干旱胁迫对两种速生树种苗木生理指标的影响
2013-12-28谢德新李吉跃
潘 昕 ,谢德新 ,邱 权 ,李吉跃 ,苏 艳 ,何 茜
(1.华南农业大学 林学院,广东 广州 510642;2.莲化县林业局大乐坪林场,江西 莲花 337100)
干旱胁迫对两种速生树种苗木生理指标的影响
潘 昕1,谢德新2,邱 权1,李吉跃1,苏 艳1,何 茜1
(1.华南农业大学 林学院,广东 广州 510642;2.莲化县林业局大乐坪林场,江西 莲花 337100)
以竹柳和尾巨桉2种速生树种为试材进行干旱胁迫模拟盆栽试验,通过对比干旱胁迫后叶片中膜脂过氧化及保护酶和渗透调节物质含量的变化,对供选树种进行抗旱性能分析与评价,为在华南地区选育优良的抗旱、节水速生树种提供重要的理论支持和参考依据。试验结果表明, 两个树种在干旱胁迫下都表现出MDA、SOD、可溶性蛋白质含量先升高后降低的趋势,尾巨桉较竹柳都先达到峰值。POD活性呈先升高后降低再升高的趋势,尾巨桉的两个峰值都比竹柳出现的早,但竹柳的最大值比尾巨桉高出199.73 mg·g-1min-1。脯氨酸、可溶性糖含量为逐渐增加的趋势,在干旱胁迫24 d(重度干旱)达到最大值。含量表现出先增大后减小的规律,竹柳的峰值比正常水分条件下含量增加了4.60倍,高于尾巨桉的4.44倍。经过对6项指标的综合分析并结合植物的抗旱性表现,得出竹柳比尾巨桉的适应性强,抗旱性能较好。
竹柳;尾巨桉;速生树种;干旱胁迫;苗木生理指标;膜脂过氧化及保护酶;渗透调节物质
竹柳bamboo willow又名美国竹柳,具有抗性强、成材快、材质好、栽培效益高等特性,是一个亟待开发而十分难得的优质超速生新能源树种[1-2]。尾巨桉Eucalyptus urophylla×Eucalyptus grandis是我国南方速生丰产林的首选树种。本试验挑选了竹柳、尾巨桉两树种为试验对象。通过研究干旱胁迫对植物膜脂过氧化和保护酶活性、渗透物质的影响,得出不同植物品种的抗旱性能差异[3-6],国内外许多专家学者的大量的研究表明干旱胁迫下植物受到的伤害程度与此密切相关[7-10]。目前速生树种的抗旱性能研究较少,而此方面对竹柳和尾巨桉研究亦是鲜见报道。
本实验以竹柳和尾巨桉为试材进行干旱胁迫模拟盆栽试验,通过对比叶片中丙二醛(MDA)、超氧化物歧化酶(SOD)、过氧化物酶(POD)活性、脯氨酸含量、可溶性糖、可溶性蛋白质含量的变化,综合分析及评价其抗旱性能的差异,为在华南地区选育优良的抗旱、节水速生树种提供重要的理论支持和参考依据。
1 材料与方法
1.1 试验材料
