欧洲鹅耳枥2年生播种苗年生长动态
2014-01-02程龙霞金纯子祝遵凌
程龙霞 ,金纯子 ,祝遵凌 ,b
(南京林业大学 a.风景园林学院; b.艺术设计学院,江苏 南京 210037)
欧洲鹅耳枥2年生播种苗年生长动态
程龙霞a,金纯子a,祝遵凌a,b
(南京林业大学 a.风景园林学院; b.艺术设计学院,江苏 南京 210037)
以欧洲鹅耳枥2年生播种苗为研究对象,对其苗高、地径、生物量的年动态变化和苗高与地径、生态因子的关系等进行研究。结果表明:苗高、地径两者生长基本符合“慢—快—慢”的生长规律,可用Logistic方程来拟合,同时把苗高和地径的生长可分为生长初期、生长盛期和生长末期;苗高在生长盛期有一连续生长高峰期,而地径在生长盛期有2次高峰期,且后期比前期更明显;苗高与地径变化之间呈线性相关,播种苗各部分生长具密切相关性,呈负相关性;通过苗高、地径生长规律与气象因子的相关性分析,表明旬平均气温是影响苗高的主导因子,其他气象因子对地径生长无显著影响。
欧洲鹅耳枥;生长动态; 苗高; 地径; 生物量
欧洲鹅耳枥Carpinus betulus,又名西洋千金榆和“欧洲角木”,隶属桦木科Betulaceae鹅耳枥属Carpinus植物。欧洲鹅耳枥约5 000年前就广泛分布在欧洲中部,是原始森林群落重要的组成部分之一[1-2]。欧洲鹅耳枥因其枝型紧凑、树冠丰满、季节色相变化明显等特征,深受人们喜爱,在欧洲已有1 800多年的栽培历史[3]。作为著名的彩色叶树种,欧洲鹅耳枥在欧美等国家园林绿化上多作行道树、绿篱等得到广泛应用。对欧洲鹅耳枥的研究主要集中在育种、生理生化和地理分布等方面,并取得一定成果[4]。随着我国园林事业的发展,对彩叶树种的种类要求越来越多。上海、芜湖等地亦有少量引种栽培,但以国外购进大规格苗木为主,缺乏引种驯化及适应性的系统性研究。本研究以欧洲鹅耳枥2年生播种苗为材料,对其年生长规律进行了探讨,研究苗木生长特性,以期对我国欧洲鹅耳枥的引种、驯化工作提供一定的理论和技术支持。
1 材料与方法
1.1 试验地概况
试验地设在南京林业大学实验教学中心。地理位置为北纬 32°04′34″、东经 118°48′42″,属亚热带季风气候,年平均气温15.7 ℃,年极端最高温43 ℃,最低温-14 ℃,四季分明,雨量充沛,年均降水量1 021.3 mm,无霜期较长。
1.2 试验材料与方法
试验的种子于2010年采自匈牙利,经6~7个月变温层积后,于2011年3月播种。2012年4月至10月,选50株生长良好的小苗作为标准株,每15 d测定1次苗高、地径;苗高生长量用钢尺测量,地径生长量用游标卡尺测量。每月中旬选3株与标准株长势一致的植株,测其根长及生物量,分别称取根、茎、叶鲜质量,经105℃烘干至恒质量后,进行生物量的测定,重复3次。栽培期间进行常规管理。
1.3 数据处理及模型建立
用SPSS 17.0、Excel 2010软件等对数据进行统计分析及作图。利用Logistic方程对2年生欧洲鹅耳枥苗高、地径的生长规律进行拟合。K、a、b的初始值用三点法估计[5]。
2 结果与分析
2.1 苗高年生长规律
对2年生欧洲鹅耳枥幼苗的全年苗高生长定期观测,结果如表1所示。欧洲鹅耳枥幼苗的初期生长迟缓,6月中下旬以后,苗高生长加快,7月中下旬出现生长高峰,此后一直保持较高的生长速度,到9月中旬增速减缓,呈现明显的“慢—快—慢”的节律,这与杨树幼苗生长规律基本一致[6],到10月中旬苗高生长基本停止。
苗高Logistic方程为:y=24.427/(1+e2.044-0.228t),曲线拟合度为0.951,拟合性较好(见图1)。方程的回归性检验达到显著水平(p<0.01),说明模拟的欧洲鹅耳枥苗高Logistic曲线与实测曲线间的符合程度比较高,用回归值来推测实际值有较高准确性[7]。
对方程一阶求导可得连日苗高生长量随时间变化的方程,二阶求导得到苗高连日生长量变化速率曲线[8]。令二阶导数等于零,t=8.96,即生长第119天(8月12日)达速生点。令方程三阶导数等于零,得到t1=3.189、t2=14.74,即苗木生长的第33天(5月18日)、第206天(11月8日)分别为生长初期到生长盛期及由生长盛期转入生长后期的分界点。由此可将2年生欧洲鹅耳枥幼苗苗高生长划分为3个时期:生长初期(5月18日之前)、生长盛期(5月18日~8月12日)和生长末期(8月13日~ 11月8日)。与实际观测结果较为接近,说明方程拟合效果明显。

