退化华北落叶松人工林林隙更新特征
2014-01-03董伯骞黄选瑞徐学华张玉珍
董伯骞,黄选瑞,徐学华,张玉珍
(1.北京林业大学 林学院,省部共建森林培育与保护教育部重点实验室,北京 100083;2. 河北农业大学 林学院,河北 保定 071000)
退化华北落叶松人工林林隙更新特征
董伯骞1,黄选瑞2,徐学华2,张玉珍2
(1.北京林业大学 林学院,省部共建森林培育与保护教育部重点实验室,北京 100083;2. 河北农业大学 林学院,河北 保定 071000)
通过对冀北山地退化华北落叶松人工林52个林隙的调查,研究了林分的林隙特征和实施封禁后林隙开敞度对林下更新的影响。研究区域林隙分布广泛,且大多数林隙的开敞度较小。封禁3年后,天然更新密度显著提高,树种多样性增加;但更新密度仍然很低,更新群落主要由华北落叶松和山杨组成。总体上,封禁后林隙内更新密度高于林冠下,在开敞度为0.5~0.6的林隙内更新密度最大,华北落叶松和山杨分别在开敞度0.4~0.5和0.7~0.8的林隙内生长状况最好。人工栽植的油松和樟子松分别在开敞度0.4~0.6的林隙中央和开敞度0.6~0.8的林隙边缘生长最好。随时间延长林隙和林隙内位置对人工更新高生长的存进作用趋于明显。单纯封禁促进了天然更新,但作用有限,因此人工更新很有必要。
华北落叶松;退化;近自然经营;封禁;林隙特征;更新
华北落叶松Larix pricipis-rupprechtii是中国华北地区森林经营和人工造林的主要树种之一。在河北北部有大面积的华北落叶松人工纯林,由于土地贫瘠、经营粗放、樵采和放牧等原因,部分成为低质低效林。如何对退化的华北落叶松林施加人工干扰,将退化林分逐步改造成复层异龄混交林,实现林分的近自然化经营是当前亟待解决的问题。利用林隙,促进天然更新和人工引入乡土树种是人工林近自然化改造最普遍的方式[1-2]。落叶松人工林具有明显的自然化特征,只要种源存在落叶松人工林内,阔叶树就会侵入[3]。研究表明对退化森林生态系统封禁会增快林木生长[4],促进林分的天然更新[5],加速养分循环和能量生产能力恢复[6],改善土壤结构和水肥条件,增加树种多样性[7-8]。
研究显示借助人为制造林隙可以减少植物演替时间[9-11],基于林隙的经营技术能有效促森林更新[12-15]。很多研究证实了林隙内人工栽植幼苗的可行性[16-17],并且林隙大小和林隙内位置对栽植幼苗成活和生长具有重要影响[18-21]。目前,对林隙更新的研究主要集中在天然林和天然次生林内林隙对更新规律的影响,尤其是林隙对更新密度、幼苗生长和树种组成的影响,涉及人工林的相关研究很少[22]。因此充分认识实施封禁后林隙对天然与人工更新的影响,有助于加深对封禁措施与林隙耦合作用下退化华北落叶松林恢复过程的认识,优化经营措施,促进林下更新,实现林分近自然经营与生态功能的恢复。本文研究目的是:(1)探究退化林分内的林隙特征;(2)揭示实施封禁后林隙对天然更新树种组成、种群密度、生长状况和数量动态的影响;(3)研究林隙开敞度和林隙内位置对人工更新的影响。
1 研究区概况与研究方法
1.1 研究区概况
研究地区位于河北省承德市木兰林管局,地处河北省最北部的围场县,地理位置为116°32′~118°14′N,41°35′~ 42°40′E,位于北京以北 340 km,海拔1 011~1 230 m;属于北温带大陆性燕山山地季风气侯,年平均气温3℃,最热月(7月) 平均气温20.7℃ ,最冷月( 1月) 平均气温-13.2℃,年均降水量为445 mm,多集中于7~9月。年蒸发量1 462.9~1 556.8 cm。