试验材料为长势良好、形态特征相近的盆栽竹柳和尾巨桉1 a生苗木各30株,苗高、地径分别为:竹柳0.45±0.07 m、9.10±2.12 mm;尾巨桉:0.40±0.03 m、3.92±0.65 mm。于2011年5月在华南农业大学林学院温室内盆栽培育,花盆直径为25 cm,高30 cm,所用土壤为华南农业大学树木园内林地赤红壤。土壤田间持水量为(26.87±2.07)%,容重为 (1.34±0.07) g·cm-3。
1.2 研究方法
2011年7月1日浇透水后,对花盆进行套袋处理,以防止土壤水分蒸发而影响叶片蒸腾耗水的研究。对每株植株中生长位置和叶片朝向相同、大小相似的5~8片叶片进行挂牌标记,并于7月1日第一次采样,作为正常水分条件的对照,之后在干旱第3、6、9、12、15、19、24 d采样,同树种每次取3株的挂牌叶片进行试验,共3个重复。每次选用未取过叶片的完整植株进行取样,用以测定土壤含水量及各项生理指标。
1.3 测定指标与方法
1.3.1 土壤质量含水量
土壤质量含水量用土壤含水量快速测定仪Easttest(美国)测定。
1.3.2 丙二醛( MDA)的测定
丙二醛含量的测定用硫代巴比妥酸(TBA) 法[13]。
1.3.3 超氧化物歧化酶( SOD) 的测定
SOD活性测定用氮蓝四唑( NBT ) 法[13]。
1.3.4 过氧化物酶(POD)的提取和测定
进行实验室生化试验,发现该菌具有发酵葡萄糖、蔗糖、麦芽糖等功能,不能发酵阿拉伯糖、甘露醇,在对其进行MR、VP试验后,发现呈阴性,能使明胶液化,但是不具备运动能力,不能对尿毒酶进行水解。
POD活性采用愈创木酚比色法测定[14]。
1.3.5 脯氨酸含量的测定
脯氨酸含量的测定用:酸性茚三酮法[15]。
1.3.6 可溶性糖含量的测定
可溶性糖含量的测定用蒽酮法测定[16]。
1.3.7 可溶性蛋白含量测定
利用考马斯亮蓝G-250 法测定可溶性蛋白含量[17]。
2 结果与分析
2.1 干旱胁迫对苗木MDA的影响
图1表明,在正常水分条件下,竹柳的MDA含量较低,为 1.91 μmol·g-1,尾巨桉为 2.33 μmol·g-1。随着土壤含水量的减少,两种植物叶片中MDA含量表现出先增大后减小的规律。其中竹柳MDA含量在干旱胁迫初期先缓慢减少,再逐渐上升,而尾巨桉初期则一直上升。两种植物都在中度干旱时期达到峰值,尾巨桉在12 d最先达到4.81 μmol·g-1,竹柳在第 15 d 达到峰值 3.90 μmol·g-1,尾巨桉比竹柳的高出23.3%。当植物遭受重度干旱胁迫时(19~24 d),MDA含量又都迅速下降,第24 d两种苗木都已略高于正常水分条件的值,竹柳为 2.54 μmol·g-1,尾巨桉为 2.96 μmol·g-1。干旱胁迫对不同苗木叶片MDA含量有不同的影响,但竹柳MDA含量的峰值与对照增幅为104.2%,尾巨桉为106.4%,二者相近。
