三色堇小孢子发生和雄配子体发育的解剖学研究
2014-01-14王晓磊徐永清王莹胡宝忠
王晓磊,徐永清,王莹,胡宝忠*
(1.东北农业大学生命科学学院,哈尔滨 150030;2.哈尔滨学院体育学院,哈尔滨 150086;3.哈药集团技术中心,哈尔滨 150025)
三色堇小孢子发生和雄配子体发育的解剖学研究
王晓磊1,2,徐永清1,王莹3,胡宝忠1*
(1.东北农业大学生命科学学院,哈尔滨 150030;2.哈尔滨学院体育学院,哈尔滨 150086;3.哈药集团技术中心,哈尔滨 150025)
运用石蜡切片法对三色堇(Viola tricolor L.)小孢子发生及雄配子体形成进行研究。结果表明,三色堇成熟的花药有4个小孢子囊,花药壁发育为基本型,共6~7层,从外至内依次是表皮、药室内壁、中层和绒毡层,绒毡层发育类型为腺质绒毡层。小孢子母细胞减数分裂为连续型,四分体呈四面体型排列。成熟花粉为2-细胞型或3-细胞型,在一个横切面上最多具有5个萌发孔。
三色堇;小孢子发生;雄配子体
三色堇(Viola tricolor L.)属堇菜科堇菜属多年生草本花卉。原产欧洲,我国于20世纪20年代初引进,全国各地均有栽培[1],可用作庭院、花坛、景区栽培及盆栽。三色堇可全草入药,具清热解毒、散瘀、止咳等功能,用于治疗小儿瘰疬、咳
嗽及呼吸道炎症等疾患[2]。三色堇中含芸香甙、类胡萝素类、挥发性精油、花青素等组分,是生产化妆品重要活性成分。近年来,学者对三色堇研究主要集中在栽培育种[3-5]、抗性生理[6-10]、亲缘关系[11-12]和性状分析[13-15]等方面。对三色堇花粉形态和培养[16-19]、组织培养[20-21]、核型分析等[22-33]方面开展研究。但有关生殖生物学研究尚无报道,植物生殖成功是物种繁衍和生存保证,本文利用光镜观察三色堇小孢子发生和雄配子体发育,以期为三色堇在生殖生物学和发育生物学研究奠定理论基础。
1 材料与方法
1.1 材料
试验材料为东北农业大学苗圃三色堇。
1.2 方法
采样时间为2011~2012年5~10月。用手术刀切掉花梗后,把不同发育阶段的三色堇花芽及开放的花蕾立即固定在固定液中,固定液为FAA [70%乙醇90 mL,冰醋酸5 mL,福尔马林(38%甲醛)5 mL],然后带回实验室。在室温下保存,试验材料经酒精逐级脱水,二甲苯置换,包埋于石蜡中。采用常规石蜡切片法制片,切片厚度为6~8 μm,加拿大树胶封片,染色方法为爱氏苏木精整染法和番红-固绿双染法,光学显微镜下观察花药发育过程。
2 结果与分析
2.1 花药壁的发育