试验地选择木兰林管局龙头山林场道坝子营林区15林班140小班(图1),坡度20°,坡向西北,海拔1 210 m。研究对象为1972年人工营造的华北落叶松人工纯林。封禁前林木平均高10.4 m,平均胸径13.6 cm,郁闭度0.7,平均密度为850株/hm2。由于土层薄与人畜危害相耦合,造成林分结构差、林下植被稀少,导致林分内水土流失严重,侵蚀沟大量存在。到实施改造前的华北落叶松林生长状况极差,乔木落叶松由于过度樵采仅剩下最上部三分之一的枝叶。林内优势草本为:乌苏里苔草Carex ussuriensis、龙牙草Agrimonia pilosa、细裂叶蒿Artemisia tanacetifolia、草地老鹳草Geanium daharicu、早熟禾Poa pratensis、鹅绒萎陵菜Potentilla anserina和小红菊Dendranthema chanetii等;主要灌木有柔毛绣线菊Spiraea pubescens、美丽胡枝子Lespedeza formosa。土壤为棕壤。
图1 试验地位置Fig. 1 Location of the study area
1.2 经营措施
在2007年初对试验区用铁丝网封禁,在现有的林隙中,设置3个林冠空隙开敞度等级(0.3~0.5、0.5~0.7和0.7~0.9),每个等级6个林隙,共18个林隙;每个开敞度等级的3个林隙内人工栽植油松Pinus tabulaeformi和另3个林隙栽植樟子松Pinus sylvestrisvar.mongolica,并在林冠下栽植油松和樟子松。栽植时间为2007年5月,栽植密度都为1.5 m×1.5 m(300株/亩),2年生营养杯苗,挖穴载苗,穴直径40~50 cm,深度20 cm。
1.3 研究方法
采用样带调查法测算林隙特征,于2007年8月进行。从坡下林缘向上走50 m,从起点沿罗盘仪所指的方向,从坡下向梁顶行进,随机用测绳设置两条相隔100 m的样带。从样带起点开始调查每个林隙,记录被样带穿过的林隙的个数和行进距离。目前一般认为林隙的林冠空隙(CG)面积下限为4 m2,但人工林中株行距较大与天然林中情况不同,若下限为4 m2就可以认为人工林中几乎没有林下环境,将全部由林隙组成,因此根据实际情况,以大约4颗株乔木树干间面积(≈16 m2)的两倍30 m2设定为扩展林隙(EG)最小面积,并结合落叶松冠幅将CG的面积下限设定为10 m2。两条样线在林中共穿行1 068 m,遇到52个林隙;样带中心线共穿过其中32个林隙。记载林隙的形状、大小和形成原因。
应用等角八边形法计算林隙面积,分别计算林隙的林冠空隙面积(ASCG)和扩展林隙面积(ASEG)。
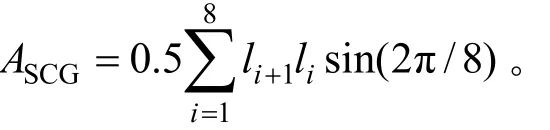
式中l是林窗中心到林窗边缘的水平距离,ln+1=l1,ASCG为林隙面积 。
计算林隙开敞度和EG形状指数。CG开敞度DCG=林隙平均直径(dMD)/边界木平均高(H),EG开敞度DEG=林隙平均直径(dED)/边界木平均高(H)。EG形状指数计算公式如下:

式中L为EG周长,A为EG面积,如果I越接近1,扩展林隙的形状就越近似圆形。
在2007年(封禁前夕)和2010年(封禁3年后)调查天然更新的密度、树种组成、高度和基茎。对于人工栽植的苗木,2008、2009和2010每年秋季调查其生长状况,栽植油松和樟子松的每个林隙内从林隙中央、南部边缘和北部边缘各抽取5株苗木,每一树种每个CG开敞度等级分别测量45株,林冠下抽取油松和樟子松各100株苗木,测量苗高、基茎和年高生长量。