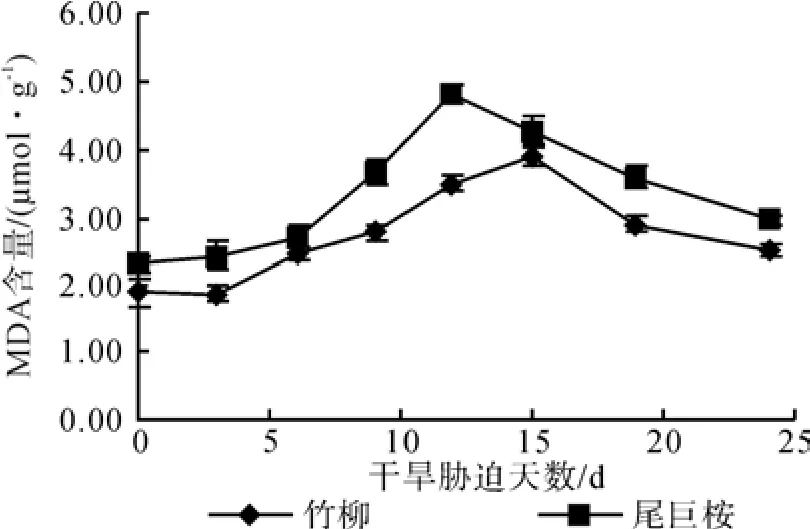
图1 干旱胁迫对苗木MDA含量的影响Fig.1 Effects of drought stress time on MDA content of seedlings
2.2 干旱胁迫对苗木SOD活性的影响
从图2可以看出,在正常条件下,两种植物的SOD活性有一定差距,竹柳41.20 U·g-1,尾巨桉61.30 U·g-1。在水分胁迫下,供试苗木的SOD活性都表现出先增大后减小的规律,其中尾巨桉SOD活性峰值出现在干旱胁迫的第9 d(轻度干旱时期),为234.83 U·g-1,竹柳出现峰值的时间稍晚,在第12 d(中度干旱时期)达到258.00 U·g-1,可知竹柳比尾巨桉高出9.9%,与对照相比,竹柳增加526.21%,尾巨桉较低增加了283.08%。在重度水分胁迫下,苗木叶片的SOD活性都逐渐下降,竹柳降低6.95 U·g-1,尾巨桉降低30.73 U·g-1,在第24 d都低于正常水分条件下的SOD活性值。
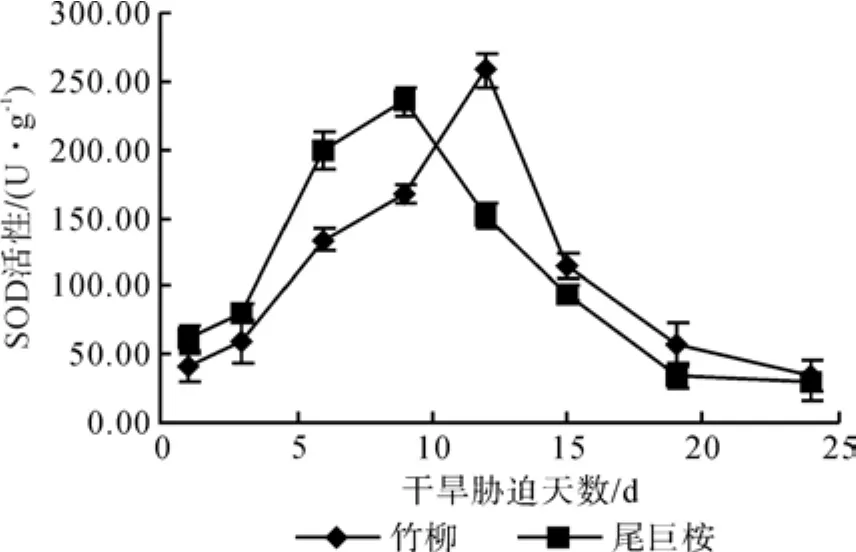
图2 干旱胁迫对苗木SOD含量的影响Fig.2 Effects of drought stress time on SOD content of seedlings
2.3 干旱胁迫对苗木POD活性的影响
由图3可知,对照组中两种植物的POD活性平均为153.64 mg·g-1min-1。当植物遭受干旱胁迫后, POD活性表现出先增大后减小再增大的波浪式变化,且都高于正常水分条件下的活性含量值。尾巨桉呈现“双峰曲线”,在水分胁迫第9 d(轻度干旱)达到第一个峰值177.60 mg·g-1min-1,而竹柳在水分胁迫第12 d(中度干旱)才达到第一个峰值,为129.67 mg·g-1min-1,尾巨桉的含量比竹柳高出37.0%。随着干旱胁迫的加深POD活性逐渐下降,但仍高于正常水分条件下的活性值,随后又逐渐上升。第二次峰值依旧是尾巨桉先达到,第19 d:634.67 mg·g-1min-1,而竹柳在第24 d才达到第二个峰值,为834.40 mg·g-1min-1,而且还有继续上升的趋势。与正常的POD活性值相比,竹柳的最大值增加4.43 倍,高于尾巨桉的2.48 倍。
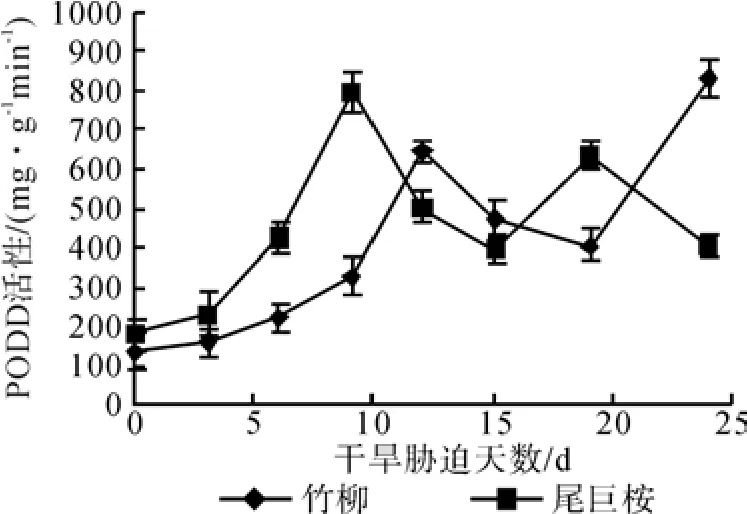
图3 干旱胁迫对苗木POD含量的影响Fig. 3 Effects of drought stress time on POD content of seedlings
