尾叶桉GLU4肉桂酰-辅酶A还原酶基因克隆及原核表达
2014-01-02陈博雯蒋湘宁
陈博雯 ,盖 颖 ,蒋湘宁
(1.北京林业大学 生物科学与技术学院,北京 100083;2. 广西林科院 广西优良用材林资源培育重点实验室,广西 南宁 530002)
尾叶桉GLU4肉桂酰-辅酶A还原酶基因克隆及原核表达
陈博雯1,2,盖 颖1,蒋湘宁1
(1.北京林业大学 生物科学与技术学院,北京 100083;2. 广西林科院 广西优良用材林资源培育重点实验室,广西 南宁 530002)
肉桂酰-辅酶A还原酶(Cinnamoyl Co-A Reductase,CCR)是木质素合成中的关键酶,根据植物中CCR保守序列设计引物,以尾叶桉GLU4嫩茎为材料克隆到其CCR基因,命名为EuCCR。该基因gDNA长2 918 bp,cDNA长1 045 bp,CDS区编码336个氨基酸。EuCCR核酸序列与GenBank已登录的桉属植物CCR基因同源性达到96%以上,与伞房属、杯果木属植物CCR的同源性达85%以上,其编码的氨基酸序列经比对发现具有完整FR_SDR_e结构域及NADP结合位点和底物结合位点,与可可树等植物中CCR基因编码序列同源性也在84%以上,确定为CCR基因。对EuCCR蛋白序列理化性质及结构进行生物信息学分析,利用MEGA软件对基因序列进行系统进化树分析。采用pQE30/M15系统对EuCCR进行原核表达,重组质粒成功表达分子量约36 kD的目的蛋白。本研究从尾叶桉GLU4中克隆得到EuCCR基因并原核表达,为该基因的酶学分析以及利用该基因转化调控尾叶桉木质素合成奠定基础。
尾叶桉;肉桂酰-辅酶A还原酶;生物信息学分析;原核表达
桉树是桃金娘科桉属植物的总称,原产地为澳大利亚,桉树品种多、生长快,其木材工业特性好,坚韧耐腐,是最常用的制造纸浆的主要原料[1-2]。在我国热带、亚热带地区多个省份都种植有桉树人工林[3-8]。广西的桉树研究在我国处于领先地位。20世纪70年代初期就开始引种,经过多年的引种试验及杂交选育,得到了多个优良树种、优良种源及优良家系[2],截止2010年,全区桉树面积达2 480万亩,占全区人工商品林面积的30.5%,居全国第1位,桉树已经成为短周期原料林的主要造林树种,在造纸木浆生产中有着极重要的地位。
木质素是植物细胞壁中的一类苯丙烷类衍生物,去除木质素是制浆工艺中的重要步骤,直接关系到整个制浆生产的污染情况和成本高低。利用转基因技术转化木质素合成途径中的关键酶,从而降低植物木质素含量,从根本上减少制浆中都污染成本,是目前的研究热点。肉桂酰-辅酶A还原酶(Cinnamoyl Co-A Reductase,CCR)催化木质素单体的生物合成中第一个特定步骤,它将相应的化合物的CoA 形式转化为醛的形式。是几种木质素单体合成途径中的共有酶,对木质素的合成起重要作用[9],是调控木质素合成的首选基因。
尾叶桉GLU4无性系(Eucalyptus urophyllaclone GLU4)是广西审定的桉树良种,材性适宜造纸,在广西种植范围广,以此树种作为转基因育种材料,具有良好的应用前景。本研究以尾叶桉GLU4无性系为材料,克隆得到EuCCR基因,并对其序列进行生物学分析, 而后对EuCCR基因进行原核表达分析,为该基因的酶学分析以及更进一步利用该基因转化调控尾叶桉木质素合成奠定基础。
1 材料与方法
1.1 实验材料