三色堇开花早,花期长,花为两性花,花瓣5枚;雄蕊5个,花药围绕子房排列成一环,下方的两个雄蕊基部有距状的蜜腺,属于雄蕊蜜腺,为花药下端药隔的棒状延伸物。成熟花药具有4个花粉囊。在横切面上,三色堇初期的花药为椭圆形,最外面是一层表皮,在表皮下是一群分裂活跃排列紧密的细胞。随着花药的发育,在花药四个角隅处的细胞分裂较快,分化出孢原细胞。孢原细胞经过一次平周分裂,形成两层细胞,外层的细胞为初生壁细胞,内层的细胞为初生造孢细胞。初生壁细胞进行垂周分裂和平周分裂,形成环绕初生造孢细胞的一层细胞。在经过两次连续的平周分裂后,此层细胞分化成4层细胞。从外到内依次是药室内壁、中层和绒毡层细胞。因此,三色堇成熟的花药壁共6~7层(见图版Ⅰ-1,2),从外到内分别是表皮1层,药室内壁1层,中层2~4层,绒毡层1层,其发育方式为基本型。分化初期时花药壁的几层细胞在形态上差别不大,随着花药的发育,药壁各层细胞形态结构发生一系列变化。表皮细胞最初是1层近等径的细胞,细胞质浓厚,细胞核大,随着花药的发育,表皮细胞逐渐变得扁平,在细胞外壁上具有明显的角质层。当花药成熟时,表皮细胞仍然存在于最外层,但形态变得不规则并且逐渐瓦解,在整个发育过程中,表皮细胞只进行垂周分裂。
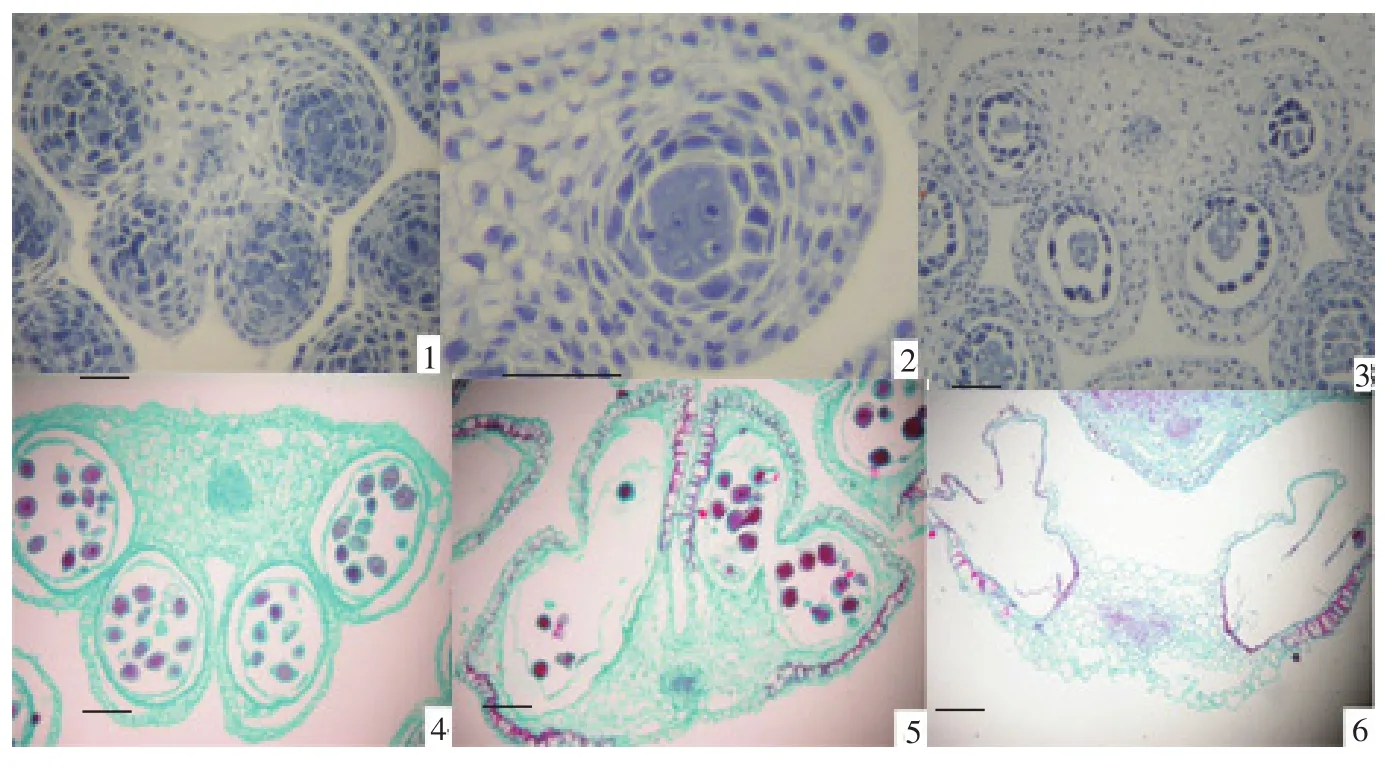
图版Ⅰ 花药壁的发育PlateⅠ Development of anther wall
药室内壁由1层细胞构成,在形成小孢子母细胞阶段,药室内壁的细胞壁上开始形成不同程度的带状径向加厚;单核花粉粒时期,细胞由近正方形变为径向延长;在花药成熟时,细胞呈栅栏状排列。药室内壁细胞在加厚时,在两个花粉囊交接处留下一条不发生加厚的区域,当花药成熟时,在此处开裂散出花粉粒。
中层位于药室内壁的内侧,由2~4层细胞构成,在细胞形态上与药室内壁无明显差别。小孢子母细胞阶段,中层细胞变扁平,然后逐渐瓦解并被吸收。当花药成熟时中层细胞完全消失。
绒毡层细胞是由初生壁细胞发育成的,属腺质绒毡层。刚分化形成的绒毡层细胞是单核的,随着花药的发育,绒毡层细胞进行有丝分裂,但在分裂过程中不形成细胞板,使形成的两个二倍体子核保留在一个细胞内,使细胞具有双核;其中有的继续分裂成具多核的细胞。当绒毡层发育到最高程度时,其细胞比药壁的其他细胞大,细胞质变浓厚,液泡变小并且变少,细胞排列紧密为二核或多核的细胞,在花药横切面上看呈近方形且排列成规则的环形。绒毡层表面能分泌各种物质为小孢子发育提供营养。小孢子发育阶段,绒毡层开始表现出明显的解体迹象,细胞体积变小且扁平(见图版Ⅰ-3);当花粉成熟后,绒毡层细胞完全自溶。因此成熟的花药壁仅剩下表皮细胞和药室内壁两层细胞(见图版Ⅰ-4,5)。