2 结果与分析
2.1 林隙特征
2.1.1 林隙面积比例
林隙的线密度为29.03个/km。在1 068 m的线段上,处于EG中的线段总长为247.3 m,则EG在整个林分中所占比例平均为(247.3/1068)×100%=23.16%,即理解为EG面积在林分面积中的比例。在所调查的52个林隙中,CG和EG的平均面积分别为ASCG=57.88 m2和98.31 m2。CG和EG的平均面积相对比例为ASCG/ASEG=58.88%,可得出CG在整个林分中的相对面积比例为23.16%×58.88%=13.64%。
2.1.2 林隙形状与开敞度
由表1可知,大多数林隙的EG形状指数变化在1.0~1.2之间。指数大于1.2的林隙占总数的36.54%,说明一部分林隙的形状与圆形相差很远。因为在人工林中林木栽植时基本呈均匀分布,所以当林冠层一株或几株林木死亡后形成的空隙往往有明显的折角并更近似于矩形。

表1 EG形状指数IEG分布Table 1 Distribution of shape index
CG开敞度DCG大多为0.3~0.8(表2),占所调查林隙总数的90.38%;大于1.0的很少,但面积比例高达56.59%。EG开敞度DEG大多为0.6~2.0(表3),占所调查林隙总数的82.69%;大于2.0的林隙只有2个,但面积比例高达32.29%。
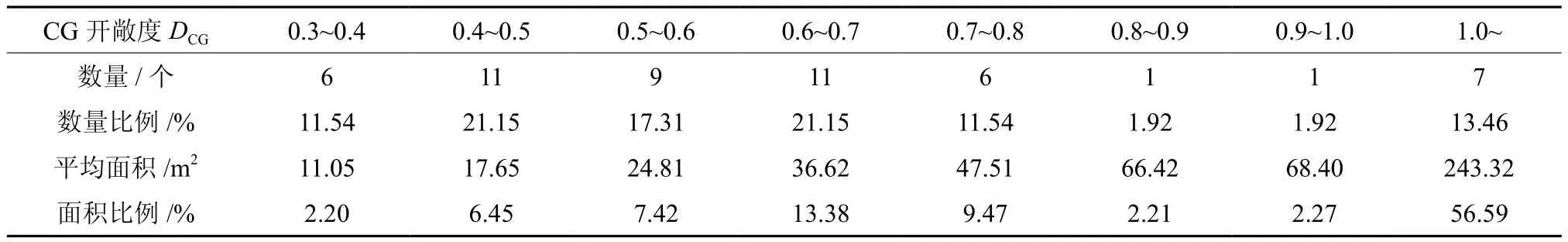
表2 CG开敞度DCG分布Table 2 Distribution of CG open degree
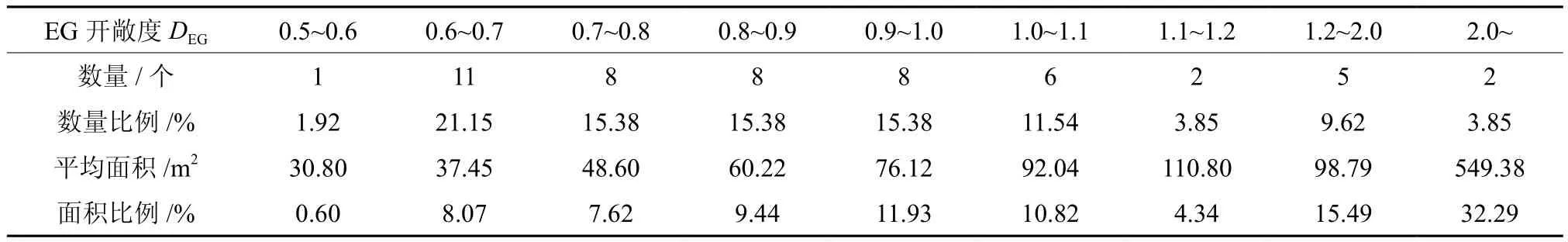
表3 EG开敞度DEG分布Table 3 Distribution of EG open degree
2.1.3 林隙的大小结构
CG的最小面积为10.04 m2,最大面积为671.35 m2,平均面积为57.88 m2,EG的最小面积为30.09 m2,最大面积为774.29 m2,平均面积为98.31 m2。CG的面积主要分布在10~50 m2,10~50 m2的林隙所占面积比例为37.08%;130 m2以上的林隙数量比例只有7.69%,但面积比例高达46.43%(图2a和2c)。EG大多在30~90 m2的范围内,大于400 m2的林隙数量比例占7.69%,面积比例为36.05%(图2b和2d)。

图2 林隙数量和面积分布Fig. 2 Distribution of gaps quantitative and area
2.1.4 林隙形成原因
调查发现,林隙主要形成方式有3种类型:人工造林的苗木死亡所形成的林隙;树木在生长过程中受风折、雪压、旱害、病虫害和雷击影响致死所形成的林隙;盗伐所形成的林隙。由于立地条件较差和缺乏经营,第一种原因是林隙形成的主要原因。林隙年龄最小为2年,是雷击所致,最大的为28年,是幼龄林阶段林木被盗伐后没有补植形成的。
2.2 森林更新
2.2.1 天然更新树种组成
封禁前天然更新树种只有华北落叶松,封禁3年后天然更新群落树种组成包括华北落叶松、山杨Populus davidiana、小叶杨Populus simoniiCarr、榆树Ulmus pumila、白桦Betula platyphyllaSuk、山杏Armeniaca sibirica(L.) Lam和油松Pinus tabuliformis(表4)。在林隙内华北落叶松和山杨数量比例都超过90%;林冠下只有华北落叶松和山杨,数量比例分别为20.45%和79.55%。在不同开敞度的林隙内华北落叶松和山杨的比例变化较大。