供试材料为尾叶桉GLU4无性系Eucalyptus urophyllaclone GLU4,取自广西林科院生物工程与技术研究所苗圃。
DNA secure新型植物基因组DNA提取试剂盒、RNA prep Pure多糖多酚植物总RNA提取试剂盒、质粒提取试剂盒及DNA纯化、凝胶回收试剂盒购自北京天根生化科技有限公司。RNA LA PCR反转录试剂盒、LA Taq DNA 聚合酶、克隆载体pMD-18T购自TaKaRa公司,限制性内切酶购自 Promega公司,IPTG、X-Gal、Ampicillin、kanamycin购自北京索莱宝科技有限公司。E.coliDH5α、M15菌株及表达载体PQE30为本实验室保存。基因扩增引物由北京华大基因生物技术有限公司合成。
1.2 EuCCR序列PCR扩增
取移栽培养3~4周的GLU4组培苗茎部组织,分别按照试剂盒说明书方法提取基因组DNA和总RNA,并取总RNA按照反转录试剂盒方法合成cDNA,于-80 ℃保存备用。
在NCBI检索其他植物中已报道的CCR基因序列,blast分析保守区用Vector NTI软件设计引物:
Sense primer CCR-F1: CAATCCCACCATGCC CGTCG
Anti-sense primer CCR-R1: GTGCGCGCCATG ATGGATCTAAGATC
分别以gDNA和cDNA作为模板,扩增目的基因的gDNA片段及cDNA片段。PCR反应扩增程序:95 ℃预变性5 min;95 ℃变性30 s,60 ℃退火30 s,72 ℃延伸3 min(cDNA扩增延伸时间1 min),28个循环;72 ℃继续延伸10 min,4 ℃保存30 min。
1.3 克隆载体构建
gDNA及cDNA PCR扩增产物分别经0.8%琼脂糖凝胶电泳检测,切下目的条带凝胶回收后与pMD-18T载体16℃连接过夜,转化E.coliDH5α,随机挑取3个阳性克隆用CCR-F1/CCR-R1引物PCR验证后,由华大基因生物技术有限公司完成测序。
1.4 序列生物信息学分析
通过NCBI搜索引擎中的Blastn在线工具进行同源性分析;利用NCBI的ORF Finder寻找阅读框,完成DNA序列的翻译,并用Blastp分析蛋白序列同源性及保守区域、活性位点;利用Vector NTI中的AliginX软件将gDNA和cDNA序列测序结果比对分析外显子和内含子;运用Expasy网站的ProtParam完成蛋白序列的氨基酸组成、等电点及亲疏水性分析;应用丹麦科技大学(DTU)提供的TMHMM和SignalP工具分析序列中的跨膜区和信号肽;利用ExPaSy中的psipred分析序列二级结构;应用SWISS-MODEL软件对基因氨基酸序列进行同源三级结构建模并分析;应用MEGA 5.0工具的ClustalW分析EuCCR与其他物种中CCR基因序列,并采用Neighbor-Jioning算法构建进化树
1.5 表达载体构建
根据克隆到pMD-18T载体上的CCR cDNA序列,采用Vector NTI软件设计引物(带有酶切位点)
Sense primer CCR-F2(BamHⅠ): GGGGGA TCCATGCCCGTCGACGCCCTC
Anti-sense primer CCR-R2(HindⅢ):GGGAAGCTTTCATCCCTGAATACGCAC