2.2 小孢子的发生
孢原细胞(见图版Ⅱ-1)经过平周分裂形成初生壁细胞和初生造孢细胞(见图版Ⅱ-2,3),随着药壁的分化,初生造孢细胞进行有丝分裂形成多个排列紧密的次生造孢细胞(见图版Ⅱ-4),次生造孢细胞呈四边形,细胞核大且细胞质浓厚,次生造孢细胞长大、变圆直接发育成小孢子母细胞(见图版Ⅱ-5)。小孢子母细胞的数量相对较少,在一个横切面上最多不超过10个。小孢子母细胞为圆形,细胞体积大,细胞核大且核仁明显,细胞质浓厚但无液泡。小孢子母细胞不断长大并进入减数分裂期(见图版Ⅱ-6,7),经过两次减数分裂形成四分体。观察表明,在减数分裂时,小孢子母细胞第一次减数分裂形成两个有细胞壁的二分体(见图版Ⅱ-8)。经过短暂的分裂间期,进行第二次减数分裂并形成四面体型的四分体(见图版Ⅱ-9)。因此,小孢子母细胞减数分裂过程所发生的胞质分裂为连续型。
2.3 雄配子体的发育
新形成的小孢子四分体被胼胝质壁所包围,随着花药的发育胼胝质壁消失,小孢子四分体分散开,刚散开的小孢子形状近球形,细胞质浓厚,无液泡,位于中央的细胞核较大,外壁加厚明显,发育成单核花粉粒(见图版Ⅱ-10)。随着小孢子体积的不断增大,细胞质发生液泡化,在小孢子的中央逐渐形成一个大液泡,细胞质呈一薄层贴着细胞壁,细胞核也从中央移到细胞的一侧,此时期为单核靠边期(见图版Ⅱ-11)。接着单核靠边的小孢子进行一次不均等的有丝分裂,形成两个大小悬殊的细胞,大的是营养细胞,包含原来小孢子的大部分细胞质和液泡;小的是生殖细胞,只有少量的细胞质,细胞呈凸透镜状,靠近花粉壁。此后,生殖细胞脱离花粉的壁,游离在营养细胞的细胞质中,有的生殖细胞进行一次有丝分裂形成两个精子。因此三色堇成熟的花粉粒以2-细胞型为主(见图版Ⅱ-12),兼有少量的3-细胞型花粉(见图版Ⅲ-1~3)。
在花药的切片中观察到,三色堇的5个花药在发育初期具有不同步性。位于上方的3个花药发育早于下方的两个花药(见图版Ⅲ-4,5),当下方的两个花药四分体刚分开不久,小孢子细胞质液泡化,形成一薄层贴着细胞壁,在中央逐渐形成一个大液泡,细胞核也从中央移到一侧时,上方的3个花药已发育到2-细胞型花粉时期。
2.4 花粉粒的形态结构
成熟花粉粒的特点是花粉粒数量少且体积大,花粉内的营养核和生殖细胞较大。随着花药发育,花粉粒内积累大量淀粉粒和脂肪,使花粉粒呈球形,在一个平面上最多能看到5个萌发孔(见图版Ⅲ-3)。
2.5 小孢子及雄配子体发育与绒毡层细胞发育的关系