2.2.2 天然更新密度
由表5可知封禁3年后更新密度显著增加,所有林隙内更新密度增幅高于林冠下(表5)。在CG开敞度为0.8~1.0的林隙内没有更新,其余林隙内全部更新密度(幼苗+幼树)、幼树和幼苗密度、华北落叶松和山杨的幼苗和幼树密度都大于林冠下;都在CG开敞度为0.5~0.6的林隙内最高,且远高于其它CG开敞度的林隙内,说明更新密度对林隙的开敞度的响应十分敏感。在各个CG开敞度等级的林隙内和林冠下都表现为华北落叶松幼苗密度大于幼树密度,山杨幼苗密度小于幼树密度。
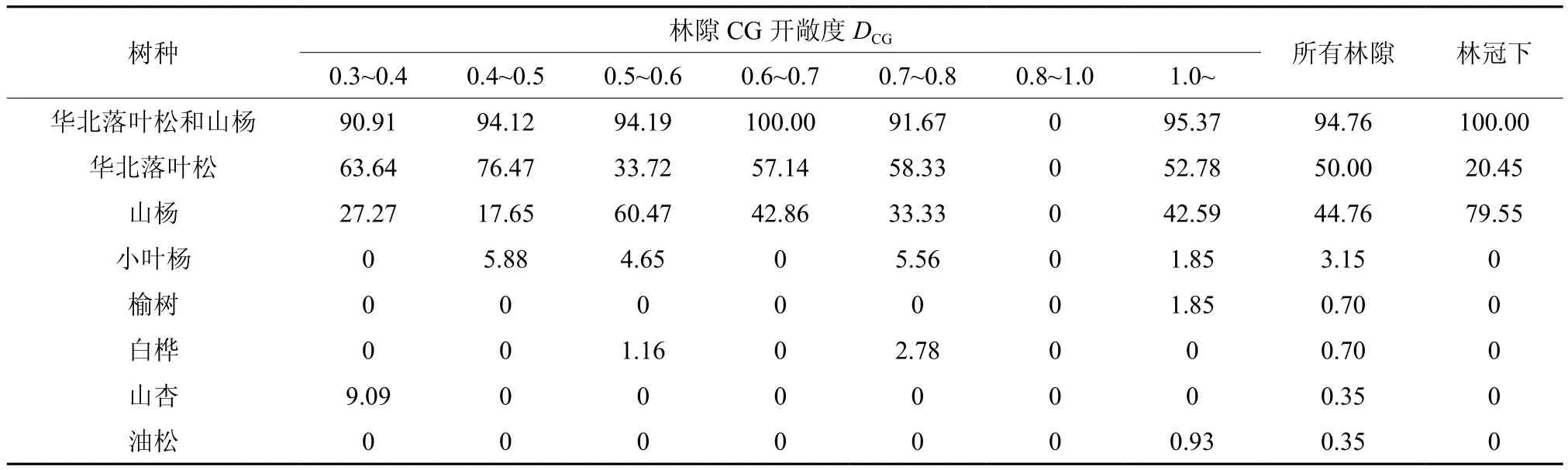
表4 天然更新树种组成Table 4 Species composition of natural recruitment %
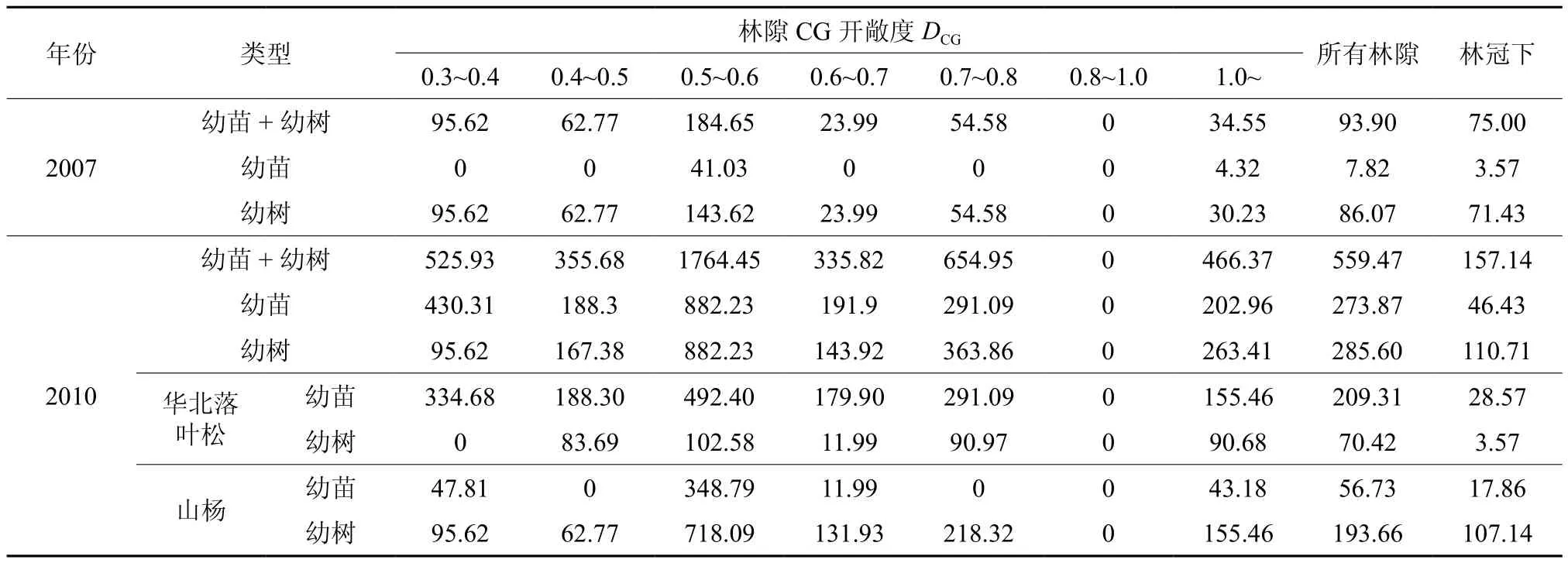
表5 不同CG开敞度林隙内天然更新密度(hm2)Table 5 Natural recruit density in gaps of different CG open degree (hm2)
2.2.3 天然更新的生长状况
所有林隙内全部更新的平均高度小于林冠下,所有林隙内华北落叶松和山杨平均高度与林冠下差异不大(表6)。全部更新、华北落叶松和山杨都表现为在CG开敞度0.3~0.4的林隙内的平均高度显著小于其余林隙内(表6)。华北落叶松平均高度在CG开敞度0.4~0.5的林隙内最高(62.23 cm),在CG开敞度0.4~0.5和大于1.0的林隙内大于林冠下,在其余林隙内小于林冠下;山杨平均高度在开敞度为0.7~0.8的林隙内最大(112.08 cm),0.4~0.5和0.6~0.8的林隙内大于林冠下,其余林隙内小于林冠下(表6)。

表6 不同CG开敞度林隙内外天然更新个体平均高度(cm)Table 6 Average height of natural recruitment in gaps of different CG open degree (cm)
所有林隙内全部更新、华北落叶松和山杨的平均基茎都大于林冠下(表7)。全部更新的平均基茎在CG开敞度为0.3~0.4的林隙内小于林冠下,其余开敞度林隙内大于林冠下(表7)。华北落叶松平均基茎在CG开敞度为0.4~0.5的林隙内最大(13.92 mm),在开敞度为0.3~0.4的林隙内小于林冠下,其余林隙内大于林冠下;山杨平均基茎在CG开敞度为0.7~0.8的林隙内最大(7.64 mm),在CG敞度度为0.3~0.5的林隙内小于林冠下;其余林隙内大于林冠下(表7)。

表7 不同CG开敞度林隙内天然更新个体平均基茎(mm)Table 7 Average basal diameter of natural recruitment in gaps of different CG open degree (mm)
2.2.4 人工更新的生长状况
人工栽植的油松在CG开敞度0.8~1.0的林隙内平均高度最大,在其余林隙内和林冠下基本相当,油松基茎在林隙内与林冠下差异不大;高度和基径都为在开敞度0.4~0.6林隙内边缘<中央,在开敞度0.6~0.8的林隙内边缘>中央,在开敞度0.8~1.0林隙内中央和边缘差异不明显。栽植后第1年,油松年高生长量在开敞度0.4~0.6的林隙内最大,其余林隙内小于林冠下;第2年各位置差异很小;第3年各位置基本一致。随时间延长,在0.4~0.6的林隙内油松年高生长量南北缘趋于一致,中央显著高于边缘;在0.6~0.8林隙内南北边缘趋于一致,且高于中央;在开敞度0.8~1.0林隙内南北边缘与中央趋于一致。