PCR扩增CCR cDNA的CDS序列,扩增产物回收后BamHⅠ/HindⅢ双酶切,同时将PQE30质粒BamHⅠ/HindⅢ双酶切。 37℃酶切4 h后0.8%琼脂糖凝胶电泳检测、回收目的基因片段及线性PQE30质粒,用T4 DNA连接酶16℃连接过夜,转化E.coliDH5α。挑选阳性克隆用CCR-F2/CCR-R2引物PCR验证,并用BamHⅠ/HindⅢ双酶切验证。
1.6 原核表达及SDS-PAGE检测
将验证后的重组质粒转化至E.coliM15,Ampicillin、kanamycin双抗性筛选阳性克隆,接种于 10mL 含Ampicillin、kanamycin的 LB 液体培养基,37℃200 r/min过夜培养,按1%接种量转接至100 mL LB培养基中(含Ampicillin,kanamycin),振荡培养 3~5 h 至 OD600为0.4~0.6,加入IPTG 至终浓度为 1 mmol/L,同时设立不加入IPTG的处理作为对照。继续振荡培养 4 h,离心收集菌体,用 1 mL PBS 缓冲溶液悬浮后,超声波破胞, 离心取8 μL上清液加入2 μL 5×上样缓冲溶液, 混匀后沸水浴 5 min,离心取5 μL上清SDS-PAGE检测。 蛋白条带用考马斯亮兰R250染色。
2 结果与分析
2.1 GLU4中CCR基因克隆
通过PCR成功的扩增获得长度分别为3 000 bp、1 000 bp左右的目的片段(见图1a),与推测的CCR基因的gDNA片段以及cDNA片段长度相符,条带清晰无非特异性杂带。目的片段连接至pMD-18T载体后挑选阳性克隆用CCR-F1/CCR-R1引物进行PCR 验证,分别得到长度为3 000 bp和1 000 bp左右的片段(见图1b),说明目的片段已经成功克隆到pMD-18T载体上,重组质粒分别命名为pT-CCR-gDNA和pT-CCR-cDNA,各取3个克隆测序。测序结果显示克隆得到的CCR gDNA序列长度2 918 bp,cDNA序列长度1 045 bp。利用NCBI在线blast分析,序列与冈尼桉(X79566.1),蓝桉(AB591264.1),柳叶桉(AF297877.1),圆果桉(FJ834288.1)中CCR基因一致性分别为98%、99%、98%、96%,与GenBank已登录的伞房属、杯果木属植物的CCR基因一致性也达到90%(DQ309047.2),89%(EF612728.1),85%(DQ309061.1),且序列中包含有完整CDS,初步说明克隆所得片段为的CCR基因,命名为EuCCR。

图1 EuCCR基因的克隆Fig.1 Cloning of EuCCR gene
利用NCBI的ORF Finder将EuCCR基因编码区翻译氨基酸序列,编码336个氨基酸,应用NCBI的blastp比对分析氨基酸序列,结果显示序列中包含完整的FR_SDR_e保守结构域以及NADP结合位点和底物结合位点(见图2b),FR_SDR_e属于是NADP依赖还原酶家族蛋白特有的保守结构域,在植物黄酮类等次级代谢途径中的异构酶、还原酶和裂解酶中都含有此结构域,与CCR酶学功能相符。氨基酸序列比对表明,EuCCR编码序列与蓝桉(AAT74879.1)、柳叶桉(AAG16242.1)、冈尼桉(CAA56103.1)CCR基因编码序列一致性为99%,与异心叶桉(AAT74875.1)、圆果桉(ACZ59063.1)、可可树(EOY26038.1)、橡胶树(ADU64758.1)、碧桃(EMJ16703.1)、毛白杨(ACE95172.1)、毛果杨(CAC07424.1)中CCR基因编码序列一致性分别为98%、96%、85%、85%、84%、84%,进一步证实克隆到的基因序列确实为CCR基因。

