水稻直链淀粉含量QTL图谱整合研究
2014-01-14李修平马文东刘方明吴恒梅
李修平,马文东,王 仲,郭 勇,刘方明,吴恒梅
(1.佳木斯大学生命科学学院,黑龙江 佳木斯 154007;2.黑龙江省农业科学院佳木斯水稻研究所,黑龙江 佳木斯 154026)
水稻直链淀粉含量QTL图谱整合研究
李修平1,马文东2,王 仲1,郭 勇1,刘方明1,吴恒梅1
(1.佳木斯大学生命科学学院,黑龙江 佳木斯 154007;2.黑龙江省农业科学院佳木斯水稻研究所,黑龙江 佳木斯 154026)
水稻直链淀粉含量是水稻品质育种重要指标之一。研究收集整理来自34个作图群体共141个与水稻直链淀粉含量相关的QTL信息。利用Biomercator 2.1软件和共有标记映射,将收集到的QTLs整合到参考图谱Cornell 2001上,通过元分析方法获得“真实QTLs”,建立水稻直链淀粉含量QTLs的一致性图谱,在第1、2、3、6、7、8、9和10染色体上共分析得到30个“真实QTLs”及其连锁标记。为水稻直链淀粉含量基因的精细定位、图位克隆及水稻品质的分子辅助育种提供理论基础。
水稻;直链淀粉含量;QTL整合;元分析
直链淀粉含量是影响稻米食味品质的主要因素之一。探明水稻直链淀粉含量遗传规律,对稻米品质改良具有重要意义。随着水稻基因组测序计划的完成,水稻高密度分子遗传连锁图谱和各种近等基因系群体构建,与水稻直链淀粉相关数量性状位点不断被定位。Lanceras等利用组合KD⁃ ML105/CT9993衍生的RIL群体,在第3、4、6和7染色体定位到4个影响AC的QTLs,解释AC变异的80%[1]。王茂青利用日本优质粳稻品种越光和国内著名高产水稻品种桂朝二号构建的重组自交系群体,共检测到3个控制直链淀粉含量的加性QTL,分布于第1、2和6染色体[2]。由于群体数量小和作图密度低等原因,使得已发表的QTLs存在标记区间大、精度低、置信区间大等问题,导致在不同研究中定位的QTL(即使是同一位点)存在位置差异,无法确定“真实的”QTLs,很难找到与直链淀粉含量紧密连锁的分子标记,至今能够在育种中应用的有效标记较少。
图谱整合是弥补单个作图群体因分子标记多态性的局限性而难以构建高密度图谱的有效方法。张闻博等将与抗大豆胞囊线虫相关的151个QTLs整合并利用元分析技术发掘了16个“真实”QTL[3]。刘硕等将已发表与大豆蛋白质相关的QTL进行整合,得到连锁群上QTL的有效遗传位置,并筛选出稳定性标记Satt127[4]。高利芳等对大豆株高QTL进行整合,得到15个株高的“通用”QTL,分别位于大豆6、7、11、13和18号染色体[5]。