表8 不同CG开敞度林隙中油松平均高度(cm)、基茎(mm)与年高生长量(cm)Table 8 Average height (cm), basal diameter (mm) and current height increment (cm) of Pinus tabulaeformis in gaps of different CG open degree
栽植的樟子松高度和基径都为在CG开敞度0.6~0.8的林隙内最大,在其余林隙内和林冠下差异不显著;高度和基径在开敞度0.4~0.6林隙内都为边缘<中央,在其余林隙内边缘>中央。栽植后第1年,樟子松年高生长量在开敞度0.8~1.0的林隙内最小,其余位置差异不大 ;第2年在开敞度0.4~0.6的林隙内最小,各位置差异很小;第3年为开敞度0.6~0.8的林隙>开敞度0.8~1.0的林隙>林冠下>开敞度0.4~0.6的林隙。随时间延长,在开敞度0.4~0.6的林隙内樟子松年高生长量在南缘最大;在开敞度0.6~0.8和0.8~1.0林隙内边缘略高于中央。

表 9 不同CG开敞度林隙中樟子松平均高度(cm)、基茎(mm)与年高生长量(cm)Table 9 Average height (cm), basal diameter (mm) and current height increment (cm) of Pinus sylvestris var. mongolica in gaps of different CG open degree
3 讨论与结论
相对于北京地区的人工针叶林和本地区的华北落叶松人工林,试验地的林隙面积分布范围更广,出现较大的林隙[23-24],说明林分林相被严重破坏。森林采伐更新管理办法规定:每公顷皆伐迹地应当保留健壮目的树种幼树不少于3 000株或者幼苗不少于6 000株。封禁3年后林隙内和林冠下更新密度都远低于规定中的要求,同时密度也低于华北落叶松天然林内天然更新密度,即该林分40年生时若实施采伐更新密度过低,说明在实施封禁措施的同时人工栽植幼苗很有必要[25-26]。
相比林冠下,虽然林隙内出新了华北落叶松和山杨以外的树种,但前两者所占比例都高于90%。这与试验地周边只分布有华北落叶松纯林缺少其他树种的种源[27]和华北落叶松本身自然更新更新能力较差有关,研究发现林隙对更新树种多样性的增加作用有限[28]。除开敞度为0.8~1.0的林隙没有天然更新外,其余林隙内更新密度都高于林冠下,其他研究表明林隙内通常具有更高的更新幼苗密度[29-31],因为林隙可以提高光照到达林地的可利用性,促进不耐阴种的定居与生长[14]。
华北落叶松为喜光树种,不耐阴种的萌发、存活和生长常与林隙开敞度相关[32]。更新密度在CG开敞度为0.5~0.6的林隙内最大,说明在开敞度高于0.6的林隙内光可利用性的增加对幼苗的积极影响会被其他因素抵消,比如干旱、光合抑制作用和热胁迫[33]。研究发现由于没有足够的时间发育出伸展范围广而且植根于深层土壤的根系系统,木本植物幼苗容易遭受干旱胁迫[34]。华北落叶松幼苗在开敞度0.3~0.4的林隙内幼苗密度较高但无幼树出现,可能是有利于幼苗萌发的生境可能会阻碍幼苗的生长[35]。