2.4 干旱胁迫对苗木脯氨酸含量的影响
图4可以看出对照组中,两种植物叶片脯氨酸含量有一定差距,竹柳为23.57 μg·g-1,尾巨桉54.20 μg·g-1。随后植物叶片脯氨酸含量不断增加。在干旱胁迫初到中期(0~12 d)增加较缓慢,增幅维持在40%左右,且竹柳的含量始终低于尾巨桉,干旱胁迫12 d后脯氨酸含量开始大幅度提升。尾巨桉在24 d达到峰值311.49 μg·g-1。竹柳含量在第15 d大幅度上升,干旱胁迫19 d后超过尾巨桉,24 d 达到峰值 392.78 μg·g-1。竹柳的最大含量是尾巨桉的1.26 倍。峰值与对照相比,竹柳增加了15.66 倍,尾巨桉4.78 倍,可见水分胁迫对苗木叶片中脯氨酸含量有较大的影响。
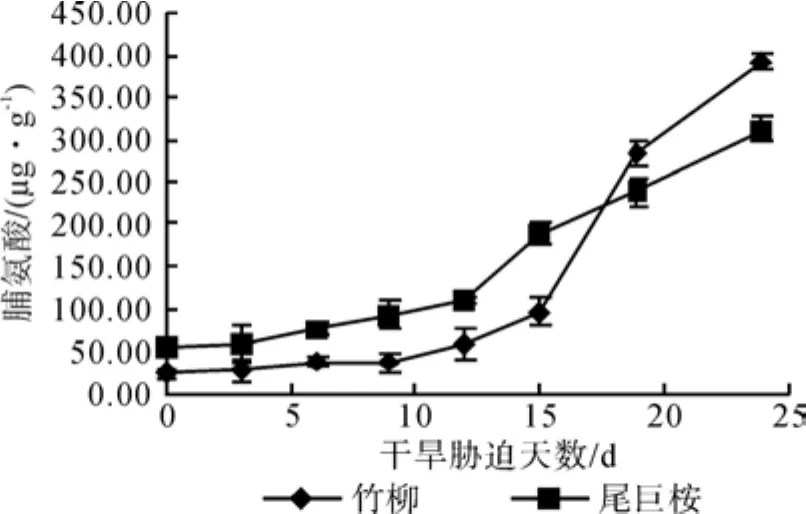
图4 干旱胁迫对苗木脯氨酸含量的影响Fig. 4 Effects of drought stress time on proline content of seedlings
2.5 干旱胁迫对苗木可溶性糖含量的影响
图5表明,在正常水分条件下,两种植物叶片所含可溶性糖水平基本一致,平均为11.28 mg·g-1。干旱胁迫0~6 d内,增幅不大且种间含量差异不显著,0~9 d竹柳始终高于尾巨桉。在重度干旱时期(24 d)达到峰值,竹柳38.87 mg·g-1,比尾巨桉高出4.19 mg·g-1。较正常水分条件相比,尾巨桉叶片中可溶性糖含量增加24.1 mg·g-1,竹柳增加26.89 mg·g-1。在水分胁迫0~9 d,2种苗木都增加缓慢,9 d后尾巨桉开始迅速增加,且高于竹柳,而竹柳在第12 d才开始大幅度增加,15 d后超过尾巨桉。与对照相比较,竹柳的增幅(224.46%)略高于尾巨桉(218.34%)。
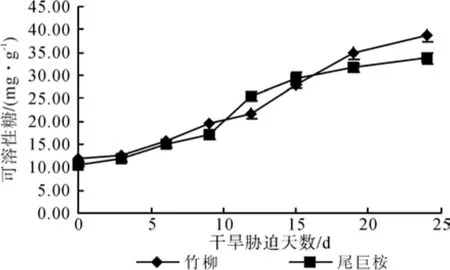
图5 干旱胁迫对苗木可溶性糖含量的影响Fig. 5 Effects of drought stress time on soluble sugar content of seedlings
2.6 干旱胁迫对苗木可溶性蛋白质含量的影响
由图6可知,在干旱胁迫程度逐渐加深的情况下,苗木叶片可溶性蛋白质含量总体表现出先增大后减小的规律,且两种苗木间存在明显的差异。正常水分条件下,竹柳和尾巨桉的可溶性蛋白质含量接近,平均为1.14 mg·g-1,第3 d开始,尾巨桉的可溶性蛋白质含量迅速增加,但0~6 d始终低于竹柳,第9 d达到峰值5.82 mg·g-1,之后开始下降。竹柳在干旱胁迫第12 d达到峰值6.78 mg·g-1,之后也逐渐降低但其含量一直高于尾巨桉。干旱胁迫的过程中可溶性蛋白质含量始终高于对照值,竹柳的峰值含量比尾巨桉高16.49%,竹柳的峰值比正常水分条件下可溶性蛋白质含量增加了4.60倍,高于尾巨桉的4.44倍。