本研究对Gramene数据库及发表的与水稻直链淀粉含量相关的QTLs进行整合并利用元分析技术进行“真实”QTL挖掘,以期为水稻直链淀粉含量遗传机制研究及水稻品质分子辅助育种提供理论依据。
1 材料与方法
1.1 水稻直链淀粉含量QTL的收集和整理方法
收集水稻直链淀粉含量相关QTLs定位信息包括QTL名称、所在染色体位置、临近标记、LOD、贡献率、作图群体和作图分析方法等。整理方法为:对于原始图谱和参考图谱共有的标记,若QTL为单标记,则记录其在原始图谱上的坐标;若QTL为双标记,则分别记录双标记在原始图谱上的坐标,即左标记和右标记。位置(最大可能性的位置及其置信区间)和贡献率作为QTL的2个重要参数,若QTL的置信区间未知,则根据Darvasi等的公式计算95%置信区间[6]:

C.I.为QTL的置信区间,N为作图群体的大小,R2为QTL的贡献率。公式(1)适用于回交和F2群体,公式(2)适用于RILs群体。
本研究收集来自不同亲本的34个作图群体,共141个与水稻直链淀粉含量相关的QTLs(见表1)。

表1 已报道的水稻直链淀粉含量相关QTLsTable 1 QTLs reported on amylose content of rice
1.2 水稻直链淀粉含量相关QTLs的映射
QTL映射是利用奇序函数(Homothetic func⁃tion)计算左右标记与QTL的距离,将原始图谱上的QTL按比例投射到参考图谱上。利用软件Bio⁃Mercator 2.1的映射功能将其他作图群体的水稻直链淀粉含量相关QTL映射到参考图谱,构建一张水稻直链淀粉含量相关QTL“一致性图谱”。
由于已报道的QTL来源于不同的作图群体,具有不同研究背景以及分析方法,定位在各自连锁图谱上,且图谱密度较小,因此QTL定位的原始图谱之间共同标记较少。本研究采用Cornell 2001作为参考图谱,该图谱为高密度遗传连锁图谱,具有较高的可信度,与原始图谱间有大量的共同标记,便于图谱及QTL整合。
1.3 水稻直链淀粉含量QTL的元分析
利用BioMercator 2.1软件中的Meta-analysis程序进行水稻直链淀粉含量QTL的元分析。对同一染色体上同一座位或者有重叠座位的QTL计算出一个“一致性”QTL,该QTL会给出5个计算模型,其中AIC(Akaiketype criteria values)值最小的模型为最优模型,即最优“一致性”QTL,该模型也是比较接近“真实”QTL的模型。每个模型都是按照最大似然函数比通过高斯定理绘出在染色体上最大可能排列的位置。
2 结果与分析
2.1 水稻直链淀粉含量QTLs在染色体上的分布