华北落叶松冠层木对林隙内更新幼苗的作用可能有两方面:一是林隙边界木通过减少不良气候变化的幅度而对幼苗生长有利;二是林隙边界木的遮蔽作用造成幼苗生长光照不足 , 对幼苗生长不利[36]。林隙内和林冠下油松第1年高生长量为8.47~13.67 cm,与太行山西侧阳坡人工栽植的油松苗基本一致,说明林冠下和林隙内生境对幼苗的遮蔽作用可能被土壤贫瘠和草根盘结等因素抵消[37]。樟子松在第3年时年高生长量高于20 cm,生长速度高于同一气候区的落叶松林皆伐迹地内的栽植苗,说明林冠下和林隙内生境促进了樟子松的生长[38]。第1年林冠下樟子松年生长量高于开敞度0.6~1.0的林隙内,到第2年和第3年林隙内年生长量高于林冠下,研究表明遮阴对幼苗第1年的生长有利,但林隙有利于幼苗一年后的生长[35]。
林隙开敞度和林隙内位置对光环境会产生显著影响[39]。从林隙中心到森林群落内部光、热、水等环境因素存在显著梯度变化,使得森林内部构成一定的生态分异[40]。吴祥云等(2008)研究表明开敞度0.5、1.0和1.5的林隙内南北轴上的光量子通量密度(PPFD)峰值均出现在中心位置[41],并且 开敞度为0.5的林隙内中心点正午的PPFD 为林冠下的10倍以上,为一些对光强要求适中的物种提供了生存条件[39]。
在开敞度为0.4~0.6的林隙内,栽植在林隙中央的油松和樟子松高度和基茎最大,因为小林隙内光照异质性低,中央受边界木的影响较小,中央可以接受更多地太阳辐射[17,19]。油松和樟子松的高度和基茎分别在开敞度0.6~0.8和0.6~1.0林隙内边缘高于中央,因为实验区较为干旱,较大林隙内中央位置水分不足。栽植后第1年,油松和樟子松的年高生长量分别在开敞度0.6~0.8和0.6~1.0林隙内南缘最大,因为南缘适中的光照强度造就的较低的土温和较高的土壤水分含量适合幼苗的生长[42,17]。但随着时间延长,林隙边缘对幼苗生长的促进作用减弱,与其他研究结果一致[13]。但在开敞度0.8~1.0林隙内油松的高度和基径在边缘小于中心,因为林隙内位置对林隙小气候的影响比较复杂,不能得出简单的相关性[43]。
实施封禁后天然更新密度显著提高,树种多样性增加,但密度仍然过低,人工栽植幼苗很有必要。与生长良好的人工针叶林相比,林隙面积和开敞度分布范围较大,存在面积和开敞度较大林隙。封禁凸显了林隙对天然更新的存进作用,开敞度0.5~0.6的林隙内天然更新密度最大,生长状况最好。在开敞度0.4~0.6林隙中央最适合油松生长,开敞度0.6~0.8林隙南北边缘最适合樟子松生长。本研究证实了在林隙内进行人工更新的可行性。未来可以通过伐除劣质的、生长不良的或病虫害严重的林木,人为制造林隙,并栽植阔叶树种提高树种多样性,在人工栽植前进行除草和松土;实现林冠梯级郁闭,林内大中小立木都能直接接受阳光,形成复层异龄混交林,增强森林生态系统的生态防护功能的经营目标。
[1] 胡明芳, 袁国胜, 甘代奎. 人促天然更新恢复南方地带性阔叶林植被的探讨[J].林业勘查设计, 2002, (1): 62-67.
[2] 新 刚, 郑焰锋, 赵浩彦, 等. 北京市生态公益林主要经营类型的经营措施研究[J]. 北京林业大学学报, 2011,33(4):13-21.
[3] 刘庆洪. 落叶松人工林中红松种群发生的初步研究[J]. 东北林业大学学报, 1986, 14(3): 27-33.
[4] 李贵祥, 孟广涛, 方向京, 等. 封禁措施对云南金沙江流域主要林分的生态功能影响[J]. 水土保持学报, 2007,21(1): 88-91.
[5] 于 飞, 王得祥, 史晓晓, 等.不同生态条件下松栎混交林3种优势乔木种群的更新规律[J]. 西北植物学报, 2013, 33(5):1020-1026.