绒毡层细胞的细胞质浓厚,含较多的RNA和蛋白质,还有丰富的油脂和类胡萝卜素等,能为小孢子的发育提供营养。现将小孢子及雄配子体与绒毡层细胞发育的关系归纳为表1。
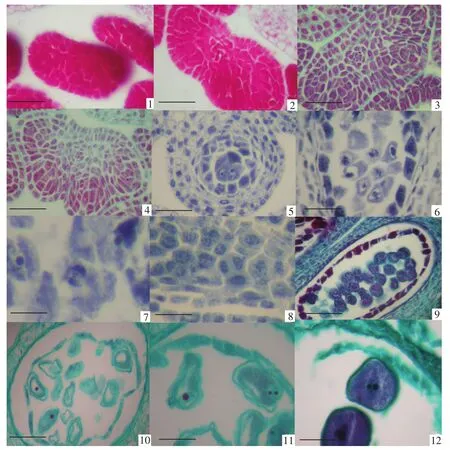
图版Ⅱ 小孢子的发生与雄配子体的发育PlateⅡ Microsporogenesis and male gametophyte development
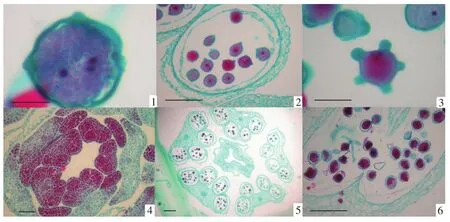
图版Ⅲ 花粉和不同步发育的花药PlateⅢ Pollen and different developments of anther

表1 小孢子及雄配子体与绒毡层细胞发育的相互关系Table 1 Interrelation of development of microspore and antheridium with tapetum
3 讨论与结论
一般植物的花瓣等大,排列整齐,花药发育具有同步性。本研究在花药切片中发现,三色堇5个花药开始发育过程不同步。位于上方的3个花药发育早于下方两个花药,当下方两个花药四分体刚分开不久,小孢子细胞质液泡化,形成一薄层贴着细胞壁,在中央逐渐形成一个大液泡,细胞核也从中央移到一侧时,上方3个花药已发育到2-细胞型花粉时期。这种现象在其他植物中也有发现[24-26]。胡适宜等认为,同一花粉囊的小孢子母细胞之间存在细胞质联络通道,使整个小孢子囊的小孢子母细胞形成一个合胞体,有助于细胞间物质均衡分布,也保证减数分裂具有很高同步性[27]。然而对同一花中不同花药发育的不同步现象,潘开玉等认为,这是植物延长传粉期进而适应环境的现象[28]。三色堇花药发育不同步现象,除为了延长传粉期,还可能和三色堇花的形态特征有关。下方的两个花药由于位于子房的下方,发育过程中受到子房的挤压;在下方的两个雄蕊基部有距状的蜜腺,在这两方面因素共同作用下,造成三色堇花药发育不同步。是否还有其他因素,有待进一步研究。孙坤、李方等观察到堇菜属的花粉为长球形[16,18],近球形或球形,三色堇的花粉为5孔沟。本研究也发现三色堇的花粉为球形,在一个平面上最多能看到5个萌发孔,这与孙坤等[16]研究结果基本一致。
三色堇的雄蕊具有4个花粉囊,花药壁由表皮、药室内壁、中层和绒毡层组成,中层位于药室内壁的内侧,由2~4层细胞构成,四分体呈正四面体形,这些特征与韦仲新等[29]对堇菜科植物花药特征的描述基本一致。韦仲新等总结堇菜科植物绒毡层为分泌型,细胞为双核的细胞,小孢子母细胞减数分裂为同时型,花粉粒为2-细胞型。而本试验中观察到,三色堇的绒毡层为腺质绒毡层,细胞有的为双核细胞,有的为多核细胞[29]。小孢子母细胞减数分裂为连续型,花粉粒以2-细胞型为主,兼有少量的3-细胞型,花药壁的发育方式为基本型。这些特征,可丰富堇菜科植物生殖生物学研究。