图2 EuCCR序列分析Fig.2 Analysis of EuCCR sequence
使用Vector NTI中的AliginX软件将EuCCR的gDNA和cDNA序列比对,显示基因含有5个外显子,4个内含子(见图2a),其中第4个外显子和第4个内含子为基因中最长的外显子和内含子,这与Poke文献报道一致[10]。
2.2 EuCCR序列分析
2.2.1 理化性质及结构预测
利用ExPaSy中的Protparam工具分析EuCCR编码蛋白序列,结果表明该蛋白由336个氨基酸残基组成,分子式为C1624H2592N434O489S13;相对分子量为36.44 kD;理论pI值为5.87,为酸性蛋白;总平均亲水性为-0.078,不稳定系数为30.95,小于40,推断蛋白质为稳定性蛋白;从氨基酸组成分析,带负电荷的氨基酸残基(Asp+Glu)共41个,带正电荷的氨基酸残基(Arg+ Lys)共36个;综合分析表明该蛋白质是一个带负电荷的酸性蛋白质。
应用丹麦科技大学(DTU)提供的TMHMM在线分析跨膜区,未发现跨膜结构,SignalP在线分析结果显示,EuCCR编码蛋白不包含信号肽序列,说明该蛋白不属于膜蛋白或分泌蛋白,应该是定位于细胞质基质中,这与该基因的功能定位相符。
利用ExPaSy中的psipred分析序列二级结构,显示该蛋白具有11个α-螺旋和11个β-折叠结构(见图3a),运用SWISS-MODEL对EuCCR进行同源三级结构建模。构建范围从第10个氨基酸到第326个氨基酸(见图3b),其建模参考模板为葡萄的二氢黄酮醇-4-还原酶蛋白,该蛋白也属于NADP依赖还原酶家族[11]。目标蛋白与参考蛋白的序列比对同源性为76.28%;模型评估E值为0.00e-1,说明此次同源建模可靠性极高,推测EuCCR具有NADP依赖的还原酶功能,这与CCR的酶学功能符合。
2.2.2 系统进化树分析
在NCBI蛋白质数据库检索已登录的CCR蛋白质序列,共检索到920条植物CCR蛋白序列,选择各科属植物中的26条序列与EuCCR进行序列比对分析。应用MEGA 5.0工具的ClustalW分析EuCCR与其他物种CCR编码蛋白序列同源性,并采用Neighbor-Jioning算法构建进化树(见图4)。
系统进化分析显示,EuCCR与蓝桉、圆果桉、柳叶桉等桉属植物中CCR聚为一类,之后与杯果木属、伞房属植物CCR聚为一类,进化分析结果与分类学相一致。EuCCR与杨属、松属CCR亲缘性较远,但同源性也达到83%和78%,说明CCR在各物种的进化中仍是较为保守的基因,这可能与CCR基因在植物生长中的组成型功能有关。
2.3 EuCCR原核表达
采 用 引 物 CCR-F2/CCR-R2, 以pT-CCR-cDNA作为模板,PCR扩增EuCCR的完整CDS序列,双酶切连接pQE30构建重组表达载体,采用CCR-F2/CCR-R2引物进行PCR验证,电泳结果如图5a,3个阳性克隆均扩增得到1 045 bp长度的片段。将PCR验证的3个阳性克隆提取质粒进行酶切验证,电泳结果如图5b, 3个阳性克隆均酶切得到了3 600 bp左右的载体片段和1 045 bp的插入外源片段,说明EuCCR原核表达重组载体构建成功,命名为pQE30-EuCCR。