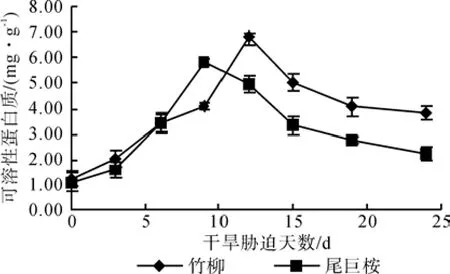
图6 干旱胁迫对苗木可溶性蛋白含量的影响Fig.6 Effects of drought stress time on soluble protein content of seedlings
3 结论与讨论
植物在逆境下往往发生膜质过氧化作用,MDA是膜质过氧化作用的最终分解产物,其含量与植物遭受逆境伤害的程度成正比[7,18]。本试验中正常水分条件下竹柳MDA含量少,中度干旱时期峰值更迟出现,且含量低于尾巨桉。SOD主要功能是清除活性氧,以保持自由基和清除剂之间的平衡[19]。竹柳在遭受干旱胁迫时能够迅速分解体内产生的超氧自由基,减缓膜脂过氧化速度,充分反映出该树种对干旱环境有较强的适应性及自我调节能力。POD与SOD、过氧化氢酶(CAT) 联合作用参与清除活性氧及过氧化物自由基的活动[20]。与尾巨桉相比,竹柳最高值较正常水分条件下增加高达4.43倍。这与黄高峰等[21]在对干旱胁迫对菊芋苗期叶片保护酶活性及膜脂过氧化作用的影响的发现一致。由膜脂过氧化及保护酶指标可知竹柳表现出更强的抗旱能力。
干旱逆境条件下,植物通过渗透调节降低水分胁迫对苗木的伤害,脯氨酸含量越高,显示其抗旱能力越强[22-24]。试验中脯氨酸含量的峰值与对照相比,竹柳远高于尾巨桉10.88 倍,这与王启明等[25]对干旱胁迫下大豆苗脯氨酸含量变化的试验结果一致。在重度干旱胁迫时期竹柳的可溶性糖含量比尾巨桉高出4.19 mg·g-1,与正常水分条件相比较,竹柳的增幅高于尾巨桉,说明竹柳对干旱环境的自我调节能力较尾巨桉强。
为了避免胁迫造成伤害,会诱导产生一些抗逆蛋白质,高含量的可溶性蛋白,以抵抗干旱引起植物体内活性氧的积累,抗旱性强的植物种类或品种的可溶性蛋白含量较高[26-27]。试验中,苗木都表现出可溶性蛋白质先增加后降低的变化规律,这可能是因为随着干旱时间的延长, 植物对干旱的忍耐能力降低, 植物体内代谢受阻, 导致蛋白质降解。干旱胁迫第12 d后可溶性蛋白质含量竹柳始终高于尾巨桉,表现出更强的抗旱性。
由上述的分析可以得出,在干旱胁迫下,较尾巨桉而言,竹柳具有较为理想的抗旱特性。希望本试验结果能为在华南地区选育优良的抗旱、节水的速生树种提供理论依据,为建立更加完善的抗旱指标体系做好基础。
[1] 王子成. 用材与绿化兼得树种——速丰竹柳 [J]. 中国林业,2011,(22):38-41.