利用BioMercator 2.1软件中tools-maps projection选项进行水稻直链淀粉含量QTL的映射。结果显示,有32个群体定位的131个QTLs得到映射,可构建水稻直链淀粉含量QTL的一致性图谱(见图1)。一致性图谱结果显示,水稻直链淀粉含量QTL分布在第3、6、7、8和9染色体上,其中第3和第6染色体上比较多,且第6染色体上的QTL成簇出现,有部分重叠,其他染色体上较少。总体上,水稻直链淀粉含量QTL在第3、6、7、8 和9染色体上分布不均匀,较为分散。
2.2 水稻直链淀粉含量QTLs的元分析
利用BioMercator 2.1软件的Meta-analysis选项进行水稻直链淀粉含量QTL的元分析。结果中,每次分析得出的5个模型中以AIC值最小的模型为准来确定“真实”QTL。将实际操作中的原始图谱中单一QTL映射产生的“一致性”QTL结果剔除,结果见表2。表2结果显示,分别在第1、第2、第3、第6、第7、第8、第9和第10染色体上,共有30个“真实”QTL被发现,且得到的QTL的图距都较小,最小的为1 cM,最大的为23.8 cM。第1染色体上检测到6个“真实”QTL,第3、第6和第7染色体上各检测到4个“真实”QTL,第2、第8、第9和第10染色体上各检测到3个“真实”QTL。其中,第8染色体上检测到2个图距分别为1和2.8 cM的“真实”QTL,分别位于52.2和58.1 cM处,标记区间为RM483-RM458和RM547-RM72,置信区间分别为(52.2-53.2)和(58.1-60.9)。第7染色体上的位于115.3 cM处检测到标记区间为RM172-RM428的“真实”QTL,图距仅为1.3 cM。
3 讨论
3.1 水稻直链淀粉含量QTL整合的重要实践意义
随着生物技术手段的发展及对水稻直链淀粉含量的研究深入,国内外研究者进行大量关于水稻直链淀粉含量QTL的基因定位,以期将其应用于水稻品质的分子辅助育种中,但由于作图群体小,群体类型和遗传背景不同,图谱精度低等原因,所定位QTL区间跨度大,标记的有效性降低,难于应用于实践中。而基于元分析的QTL整合,建立数学模型进行QTL优化,将置信区间缩小,本研究中置信区间最小值为1 cM,减小QTL误差,使QTL精度和有效性提高,可解决传统QTL分析局限。
本研究定位于第6染色体染色体0 cM处的“真实”QTL与原始图谱中QTL位置相近,但图距由原来的51.4缩小至2.2 cM;位于第3染色体0 cM处的“真实”QTL与原始图谱中QTL位置相同,图距由原来的40缩小至11 cM;位于第1染色体134.8 cM处的“真实”QTL与原始图谱中QTL位置相同,图距由原来的19.6缩小至8.4 cM。
3.2 图谱整合对MAS的应用途径
