表明,黑龙江省稻瘟病菌生理小种组成复杂,分布较广,且年际间优势生理小种变化明显。张国民等分析得出黑龙江省水稻品种遗传变异性较小,推广品种的抗稻瘟病基因型主要集中在少数几个抗病基因上,如Pi-a、Pi-i、Pi-k、Pi-b和Pi-ta等[13],因此有必要对黑龙江省抗稻瘟病种质资源进行进一步发掘、研究与利用,明确品种的抗性基因、优化品种布局。本研究通过RGA-PCR法从基因组水平分析水稻品种间差异,加之与稻瘟病表型结合,能更加准确反映品种间的抗性遗传背景。在进行抗性育种时,若利用已知持久抗瘟性和广谱抗性的品种,如本研究中的东农415、牡丹江19、合江19、龙粳14等品种,选择与之相似系数较小、不在同一类群中、且具有优良农艺性状的品种作杂交父母本,如本研究中的稻花香、五优稻一号等,通过单交、复交、聚合杂交等杂交方式使同一品种具有多个抗病基因组合,育成具有持久抗性的优良品种,拓宽水稻抗病种质的遗传基础。此外,可通过对具有不同抗性遗传背景的水稻品种进行合理布局,减缓水稻与稻瘟病菌之间互作,维持自然界病菌群体相对稳定,达到持续控制稻瘟病目的[14]。
[1]孙国昌,杜新法,陶荣祥,等.水稻稻瘟病防治策略和21世纪研究展望[J].植物病理学报,1998,28(4):289-292.
[2]马成云,申宏波,马淑梅.黑龙江省水稻稻瘟病发生危害情况调查及防治建议[J].植物保护,2006,32(5):95-97.
[3]刘鹏,魏毅东,陈由强,等.水稻稻瘟病抗性基因的归类分析及其功能研究进展[J].分子植物育种,2011,9(2):128-135.
[4]Chen X M,Line R F,Leung H.Genome scanning for resistancegene analogs in rice,barley and wheat by high-resolution electrophoresis[J].Theor Appl Genet,1998,97:345-355.
[5]刘宝海,宋福金,高存启,等.黑龙江大面积推广水稻品种遗传基础研究[J].作物杂志,2004(2):48-52.
[6]刘化龙,陈温福,徐正进,等.黑龙江水稻育种骨干亲本及系谱分析[J].东北农业大学学报,2011,42(4):18-21.
[7]Mackill D J,Bonman J M.Inheritance of blast resistance in nearisogenic lines of rice[J].Phytopathology,1992,82(7):746-749.
[8]Zheng K L,Subudi P K,Domigo J,et al.Rapid DNA isolation for marker-assisted selection in rice breeding[J].Rice Genet Newsl, 1995(12):255-258.
[9]Doyle J J,Doyle J L.Isolation of plant DNA from fresh tissue[J]. Focus,1990,12(1):13-15.
[10]Yoshimura S,Yamanouchi U,Katayose Y,et al.Expression of Xa1, a bacterial blight-resistance gene,is induced by bacterial inoculation[J].Proc Natl Acad Sci USA,1998,95:1663-1668.
[11]刘二明,肖一龙,易有金,等.水稻品种抗瘟性表型与抗病基因同源序列相似性关系[J].中国水稻科学,2005,19(3):209-216.
[12]孙雁,王云月,万瑞亭,等.黑龙江水稻品种抗病基因同源序列聚类分析[J].云南农业大学,2001,16(2):107-110.
[13]张国民,辛爱华,马军韬,等.黑龙江省水稻稻瘟病研究回顾与展望[J].黑龙江省农业科学,2008(6):156-158.
[14]孙雁,王云月,何月秋,等.云南稻种抗病基因同源序列类似性分析[J].中国农业科学,2002,35(5):502-507.
Analysis on resistance gene analog polymorphism of rice varieties in Heilongjiang Province
ZOU Detang,REN Yuekun,WANG Jingguo,LIU Hualong(Rice Research Institute,School of Agriculture,Northeast Agricultural University,Harbin 150030,China)
Comparison of clustering analysis was investigated using spectrum of resistance to blast and polymorphism of resistance gene analog(RGA)in 60 varieties for blast resistance identification.The results showed that the resistance frequency changed greatly in different varieties, which ranged from 0-93.3%.The RGA-PCR clustering analysis showed that 60 cultivars could be divided into three groups with the genetic similarity coefficient of 0.680.GroupⅠcould be divided into three subclasses,groupⅡcould be divided into seven subclasses and GroupⅢcould be divided into two subclasses with 0.820 genetic similarity coefficient.The resistance spectrum clustering analysis showed that 60 varieties could be divided into group A and group B with the genetic similarity coefficient of 0.440.The varieties were genetically highly diversed and completely distinguished from RGA analysis and resistance analysis.Therefore,the RGA technique combining the resistance frequency,reflects the resistance in different varieties more comprehensively and provides an useful and efficient way to improve the efficiency of parent selection in rice breeding and theoretical basis for mixed cultivation of rice varieties to control rice blast.
rice disease blast;resistance gene;homologous sequence
S511
A
1005-9369(2014)03-0001-07
时间2014-3-20 17:47:00 [URL]http://www.cnki.net/kcms/detail/23.1391.S.20140320.1747.004.html
邹德堂,任月坤,王敬国,等.黑龙江省水稻品种抗病基因同源序列多态性分析[J].东北农业大学学报,2014,45(3):1-7.
Zou Detang,Ren Yuekun,Wang Jingguo,et al.Analysis on resistance gene analog polymorphism of rice varieties in Heilongjiang Province[J].Journal of Northeast Agricultural University,2014,45(3):1-7.(in Chinese with English abstract)
2012-07-24
“十二五”农村领域国家科技计划项目(2011BAD35B02-01);国家科技支撑项目(2011BAD16B11)
邹德堂(1965-),男,教授,博士,博士生导师,研究方向为水稻分子遗传育种。E-mail:zoudt@163.com