图3 EuCCR蛋白质结构分析Fig.3 Protein structure analysis of EuCCR

图4 EuCCR系统树进化分析Fig. 4 Phylogenetic analysis of EuCCR

图5 pQE30-EuCCR构建Fig.5 Vector construction of pQE30-EuCCR
将pQE30-EuCCR转入表达宿主M15中IPTG诱导原核表达,SDS-PAGE结果如图6。根据EuCCR氨基酸序列预测蛋白质分子量为36.44 kD,诱导表达后,阳性转化株中出现大小约36 kD 的蛋白条带,与预期相吻合,未加IPTG的对照则没有相应的条带,说明重组质粒经 1.0 mmol/L IPTG 诱导 3 h 后成功表达目的蛋白。
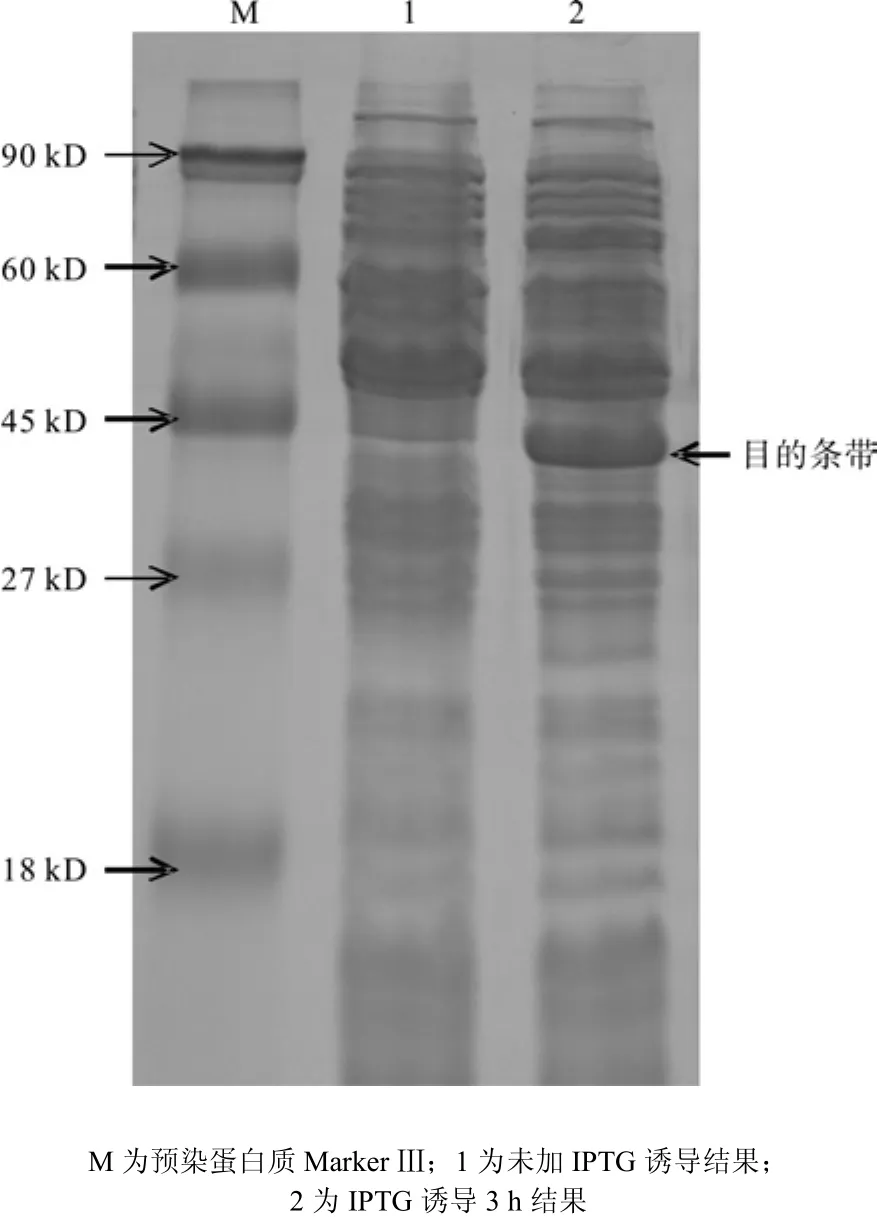
图6 EuCCR基因原核表达SDS-PAGE结果Fig.6 Prokaryotic expression SDS-PAGE of EuCCR
3 结论与讨论
CCR是木质素合成途径中的关键酶,被认为是潜在的碳向木质素分配的控制关节点,是调控植物木质素合成的理想靶标[12]。通过转基因技术负调节烟草中CCR,发现转基因植株中木质素含量明显降低同时一些非正常的酚类物质增加[13-14]。部分反义转化CCR的转基因植株中,S 与G 木质素含量均降低的同时,伴随生长停滞,叶型卷缩,花期延长,导管变形,木质部有颜色变化等异常表型,但将CCR 和CAD 负调节的单转基因植物杂交,可以得到木质素含量降低且表型正常的后代,说明两个基因可能协同作用降低木质素含量[15],也说明木质素含量明显降低的植物至少在自然条件下可以进行正常的发育。
Poke等[10]对23个桉树树种的CCR基因遗传进化关系进行研究,测序得到大约44%的CCR基因,序列只限于内含子 4和部分外显子 4和5,并未得到CCR的完整CDS。根据Poke等提交的序列分析,其部分外显子序列与本研究中得到EuCCR具有很高的一致性,更进一步确定了EuCCR的可靠性。
本研究利用RT-PCR技术从尾叶桉GLU4幼苗茎部组织中克隆到肉桂酰辅酶A还原酶基因,利用多种生物信息学工具通过同源比对、保守序列及功能位点分析、同源建模、系统进化树构建等方法对EuCCR进行序列分析,确定其为尾叶桉GLU4无性系中的CCR基因,为下一步构建反义或RNAI载体转化植株奠定基础。在本研究中,通过亚克隆构建原核表达载体对EuCCR进行原核表达,得到分子量与预期一致的CCR蛋白,也为下一步EuCCR的酶学活性分析提供了材料。
[1] 莫晓勇,龙 腾,彭仕尧,等.桉树嫩枝扦插育苗技术研究[J].中南林学院学报,2002,22(4):31-35.
[2] 项东云,陈健波,叶 露,等.广西桉树人工林发展现状、问题与对策[J].广西林业科学2006,35(4):195-201.
[3] 汤珧华,李志辉.耐寒性桉树早期选择[J].中南林学院学报,2002, 20(3):70-74.
[4] 陈茜文,杨模华,李志辉.耐寒桉树的纤维形态特征的变异[J].中南林学院学报,2002,22(4):61-65.
[5] 刘友全,刘加林,潘天玲.赤桉在湖南的抗寒与生长适应性[J].中南林学院学报,2002,22(3):86-89.
[6] 胡曰利,吴晓芙,王尚明,等.桉树人工林地有机物和养分库的衰退及防治[J].中南林学院学报,2002,22(4):49-53.