表1 欧洲鹅耳枥苗高定期生长观测结果Table 1 Regular observation results of height growth of Carpinus betulus
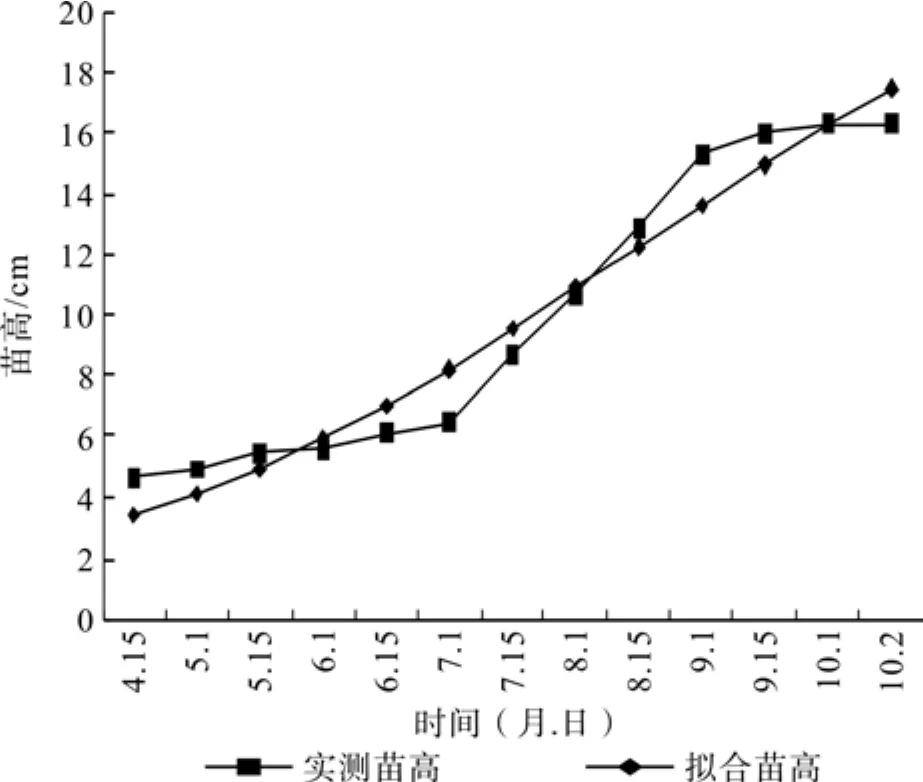
图1 欧洲鹅耳枥苗高累积生长量曲线与Logistic拟合曲线Fig.1 Height growth accumulations and simulated Logistic curves of Carpinus betulus
2.2 地径生长规律
对欧洲鹅耳枥地径生长的全年观测结果如表2所示。欧洲鹅耳枥幼苗地径生长缓慢,6月初和8月中旬是生长的2个高峰期,后一次高峰较前一次更为明显,10月中旬地径的生长基本停止。这与苗高生长的停止时间基本一致。

表2 地径定期生长观测结果Table 2 Regular observation results of ground diameter of Carpinus betulus
拟合的欧洲鹅耳枥地径Logistic方程为:y=0.532/(1+e1.399-0.176t)。曲线拟合度为0.987,方程的回归性达到极显著水平(p<0.01),说明模拟地径的Logistic曲线与实测曲线间的符合程度比较高(见图2)。同时由图2的拟合曲线也可以看出,地径的拟合曲线效果要比苗高的拟合效果好。