[6] 谢锦升, 杨玉盛, 陈光水, 等. 封禁管理对严重退化群落养分循环与能量的影响[J]. 山地学报, 2002, 20(3): 325-330.
[7] 方代有, 朱东伟, 杨富权. 封禁措施对窿缘按水土保持林群落结构的影响[J]. 中国水土保持科学, 2005, 3(4): 119-123.
[8] 王 富, 贾志军, 董 智, 等. 不同生态修复措施下水库水源涵养区土壤粒径分布的分形特征[J]. 水土保持学报, 2009,23(5): 113-117.
[9] 张象君, 王庆成, 王石磊, 等.小兴安岭落叶松人工纯林近自然化改造对林下植物多样性的影响[J]. 林业科学, 2011,47(1): 6-14.
[10] Dupuy J M, Robin L C. Interacting effects of canopy gap,understory vegetation and leaf litter on tree seedling recruitment and composition in tropical secondary forests[J]. Forest Ecology and Management, 2008, 255(11): 3716-3725.
[11] Wang G, Fang L. The influence of gap creation on the regeneration of Pinus tabuliformis planted forest and its role in the near-natural cultivation strategy for planted forest management[J]. Forest Ecology and Management. 2011, 262(3):413-423.
[12] 蔡道雄, 贾宏炎, 卢立华, 等. 我国南亚热带珍优乡土阔叶树种大径材人工林的培育[J].林业科学研究,2007,20(2):165-169.
[13] Gagnon J L, Jokela E J, Moser W K,et al.Dlynamics of artificial regeneration in gaps within a longleaf pine flatwoods ecosystem[J]. Forest Ecology and Management, 2003, 172(2):133-144.
[14] Romell E, Göran H, Anders K,et al.Artif i cial canopy gaps in a Macaranga spp. dominated secondary tropical rain forest-effects on survival and above ground increment of four under-planted dipterocarp species[J]. Forest Ecology and Management, 2008,255(5): 1452-1460.
[15] Schulze M. Technical and financial analysis of enrichment planting in logging gaps as a potential component of forest management in the eastern Amazon[J]. Forest Ecology and Management, 2008, 255(3): 866-879.
[16] Lopes J C A, Jennings S B, Matni N M. Planting mahogany in canopy gaps created by commercial harvesting[J]. Forest Ecology and Management, 2008, 255(2): 300-307.
[17] Milakovsky B, Brent R F, Marks S A,et al.Influences of gap position, vegetation management and herbivore control on survival and growth of white spruce (Picea glauca (Moench)Voss) seedlings[J]. Forest Ecology and Management, 2011,261(3): 440-446.
[18] 曾思齐,黄炎俊,肖化顺, 等. 木荷次生林闽楠导入初期生长影响因素研究[J]. 中南林业科技大学学报, 2013, 33(8): 1-5.
[19] Coates K D. Conifer seedling response to northern temperate forest gaps[J]. Forest Ecology and Management, 2000, 127(1):249-269.
[20] York R A, Robert C H, John J B,et al.Group selection management in conifer forests: relationships between opening size and tree growth[J]. Canadian Journal of Forest Research,2004, 34(3): 630-641.
[21] Diaci J, Rok P, Andrej B. Regeneration in experimental gaps of subalpine Picea abies forest in the Slovenian Alps[J].European Journal of Forest Research, 2005, 124(1): 29-36.
[22] Nakamura A, Yukihiro M, Yasyko M. Adaptive management approach to increasing the diversity of a 30-year-old planted forest in an urban area of Japan[J]. Landscape and Urban Planning, 2005, 70(3): 291-300.
[23] 谭 笑, 孙向阳, 阎海平, 等. 北京西山地区人工针叶林林窗特征的研究[J]. 北京林业大学学报, 22(6): 64-66.
[24] 徐成立. 燕山山地华北落叶松人工林林隙特征及其影响[D].保定:河北农业大学, 2007.
[25] 王洪涛. 关帝山华北落叶松林隙更新调查研究[J]. 科技情报开发与经济, 2009, 19(24): 152-155.
[26] 王永安. 封山育林的生态经济作用[J]. 世界林业研究, 2000,13(3): 19-25.
[27] Sousa W P. Intertidal mosaics: propagule availability and spatially variable patterns of succession[J]. Ecology, 1984, 65(6): 1918- 1935.
[28] Bolton N W, Anthony W D. 2011. Regeneration responses to gap size and coarse woody debris within natural disturbance-based silvicultural systems in northeastern Minnesota, USA[J]. Forest Ecology and Management. 262(7): 1215-1222.
[29] Dale M R T, Powell R D. A new method for characterizing point patterns in plant ecology[J]. Journal of Vegetation Science, 2001,12(5): 597-608.
[30] Coates K D. Tree recruitment in gaps of various size, clearcuts and undisturbed mixed forest of interior British Columbia, Canada[J].Forest Ecology and Management, 2002, 155(1): 387-398.