[2] 吕德文. 竹柳引种试验初报 [J]. 防护林科技, 2011,(5): 42-43.
[3] Bowler C, Van Montagu M, Inze D. Superoxide dismutase and stress tolerance [J]. Annual Review of Plant Physiology and Plant Molecular Biology, 1992, 43: 83-116.
[4] 王新建, 谢碧霞, 何 威, 等. 干旱胁迫对4 种豫楸1 号嫁接苗膜脂过氧化作用的影响 [J]. 中南林业科技大学学报, 2008,28(6): 30-34.
[5] Mendy MC. Active oxygen species in plan t defense against pathogens [J]. Plant Physiol, 1994, 105: 467-472.
[6] 潘 昕, 李吉跃, 王军辉, 等. 干旱胁迫对青藏高原4种灌木生理指标的影响 [J]. 林业科学研究, 2013,26 (3): 352-358.
[7] 谷文众, 刘 杨, 谷振军. 干旱胁迫对金盏菊膜脂过氧化及保护酶活性的影响 [J]. 经济林研究, 2009, 27(3): 79-81.
[8] Nuccio ML, Rhodes DR, McNeil S D, et al. Metabolic engineering of plants for osmotic stress resistance [J]. Curret Opinion Plant Biology, 1999, 2(2): 128 - 134.
[9] 梁文斌, 蒋丽娟, 马 倩, 等. 干旱胁迫下光皮树不同无性系苗木的生理生化变化 [J]. 中南林业科技大学学报, 2011,31(4): 13-19.
[10] 包 卓, 孟祥英, 张晓松, 等. 干旱胁迫对5 种园林绿化植物生理生化的影响 [J]. 经济林研究, 2010, 28(1): 46-50.
[11] 陈晓远, 高志红, 罗远培, 等. 不同土壤水分冬小麦根、冠关系及其对叶片水分利用效率的影响 [J]. 中国生态农业学报,2005, 13(2): 134-137.
[12] 蒲金涌, 姚小英, 贾海源, 等. 甘肃陇西黄土高原旱作区土壤水分变化规律及有效利用程度研究 [J]. 土壤通报, 2005,36(4): 483-486.
[13] 邹 琪. 植物生理学试验指导 [M]. 北京: 中国农业出版社,2000,98-100.
[14] 张志良. 植物生理学试验指导 [M]. 北京: 高等教育出版社,2001,123-124.
[15] 白宝璋, 汤学军. 植物生理学测试技术 [M]. 北京: 中国科学技术出版社, 1993,76-157.
[16] 王晶英, 敖 红, 张 杰, 等. 植物生理生化实验技术与原理[M]. 哈尔滨: 东北林业大学出版社, 2003,12-13.
[17] 高俊凤. 植物生理学实验技术 [M]. 北京: 世界图书出版社,2000,137-138.
[18] 潘 昕, 李吉跃, 苏 艳, 等. 干旱胁迫对华南地区三种苗木膜脂过氧化及保护酶活性的影响 [J]. 广东林业科技,2012,28(1): 13-18.
[19] 刘 宁, 高玉葆, 贾彩霞, 等. 渗透胁迫下多花黑麦草叶内过氧化物酶活性和脯氨酸含量以及膜相对透性的变化 [J]. 植物生理学通讯, 2000, 36(1): 11-14.