图2 地径累积生长量曲线与Logistic拟合曲线Fig.2 Simulated Logistic curves of ground diameter growth accumulations of Carpinus betulus
令二阶导数等于零,t=7.95,即生长期第104天(7月27日)地径生长速度最快;对方程三阶求导,t1=0.47(4月8日)、t2=15.43(10月16日),分别是生长初期到生长盛期及生长盛期转入生长后期的分界点。由分析可将幼苗的地径生长划分为3个时期:生长初期(4月8日之前)、生长盛期(4月8日~7月27日)和生长末期(7月28日~10月16日)。与实际观测结果较为接近,可信度较高。
2.3 苗高、地径的生长关系
一般来说,树木在不同生长时间苗高的生长随着地径的生长而变化,并且变化趋势基本一致[9]。通过回归分析得到欧洲鹅耳枥苗高、地径的线性回归方程为:Y=54.745X-3.4983 (R2=0.9587,p<0.01)。式中:Y为苗高的生长量;X为对应时间的地径生长量。表明苗高、地径生长呈现显著的线性关系。
比较实测苗高、地径的生长关系曲线和拟合苗高、地径的生长关系曲线(见图3),实测点基本都在拟合直线周围波动,说明模拟曲线与实测曲线间吻合度较高,以回归值来推测实际值准确性较高。

图3 欧洲鹅耳枥苗高、地径关系变化与Logistic拟合变化Fig.3 Changes of height and diameter relation and simulated Logistic curves of Carpinus betulus
由前面分析可知,苗高、地径的实际测量与理论的分析结果都有部分波动,很大一部分原因有可能与外界环境条件有关 ,所以有必要分析苗高、地径的生长与气象因子的相关性。
2.4 欧洲鹅耳枥苗高、地径年生长规律与气象因子的关系
苗木的生长受周围环境因子和植物自身生理的综合作用。不同因子对苗木生长的影响也是不一样的[10-11]。欧洲鹅耳枥幼苗的苗高、地径与气象因子的调查数据见表3。
为了了解气象因子与鹅耳枥生长的相关性,筛选其主导因子,建立线性回归模型来探讨它们之间的关系[12]。对数据进行多元线性回归分析,得到苗高与各气象因子的关系方程为:

式中:X1为旬平均气压;X2为旬平均气温;X3为旬一小时降水;H为苗高的旬生长量。
对所建立的模型进行相关性分析,得到模型的复相关系数R为0.928, 显著性明显(P<0.05),方程的拟合效果好。对各个气象因子的显著性检验,得到旬平均气温显著性明显(P<0.05),其他气象因子影响不显著性,湿度对苗高基本没有影响。得出旬平均气温是影响欧洲鹅耳枥苗高生长的主导气象因子。

表3 欧洲鹅耳枥幼苗的苗高、地径与气象数据Table 3 Height, diameter and meteorological data of Carpinus betulus
同样利用分析得出地径与各气象因子的关系方程,经t检验复相关系数没达到显著水平,偏相关系数也没达到显著水平,推知欧洲鹅耳枥播种苗地径的生长与气象因子无显著相关性和规律性。
2.5 欧洲鹅耳枥根长与苗高、地径的关系
苗高、地径的生长与苗木根系的生长以及健康状况是有一定相关性的[13]。如图4所示,主根的生长与苗高和地径生长趋势基本一致,具有一定的相关性。

图4 主根长与苗高、地径的年动态变化Fig.4 Annual changes of main root length, seedling height and ground diameter of Carpinus betulus
通过回归分析,主根(L)和苗高(H)、地径(D)生长之间的回归方程分别为:

由回归方程表明,苗木各个部分的生长有着密切的相关性,且相关系数都达到了显著水平,方程的符合程度较高。这说明幼苗的各部分生长是密切相关的,在生产实践中,应当采取合理的水肥措施,促进苗木生长发育。
2.6 欧洲鹅耳枥2年生播种苗生物量积累及分配
于10月15日对生物量进行测定,结果见表4。从表4中可看出,幼苗地上部分总干质量仅有0.906 g,占总生物量的52.4%;根干质量最大,达到0.823 g。后期为入冬需要,苗木根系干物质量积累增多。