通过图谱整合可获得区间更小的“真实”QTL和性状相关标记,直接应用于分子辅助育种(MAS中。对元分析得到的“真实”QTL和标记区间的基因进行分析,可进一步发掘候选基因,成为图谱整合应用的新途径。吉海莲等对玉米抗丝黑穗病QTL进行整合,得到2个“一致性”QTL,并对其区间序列通过同源比对初步获得4个抗病位置候选基因[39]。高利芳等将201个与大豆株高相关的QTL信息整合并对重演性较好的QTL位点所对应的区段内的基因进行分析,初步筛选出17个与大豆株高相关的候选基因[5]。图谱整合可作为标记开发的新途径。对真实QTL区段内DNA序列进行分析,寻找该区间的简单重复序列,根据其两端的保守序列,利用SSR设计软件设计新的SSR引物,开发出与之连锁的SSR分子标记,为水稻分子辅助育种提供可选择的可靠分子标记,可提高优质水稻品种选育的效率。

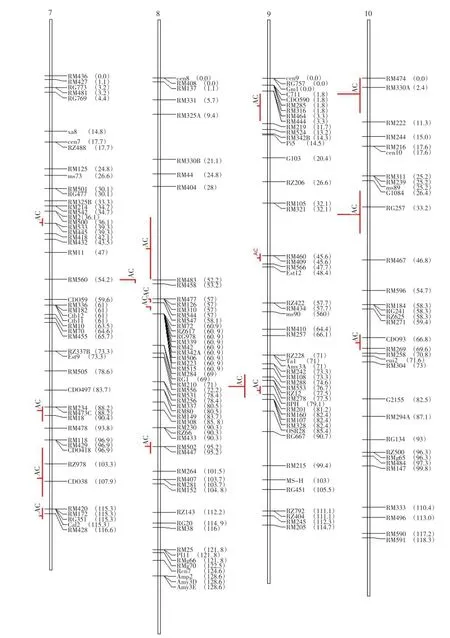
图1 水稻直链淀粉含量QTL元分析整合图谱Fig.1 Consensus map of QTLs of rice amylose content
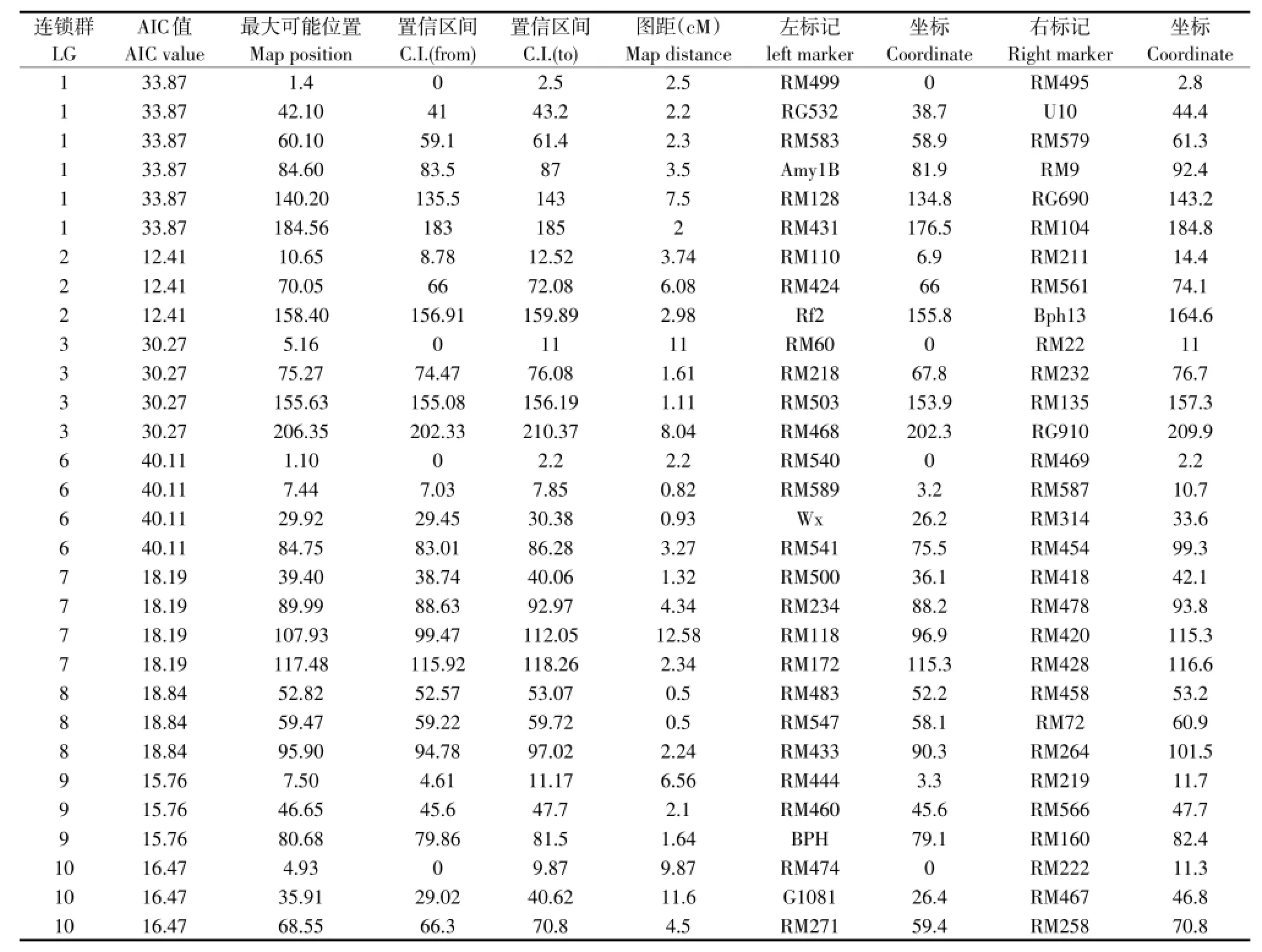
表2 水稻直链淀粉含量QTL元分析结果Table 2 Meta-analysis results of rice amylose content
4 结论
本研究收集来自34个作图群体共141个与水稻直链淀粉含量相关的QTLs信息,并利用Biomer⁃cator 2.1软件将QTLs整合到参考图谱Cornell 2001上,通过元分析方法得到“真实”QTLs及其临近标记和置信区间。结果表明,有32个群体定位的131 个QTLs得到映射,构建水稻直链淀粉含量QTL的一致性图谱,水稻直链淀粉含量QTL分布在第1、2、3、6、7、8、9和第10染色体上,元分析共发现30个“真实”QTL,所得“真实”QTL图距较小,最小图距为1 cM,图距5 cM以下的有9个。
[1]Lanceras J C,Huang Z L.Mapping of genes for cooking and eat⁃ing qualities in Thai jasmine rice(KDML105)[J].DNA Research, 2000(7):93-101.
[2]王茂青.稻米品质性状的QTL定位及遗传分析[D].南京:南京农业大学,2007.
[3]张闻博,蒋洪蔚,李灿东,等.基于元分析的大豆包囊线虫抗性QTL的整合[J].中国油料作物学报,2010,32(1):104-109.
[4]刘硕,罗玲,刘章雄,等.大豆蛋白质含量QTL的“整合”及Overview分析[J].大豆科学,2011,30(1):1-7.