[31] Wolf A. Fifty year record of change in tree spatial patterns within a mixed deciduous forest [J]. Forest Ecology and Management,2005, 215(1): 212-213.
[32] Arévalo J R, José M F. Treefall gaps and regeneration composition in the laurel forest of Anaga (Tenerife): a matter of size[J]. Plant Ecology, 2007, 188(2): 133-143.
[33] Popma J, Bongers F, Martinez-Ramos M. Pioneer species distribution in treefall gaps in neotropical rain forest: a gap def i nition and its consequences[J]. Journal of Tropical Ecology,1988, (4): 77-88.
[34] Poorter L, Yáskara H. Effects of seasonal drought on gap and understory seedlings in a Bolivian moist forest[J]. Journal of Tropical Ecology, 2000, 16(4):481-498.
[35] Collins B S, Battaglia L L. Microenvironmental heterogeneity and Quercus michauxii regeneration in experimental gaps[J].Forest Ecology and Management, 2002, 155(1): 279-290.
[36] 朱小龙, 赖志华, 黄承勇, 等. 长苞铁杉幼苗在林窗不同位置的建立[J]. 广西植物, 2008, 28 (4): 473 - 477.
[37] 薛 曦, 刘 野, 架景仁. 华北石质低山阳坡油松植苗造林技术研究[J]. 林业科技通讯, 1992, (2): 10-13.
[38] 路治林. 落叶松采伐迹地更新树种生长对比试验初报[J]. 辽宁林业科技, 1992, (3): 16-18.
[39] 陈 梅, 朱教君, 闫巧玲, 等. 辽东山区次生林不同大小林窗光照特征比较[J]. 应用生态学报, 2008, 19(12): 2555 - 2560.
[40] 洪 伟, 吴承桢, 林成来, 等. 福建龙栖山森林群落林窗边缘效应研究[J]. 林业科学, 2000, 36(2): 33-38.
[41] 吴祥云, 陈 梅, 屈雯雯, 等. 辽东地区次生林林窗生长季初期的光环境特征[J]. 辽宁工程技术大学学报, 2010, 29(4):665-669。
[42] Raymond P, Alison D M, Jean-Claude R,et al.Spatial patterns of soil microclimate, light, regeneration, and growth within silvicultural gaps of mixed tolerant hardwood-white pine stands[J].Canadian journal of forest research, 2006,36(3):639-651.
[43] 冯 静, 段文标, 陈立新. 阔叶红松混交林林隙大小和林隙内位置对小气候的影响[J]. 应用生态学报,2012,23(7):1758-1766.
[44] 王卫军, 赵婵璞, 任 仙, 等. 华北落叶松人工林林分密度对土壤持水能力的影响[J]. 中南林业科技大学学报, 2013,33(10): 95-98.
Recruitment characteristics of gaps in degraded Larix pricipis-rupprechtii plantation
DONG Bo-qian1, HUANG Xuan-rui2, XU Xue-hua2, ZHANG Yu-zhen2
(1.Key Lab. for Silviculture and Conservation of State Education Ministry, College of Forestry, Beijing Forestry University, Beijing,100083, China; 2. College of Forestry, Agriculture University of Hebei, Baoding 071000, Hebei, China)
Based on the investigation on 52 gaps in degradedLarix pricipis-rupprechtiiplantation in mountainous region of northern Hebei province, the gap characteristics of forest and effects of opening degree on natural recruitment after closing measures were studied. The results show that in studied area, the gaps widely distributed and most gaps had smaller opening degree; three years after the closed forest project, the recruit density and species diversity increased obviously, but the density was still low and the recruitment community was mainly composed ofL. pricipis-rupprechtiiandPopulus davidiana; in general, after closed the recruit density in the gaps were higher than that under the canopy; in the gaps of 0.5~0.6 opening degree the recruit density were the highest,L. pricipisrupprechtiiandPopulus davidianagrew best in gaps of 0.4~0.5 and 0.7~0.8 opening degree respectively; Artificial plantedPinus tabulaeformisandPinus sylvestrisvar.mongolicagrew best at the gaps center of 0.4~0.6 opening degree and gaps edge of 0.6~0.8 opening degree respectively; With the extension of closed forest time, the effects of gaps and position in gaps on artif i cial regeneration seedlings’ growth became more obviously. The closed forest project promoted the natural recruitment, but the effect was limited, so it is necessary to recruit by human.
Larix pricipis-rupprechtiiplantation degraded; close-nature management measure; closed forest project; gap characteristics;effect of artif i cial regeneration
S791.22
A
1673-923X(2014)08-0001-08
2014-03-05
国家公益专项“燕山山地典型森林类型健康经营技术”(20100400205);科技部十二五科技支撑项目“华北土石山区典型森林类型可持续经营技术研究”(2012BAD22B03)
董伯骞(1984-),男,博士,主要研究方向:森林经营和退化生态系统恢复
黄选瑞(1962-),男,博士后,教授,主要研究方向:林业可持续发展、森林资源管理;E-mail:hxr1962@163.com
[本文编校:吴 彬]