表4 欧洲鹅耳枥生物量分配Table 4 Biomass allocations of Carpinus betulus
对植物生物量的研究可反映植物的生长状态[14],展开植物器官生物量异速生长模式探讨具有一定研究价值。根据异速生长关系公式Y=axk,模拟出2年生欧洲鹅耳枥根与茎、根与叶、茎与叶鲜质量生物量之间的异速生长数学模式,结果见表5。相关系数均达到0.96以上,说明异速生长数学模式是适用的。苗木的根与叶的异速生长方程中指数K为1.098 8,两者的增长将近呈线性关系,叶的生长促进根系的生长,叶片生物量的增长,能增强根系吸收营养的能力,同时也能为根积累更多的干物质。茎与根的异速生长数学模式中,K为0.733 8,苗木的生长偏向于根的生长,说明幼苗的根系生长旺盛。叶与茎的异速生长数学模式中,K为1.187 9,且常数系数a为2.097,说明在生长过程中叶的生长比茎的生长更加旺盛,同时茎的疏导作用也促进了叶的生长。间接说明苗木各部分器官生长密切相关,相互促进,出现交替生长。

表5 欧洲鹅耳枥苗木各器官生物量的异速生长数学模型Table 5 Mathematical models of allometric growth of Carpinus betulus organs
3 讨论与结论
(1)欧洲鹅耳枥苗高、地径年生长基本呈现“慢—快—慢”的生长规律,阶段性明显。秦光华等[6]研究得到杨树无性系在生长期出现“慢—快—慢—快—慢”的生长规律,具明显阶段性。赵宏皂[15]、康华靖等[16]将苗木生长划分成3阶段:缓慢生长期、快速生长期和缓慢生长期。2年生欧洲鹅耳枥播种苗生长划分为3个时期:生长初期(5月18日之前)、生长盛期(5月18日~8月12日)和生长末期(8月13日~11月8日)。生长盛期生长量占全年总生长量近50%,这一时期水肥管理对苗木质量影响重大,天气炎热,需加强管理,如及时灌水、增设遮荫棚等栽培设施,力求为培育优质苗木。
(2)利用Logistic方程进行苗高、地径拟合,拟合效果显著。研究发现苗高进入生长盛期和生长后期的时间比地径早,苗高和地径之间存在负相关生长关系,联系紧密,相互影响。实践中可以通过相应栽培管理措施来调节各部分的生长[17-18]。黄三祥等[19]研究得出地温是影响沙地云杉苗高的主要因子。赖文胜[13]通过多元回归模型研究气象因子与生长规律间的关系,分析得到气温是影响苗木生长的主要因子。实验通过多元回归分析得到,影响苗高生长的主导环境因子是旬平均气温,而地径的生长与所选环境因子无较大关系。为培育优质苗木,提高苗木质量,可进一步探讨其它环境因子对地径生长的影响,科学育苗。
(3)利用回归分析得到主根、苗高和地径生长之间的回归方程,相关系数都达到显著水平,方程的符合度较高。可认为苗木的各部分生长具密切相关,生长过程中出现相互交替的现象。植物体不同器官如根、茎、叶等,出现生长速度不一致的情况称为异速生长现象。异速生长研究,有利于了解苗木生长发育规律及各器官生长发育间的比例,对于苗木培育有重要意义。幼苗生物量分配比例,地上部分∶地下部分=1∶1,茎∶叶=1.3∶1,说明幼苗根系生长量所占比重大,对于苗木影响明显。对苗木生物量可进一步进行模型构建研究,以便更深层次了解植物的生长趋势[20]。实际生产应用中,针对欧洲鹅耳枥茎干柔软、根系发达等特点,可加工成木材、盆景以及其它产品应用等,采取科学措施,实现效益最大化。
[1] Huntley B, Birks H J B. An atlas of past and present pollen maps for Europe: 0-13000 years ago[M].London: Cambridge University Press,1983:22-24.
[2] Ralska-Jasiewiczowa M, Miotk-Sapiganowicz G, Zachowica Jlatalowa M,et al.Carpinus betulus L.-Hornbeam[M]. Kraków:Polish Academy of Sciences, 2004: 69-78.
[3] Paula M, Carpinus L. The woody plant seed manual [M].Washington, D.C: USDA Forest Service, 2008: 328-332.
[4] 祝遵凌,许圆圆.欧洲鹅耳枥繁殖技术研究[J].安徽农业大学学报,2012,39(1):88-91.
[5] 章元明,盖钧镒.Logistic模型的参数估计[J].西南农业大学学报:社会科学版, 1994,(2):47-52.
[6] 秦光华,姜岳忠,马 玲,等.欧洲杨新无性系苗期年生长规律研究[J].陕西林业科技,2003,(3):8-12.
[7] 佘诚棋,杨万霞,兰传亮,等.青钱柳一年生实生苗的年生长规律[J].林业科技开发,2009,23(2):45-47.
[8] 洑香香,方升佐,旺红卫,等.青檀一年生播种苗的年生长规律[J].南京林业大学学报,2001,25(6):11-14.
[9] 董太祥.华北落叶松播种苗当年生长规律的研究[J].河北农业大学学报,1994,17(1):44-49.
[10] 邱学清,江希钿.回归积分在树木生长与气候关系中的应用[J].福建林学院学报,1989,9(4):418-422.
[11] 张春桃,朱小楼,蔡开锋,等.薹草的耐阴性研究及园林适用环境分析[J].北京林业大学学报, 2010,32(4):207-212.
[12] 唐守正.多元统计分析方法[M].北京:中国林业出版社,1984.
[13] 赖文胜.长序榆一年生播种苗的年生长规律[J].南京林业大学学报,2001,25(4):57-60.