[5]高利芳,郭勇,郝再斌,等.大豆株高QTL的整合“整合”及Overview分析[J].遗传,2013,35(2):215-224.
[6]Darvasi A,Soller M.A simple method to calculate resolving power and confidence interval of QTL map location[J].Behav Genet, 1997,27:125-132.
[7]He P,Li S G,Qian Q,et al.Genetic analysis of rice grain quality [J].Theor Appl Genet,1999,98:502-508.
[8]Aluko G C,Tohme M J.QTL mapping of grain quality traits from the interspecific cross Oryza sativa×O.glaberrima[J].Theor Ap⁃pl Genet,2004,109:630-639.
[9]Tan Y F,Li J X,Yu S B,et al.The three important traits for cook⁃ing and eating quality of rice grains are controlled by a single lo⁃cus in an elite rice hybrid,Shanyou63[J].Theor Appl Genet, 1999,99:642-648.
[10]Fan C C,Yu X Q,Xing Y Z,et al.The main effects,epistatic ef⁃fects and environmental interactions of QTLs on the cooking and eating quality of rice in a doubled-haploid line population[J].The⁃or Appl Genet,2005,110:1445-1452.
[11]Li J M,Xiao J H,Grandillo S,et al.QTL detection for rice grain quality traits using an interspecific backcross population derived from cultivated Asian(O.sativa L.)and African(O.glaberrima S.)rice[J].Genome,2004,47:697-704.
[12]Bao J S,Wu Y R,Hu B,et al.QTL for rice grain quality based on a DH population derived from parents with similar apparent amy⁃lose content[J].Euphytica,2002,128:317-324.
[13]Septiningsih E M,Trijatmiko K R,Moeljopawiro S.Identification of quantitative trait loci for grain quality in an advanced back⁃cross population derived from the Oryza sativa variety IR64 and the wild relative O.rufipogon[J].Theor Appl Genet,2003,107: 1433-1441.
[14]Yoshida S,Ikegami M,Kuze J.QTL analysis for plant and grain characters of sake-brewing rice using a doubled haploid popula⁃tion[J].Breeding Science,2002,52:309-317.
[15]Kepiro J,McClung A M,Chen M H,et al.Mapping QTLs for mill⁃ing yield and grain characteristics in a tropical japonica long grain cross[J].Journal of Cereal Science,2008,48:477-485.
[16]Wan X Y,Wan J M,Su C C,et al.QTL detection for eating quali⁃ty of cooked rice in a population of chromosome segment substitu⁃tion lines[J].Theor Appl Genet,2004,110:71-79.
[17]Wang L Q,Liu W J,Xu Y,et al.Genetic basis of 17 traits and vis⁃cosity parameters characterizing the eating and cooling quality of rice grain[J].Theor Appl Genet,2007,115:463-476.
[18]Li Z F,Wan J M,Xia J F.Mapping of quantitative trait loci con⁃trolling physico-chemical properties of rice grains(Orayza sativa L.)[J].Breeding Science,2003,53:209-215.
[19]Guo Y M,Mu P,Liu J F,et al.QTL mapping and Q×E interac⁃tions of grain cooking and nutrient qualities in rice under upland and lowland environments[J].Journal of Genetics and Genomics, 2007,34(5):420-428.
[20]包劲松,何平,李仕贵,等.异地比较定位控制稻米蒸煮食用品质的数量性状基因[J].中国农业科学,2000,33(5):1-7.
[21]王士梅.氮素对稻米主要品质性状的影响及QTLs定位研究[D].合肥:安徽农业大学,2006.
[22]郑希.稻米蒸煮和营养品质性状的胚乳和母体QTL定位[D].杭州:浙江大学,2007.
[23]田瑞.稻米蒸煮与食味品质,米饭特性相关性状的QTL定位及香味基因的精细定位[D].武汉:华中农业大学,2006.
[24]张坚勇.水稻品质性状的稳定性分析[D].南京:南京农业大学, 2003.