[1]郭兆武,萧浪涛.6-BA和GA3促进三色堇再生的研究[J].湖南农业大学学报:自然科学版,2002,28(6):499-501.
[2]姚振生,傅骞峰,曹岚.江西省堇菜属药用植物资源及利用[J].江西科学,2001,19(2):104-107.
[3]陶玉祥.三色堇日光温室杂交制种技术[J].种子科技,2012(3): 32-33.
[4]赵宇,牛常青,赵坤.三色堇对粉煤灰-醋糟基质的适应性探究[J].中国农学通报,2012,28(1):171-175.
[5]刘红玉,李方华.酒泉地区温室三色堇杂交制种技术[J].上海蔬菜,2012(3):15-16.
[6]罗玉兰,陆亮,王泰哲.本地和荷兰三色堇抗寒性的比较[J].植物生理学通讯,2001,37(1):27-28.
[7]尤扬,贾文庆.NaCI胁迫对三色堇SOD、POD及叶绿素含量的影响[J].北方园艺,2012(6):64-66.
[8]杜晓华,刘会超,袁少寒,等.三色堇花期光合生理生态特征[J].西南林业大学学报,2011(3):7-12.
[9]朱永兴,韩春叶,白志怀.干旱胁迫对盆栽三色堇生理指标的影响[J].河南农业科学,2013,42(8):109-111.
[10]彭华婷,高悦,杜红梅,等.高温胁迫对大花三色堇幼苗相关生理指标的影响[J].上海交通大学学报:农业科学版,2012,30 (6):66-70.
[11]王涛,徐进,张西西,等.43份三色堇、角堇材料亲缘关系的SRAP分析[J].中国农业科学,2012,45(3):496-502.
[12]杜晓华,刘会超,刘孟刚,等.33个三色堇品种遗传差异的初步分析[J].西北林学院学报,2010,25(4):78-82.
[13]杜晓华,刘会超,姚连芳.三色堇观赏性状的主成分分析[J].西北农业学报,2011,20(6):136-140.
[14]杜晓华,刘会超,朱方方,等.基于主成分的三色堇聚类分析[J].北方园艺,2011(5):180-184.
[15]卢兴霞,胡惠蓉,王健.花三色堇自交系主要数量性状配合力分析[J].河北农业大学学报,2013,36(1):11-15.
[16]孙坤,王漪,王晓东,等.国产堇菜属花粉形态的研究[J].植物研究,1994,14(1):81-87.