[14] 杨蓓芬,李钧敏. 南方菟丝子寄生对3种入侵植物生长的影响[J]. 浙江大学学报:农业与生命科学版, 2012,38(2):127-131.
[15] 赵宏皂.北京杨毛白杨山海关杨苗期生长规律的观测[J].河北林业科技, 1998,(1):27-28.
[16] 康华靖,陈子林,周玉鸿,等.濒危植物香果树植物种子萌发及幼苗生长动态的比较[J].中南林业科技大学学报, 2011,31(1): 32-37.
[17] 赵 燕,董雯怡,张志毅.施肥对毛白杨杂种无性系幼苗生长和光合的影响[J].林业科学,2010,69(4):78-81.
[18] 马存世,李进军,李发鸿,等.干旱荒漠区育苗措施对白刺苗木生长的影响[J].中南林业科技大学学报,2012,32(5):1-5.
[19] 黄三祥,李新彬,林田苗,等. 沙地云杉育苗技术及苗木年生长规律研究[J]. 北京林业大学学报,2003,25(6):11-14.
[20] 曾伟生,肖前辉,胡 觉,等.中国南方马尾松立木生物量模型研建[J].中南林业科技大学学报, 2010, 20(5):50-56.
Annual growth dynamics of two-year-old seedlings ofCarpinus betulus
CHENG Long-xiaa, JIN Chun-zia, ZHU Zun-linga,b
(a. College of Landscape Architecture; b. College of Arts & Design, Nanjing Forestry University, Nanjing 210037, Jiangsu, China)
In order to reveal the annual dynamic growth ofCarpinus betulus, the two-year-old seedlings were taken as materials. The dynamic variations of seedling height, ground diameter and biomass accumulations were measured,and the correlation between the height, ground diameter and ecological factors were investigated. The results indicated that in the experiments, the growth of seedling height,ground diameter basically accorded with the rhythm of slow-fast-slow, which was fi tted by Logistic equation, and the growth stage of seedling height,ground diameter could be divided into three phases: the early stage, fast-growing stage and last stage; the seedling height in the fast-growing stage had one continuous rapid growth period, while the ground diameter had two intermittent growth peaks and the latter was more obvious than the previous; besides, there was a signif i cant linear relation between seedling height and ground diameter, but there was a negative correlation between the parts of seedlings; the correlation analysis of seedling height,ground diameter related with meteorological factors showed that the average temperature in half-moon was the dominate factor of seedling height, but other meteorological factors had no signif i cant effect on the growth of ground diameter.
Carpinus betulus; growth dynamics; seedling height; seedling ground diameter; seedling biomass
S723.1+31
A
1673-923X(2014)09-0026-05
2014-01-10
国家林业局“948”项目(2011-4-44);江苏省科技支撑计划项目(BE2012345);江苏省“青蓝工程”资助项目;江苏高校优势学科建设工程资助项目
程龙霞(1988-),女,安徽安庆人,硕士生,主要研究园林植物应用、园林植物栽培等;E-mail:zwahzchenglx@163.com
祝遵凌(1968-),男,河南固始人,教授,博士,主要研究园林植物应用、园林植物栽培等;
E-mail:zhuzunling@aliyun.com
[本文编校:谢荣秀]