[25]历建萌.稻米主要品质性状的QTL定位及其相关性研究[D].南京:南京农业大学,2001.
[26]朱昌兰,江玲,张文伟,等.稻米直链淀粉含量和胶稠度对高温耐性的QTL分析[J].中国水稻科学,2006,20(3):248-252.
[27] 夏玉梅.稻米品质性状的QTL定位及遗传分析[D].南京:南京农业大学,2008.
[28]翁建峰,万向元,吴秀菊.利用CSSL群体研究稻米AC和PC相关QTL表达稳定性[J].作物学报,2006,32(1):14-19.
[29]唐绍清.稻米蒸煮和营养品质性状的QTL定位[D].杭州:浙江大学,2007.
[30]邵高能,唐绍清,焦桂爱.稻米蒸煮品质性状的QTL定位[J].中国水稻科学,2009,23(1):94-98.
[31] 范楚川.稻米品质性状的遗传基础研究以及粒长基因GS3的图位克隆[D].武汉:华中农业大学,2006.
[32]许勇.杂交稻协优57食味品质改良及水稻直链淀粉含量相关QTL定位的研究[D].合肥:安徽大学,2010.
[33]黄祖六,谭学林,Tragoonrung S,等.稻米直链淀粉含量基因座位的分子标记定位[J].作物学报,2000,26(6):777-782.
[34] 李荧.水稻蛋白质含量近红外模型建立与水旱条件下稻米直链淀粉与蛋白质含量的QTL分析[D].武汉:华中农业大学, 2006.
[35]何颖.水稻淀粉合成相关基因在决定稻米蒸煮和食味品质中的功能分析[D].扬州:扬州大学,2006.
[36]晁园.利用重组自交系群体定位水稻品质相关性状QTL[D].武汉:华中农业大学,2010.
[37] 林红梅.水稻籽粒主要品质性状的QTL定位分析[D].福州:福建农林大学,2009.
[38] 赵守环.水稻RFLP-SSR连锁图谱的构建和稻米品质的QTL定位[D].福州:福建农林大学,2003.
[39]吉海莲,李新海,谢传晓,等.基于元分析的抗玉米丝黑穗病QTL比较定位[J].植物遗传资源学报,2007,8(2):132-139.
Study on integration of QTLs related to amylose content of rice
Amylose content of rice was an important index for rice breeding.A total of 141 QTL information related to amylose content of rice from 34 mapping populations were collected.These QTLs and their common markers were integrated with the reference map Cornell 2001 by Biomercator 2.1. The real QTLs were calculated by the method of meta-analysis.Moreover,a consensus map of rice QTLs conferring amylose content was constructed.Totally,30 real QTLs and their linked markers were found from 1,2,3,6,7,8,9,and 10 chromosomes.This research offers a basis for fine mapping, map-based cloning and marker-assisted breeding of rice quality breeding.
rice;amylose content;QTL integration;meta-analysis
S511
A
1005-9369(2014)03-0008-07
时间2014-3-21 9:09:00 [URL]http://www.cnki.net/kcms/detail/23.1391.S.20140321.0909.004.html
李修平,马文东,王仲,等.水稻直链淀粉含量QTL图谱整合研究[J].东北农业大学学报,2014,45(3):8-14.
Li Xiuping,Ma Wendong,Wang Zhong,et al.Study on integration of QTLs related to amylose content of rice[J].Journal of Northeast Agricultural University,2014,45(3):8-14.(in Chinese with English abstract)
2013-08-26
佳木斯大学校级人才培养计划项目(RC2010-025)
李修平(1981-),女,副教授,博士,研究方向为植物遗传育种与生物技术。E-mail:lixiuping200@163.com
LI Xiuping1,MA Wendong2,WANG Zhong1,GUO Yong1,LIU Fangming1,WU Hengmei1(1.School of Life Sciences,Jiamusi University,Jiamusi Heilongjiang 154007,China;2.Jiamusi Rice Institute of HeilongjiangAcademy ofAgricaltural Sciences,Jiamusi Heilongjiang 154026,China)