[17]李春燕,李清斌,程振,等.大花三色堇和三色堇花粉离体萌发的培养基研究[J].北方园艺,2011(3):54-56.
[18]李方,尹祖棠.堇菜属植物花粉及叶表皮超微形态多样性的研究[J].中国农业大学学报,2000,5(3):6-12.
[19]周玉敏,徐自警.三色堇花粉生活力的测定[J].中国林副特产,2012,4,28-29.
[20]姚立平.三色堇组织培养技术研究[J].辽宁林业科技,2012(2): 26-27.
[21]王振民,付强,赵兵.三色堇组培快繁技术研究[J].辽宁农业科学,2011(1):43-45.
[22]穆金艳,杜晓华,刘会超.大花三色堇的核型分析[J].东北林业大学学报,2013,41(7):75-78.
[23]何丽君,周小燕.三色堇染色体核型分析[J].内蒙古农业大学学报,2007,28(6):200-204.
[24]曹清河,陈劲枫,钱春桃,等.黄瓜花粉母细胞减数分裂及其雄配子体发育的细胞学研究[J].西北植物学报,2004,24(9): 1721-1726.
[25]卢立娜,贺晓,易津,等.华北驼绒藜大小孢子的发生及雌雄配子体发育过程的解剖学研究[J].西北植物学报,2008,28(7): 1319-1325.
[26]高建平,贺晓,郭春燕,等.蒙古莸小孢子发生和雄配子体发育的研究[J].西北植物学报,2009,29(6):1149-1154.
[27]胡适宜.被子植物学生殖生物学[M].北京:高等教育出版社,2005:33-34.
[28]潘开玉,温洁,周世良.石香薷(唇形科)的胚胎学研究[J].植物学报,1997,39(2):111-116.
[29]韦仲新,金巧军.西域旌节花的雌雄配子体发育及其系统学启示[J].云南植物研究,2002,24(6):733-742.
Anatomical studies on microsporogenesis and development of male gametophyte in pansy
WANG Xiaolei1,2,XU Yongqing1,WANG Ying3,HU Baozhong1
(1.School of Life Sciences,Northeast Agricultural University,Harbin 150030,China;2.School of Physical Education,Harbin University,Harbin 150086,China;3.Harbin Pharmaceutical Group R&D Center,Harbin 150025,China)
The microsporogenesis and development of male gametophyte of pansy(Viola tricolorL.) were studied with the method of general paraffin section technology.The results showed that the mature anther of pansy had four microsporanges.The growth of the anther wall with six or seven layers belonged to tetrahedron type.The layers from the inside out of anther wall were epiderm,endothecium,middle layer and secretory glandular tapetum of pansy successively.The cytokinesis division of the microspore mother cell was successive type and the tetrad was tetrahedron type.Mature pollen was 2-celled or 3-celled which had five germinal pores.
pansy;microsporogenesis;male gametophyte
S681.9
A
1005-9369(2014)04-0060-06
2013-09-29
黑龙江省自然科学基金(C2008049);国家国际科技合作项目子课题(2013DFR30270)
王晓磊(1978-),女,讲师,博士,研究方向为植物分子生物学。E-mail:wxlei9234@163.com
*通讯作者:胡宝忠,教授,博士生导师,研究方向为植物分子生物学。E-mail:bzhu@neau.edu.cn
时间2014-4-21 13:25:43[URL]http://www.cnki.net/kcms/detail/23.1391.S.20140421.1326.036.html
王晓磊,徐永清,王莹,等.三色堇小孢子发生和雄配子体发育的解剖学研究[J].东北农业大学学报,2014,45(4)∶60-65.
Wang Xiaolei,Xu Yongqing,Wang Ying,et al.Anatomical studies on microsporogenesis and development of male gametophyte in pansy[J].Journal of Northeast Agricultural University,2014,45(4)∶60-65.(in Chinese with English abstract)