[20] 王宝山, 赵思齐. 干旱对小麦幼苗膜脂过氧化及保护酶的影响 [J]. 山东师范大学学报:自然科学版, 1987, 2(1): 29 -39.
[21] 黄高峰, 王丽慧, 方云花, 等. 干旱胁迫对菊芋苗期叶片保护酶活性及膜脂过氧化作用的影响 [J]. 西南农业学报, 2011,24(2): 552-555.
[22] Guehl J M, Clement A, Kaushal P, et al. Planting stress, water status and nonstructural carbohydrate concentrations in Corsican pine seedlings [J]. Tree Physiol, 1993, 12(20): 173 - 183.
[23] 潘 昕, 邱 权, 李吉跃, 等. 干旱胁迫下华南地区3种苗木渗透调节物质的动态变化 [J]. 华南农业大学学报, 2012,33(4): 519-523.
[24] 胡晓健, 喻方圆, 刘建兵. 干旱胁迫对不同种源马尾松苗木针叶内可溶性糖含量的影响 [J]. 南京林业大学学报:自然科学版, 2009, 33(5): 55-59.
[25] 王启明, 马原松. 不同抗旱品种大豆苗中脯氨酸累积的差异[J]. 商丘职业技术学院学报, 2005(2): 63-64.
[26] 吴志华, 曾富华, 马生健. ABA对PEG胁迫下狗牙根可溶性蛋白质的影响 [J]. 草业学报,2004,13(5): 75-78.
[27] Xiong L M, Schumaker K S, Zhu J K. Cellsigna ling during sold,drought, and salt stress [J]. Plant Cell 2002, 14: 165-183 .
Effects of drought stress on seedlings physiological indexes of two fastgrowing tree species
PAN Xin1, XIE De-xin2, QIU Quan1, LI Ji-yue1, SU Yan1, HE Qian1
(1.College of Forestry, South China Agriculture University, Guangzhou 510642, Guangdong, China; 2.Daleping Forest Farm, Forestry Bureau of Lianhua County, Lianhua 337100, Jiangxi, China)
By taking bamboo willow and Eucalyptus urophylla× Eucalyptus grandis seedlings as the tested materials, the drought stress experiments were conducted with pot-culture method. Through comparing the changes of membrane lipid peroxidation and protective enzyme and osmoregulation substances in the seedlings after stressed by drought, the of drought-resistance performance of the tested trees were analyzed and evaluated. The results show that after drought stress treatments, the contents of MDA, SOD and soluble protein in the two tree species increased at fi rst and then decreased; the three values of E. urophylla× E. grandis reached the peak values earlier and higher than that of bamboo willow; the POD activity increased at fi rst, then decreased and fi nally increased, the peaks of E. urophylla× E.grandis appeared earlier than that of bamboo willow, but the maximum value of bamboo willow was higher by by 199.73 mg·g-1min-1than that of E. urophylla× E. grandis; their proline and soluble sugar contents gradually increased, peaked in 24th day(severe drought), showed a law of increase fi rst then decrease, the peak value of bamboo willow increased by 4.60 times than that of the normal moisture conditions;Through the comprehensive analysis of six indicators combined with the drought resistance performance of plants, it was concluded that the bamboo willow has strong adaptability and well drought resistance performance than E. urophylla× E. grandis.
bamboo willow; Eucalyptus urophylla× Eucalyptus grandis; drought stress; seedlings physiological indexes; membrane lipid peroxidation and protective enzyme; osmoregulation substances
S718.43
A
1673-923X(2013)10-0084-05
2012-11-08
广东省科技计划项目“桉树人工林精准施肥配套技术研究”(项目编号:2010B020303006),亚热带农业生物资源保护与利用国家重点实验室开放课题(KSL-CUSAb-2012-07)
潘 昕(1989-),女,福建永安人,硕士研究生,主要从事森林培育理论与技术方面的研究;
E-mail:xiaowandoujiajia@126.com
李吉跃(1959-),男,四川金堂人,教授,博士生导师,主要从事森林培育及栽培生理生态方面的研究;
E-mail:ljyymy@vip.sina.com
[本文编校:文凤鸣]