[7] 邓运光,李群伟,张伟佳.桉树部分种类种源试验研究初报[J].中南林学院学报,2002,22(3):49-53.
[8] 谢耀坚.中国桉树人工林可持续发展经营战略初探[J].世界林业研究,2003,16(5): 59-64.
[9] Lacombe E, Hawkins K, Doorsselaere J V,et al. Cinnamyol CoA reductase, the fi rst committed enzyme of the lignin branch biosynthetic pathway: cloning,expression and phylogenetic relationships[J]. Plant Journal,1997(11): 429-441.
[10] Poke FS, Martin DP, Steane DA,et al.The impact of intragenic recombination on phylogenetic reconstruction at the sectional level in Eucalyptus when using a single copy nucleargene(cinnamoyl CoAreductase) [J]. Molecular Phylogenetics and Evolution, 2006(39):160-170.
[11] Petit P, Granier T, d’Estaintot BL,et al. Crystal structure of grape dihydroflavonol 4-reductase, a key enzyme in flavonoid biosynthesis[J]. Journal of Mol. Biol. ,2007 ,368(5):1345-57
[12] 赵华燕,魏建华,宋艳茹.木质素生物合成及其基因工程研究进展[J].植物生理与分子生物学学报,2004,30(4):361-370
[13] Ralph J, Hatf i eld RD, Piquemal J,et al.NMR characterization of altered lignins extracted from tobacco plants down-regulated for lignification enzymes cinnamylalcohol dehydrogenase and cinnamoyl-CoA reductase[J]. Proc. Natl. Acad. Sci. USA,1998,95:12803-12808.
[14] Chabannes M, Barakate A, Lapierre C,et al.Strong decrease in lignin content without signif i cant alteration of plant development is induced by simultaneous down-regulation of cinnamoyl CoA reductase (CCR) and cinnamyl alcohol dehydrogenase (CAD) in tobacco plants[J].Plant J, 2001,28: 257-270.
[15] Chabannes M, K Ruel. In situ analysis of lignins in transgenic tobacco reveals a differential impact of individual transformations on the spatial patterns of lignin deposition at the cellular and subcellular levels[J]. Plant Journal, 2001,28(3):71-82.
Cloning and prokaryotic expression of cinnamoyl Co-A reductase ofEucalyptus urophyllaclone GLU4
CHEN Bo-wen1,2, GAI Ying1, JIANG Xiang-ning1
(1. School of Biology Science and Biotechnology, Beijing Forestry University, Beijing 100083, China; 2. Guangxi Key Lab. of Superior Timber Trees Resource Cultivation, Guangxi Forestry Research Institute, Nanning 530002, Guangxi, China)
Cinnamoyl Co-A reductase is a key enzyme in lignin synthesis pathway. A CCR gene was cloned from the immature stem ofEucalyptus urophyllaclone GLU4 and namedEuCCRby using specif i c primers based on the highly conserved sequences of plant CCR. The gDNA and cDNA sequence ofEuCCRwere 2918 bp and 1045 bp respectively, which CDS encodes 336 amino acid residues.EuCCRhad more than 96% sequence homology withEucalyptusthat had been logged in GenBank, and its homology withAngophoraandCorymbiawere over 85%.EuCCRencoding sequence was analyzed, the result shows it has an entire FR_SDR_e domain, a NADP binding site and a substrate binding site. The homology with Theobroma cacao and others were over 84%. The physicochemical property, structure of EuCCR and its phylogenetic analysis were analyzed by using bioinformatics tools and MEGA. The SDS-PAGE analysis showed thatEuCCRwas transformed into pQE30/M15 system and fusion protein with molecular weighting about 39 kD was successfully expressed in transformant. The cloning and expression ofEuCCRgene provided some effective resources for enzymology research and further transgenic research.
Eucalyptus urophylla; Cinnamoyl Co-A reductase; bioinformatics analysis; prokaryotic expression
S792.39
A
1673-923X(2014)11-0071-06
2014-01-12
国家自然科学基金项目(31400522);广西林业科技项目(桂林科字[2014]第33号);广西优良用材林资源培育重点实验室开放课题基金项目(12A0302)
陈博雯(1983-),女,博士研究生,主要从事植物生物技术方面的研究;E-mail:grf i_bwchen@163.com
蒋湘宁(1958-),男,教授,主要从事植物分子生物学方面的研究;E-mail:xiangning_jiang@163.com
[本文编校:吴 毅]
