丛枝菌根真菌最新分类系统与物种多样性研究概况
2013-12-19王宇涛辛国荣李韶山
王宇涛,辛国荣,李韶山,*
(1.广东省高等学校生态与环境科学重点实验室,华南师范大学生命科学学院,广州 510631;2.生物防治国家重点实验室,中山大学生命科学学院,广州 510275)
丛枝菌根真菌(arbuscular mycorrhizal fungi,AMF)是地球上分布最广泛的一类植物共生真菌,能够与大部分高等植物的根系形成共生关系[1]。它们也是一类非常古老、起源和演化相对独立的微生物;化石证据表明,最早的AMF起源于大约4.6亿年之前,几乎与陆生植物同时出现在地球上,是植物从水生环境成功登陆的两个伴生物种之一[2]。由于AMF在农林、环境等领域的巨大应用潜力,AMF物种多样性的研究一直受到较高的关注。近年来的研究发现,AMF能够通过直接和间接作用对生态系统中植物群落的多样性和生产力产生深刻影响[3];因此,AMF物种多样性的研究愈发显得重要。不过,由于AMF只有与活体寄主植物的根系形成共生关系之后才能完成其生活史,极大增加了对其分类以及多样性研究的难度;再加上研究手段及其它因素的限制,阻碍了AMF物种多样性的研究进展。近年来,研究方法的改进与新技术的发展为AMF物种多样性的研究提供了极好的机遇。本文综述了近年来在AMF分类系统和全球物种数量、AMF物种多样性影响因素以及AMF物种多样性研究方法3个方面的最新研究进展,以期为国内同行提供参考。
1 AMF的分类系统及全球物种数量
1.1 AMF的最新分类系统
AMF分类系统的建立是开展AMF物种多样性研究重要的基础性工作。早期AMF的分类研究严重依赖于对AMF形态特征的观察。Morton等[4]依据对27类AMF结构特征(包括AMF孢子及根内的形态)的综合分析,将全部AMF归入新设立的球囊霉目(Glomerales,隶属于接合菌纲)。Schüβler等[5]依据对18S rDNA分子进化分析的结果,将AMF的分类地位提升至门,并归入新设立的球囊霉门(Glomeromycota,包括所有已知AMF种类及一种目前所知甚少的内生真菌(Geosiphon pyriforme))。刚设立时球囊霉门[5]包含1个纲(Glomeromycetes)4 个目(Glomerales,Diversisporales,Paraglomerales,Archaeosporales)7个科(Glomeraceae,Gigasporaceae,Acaulosporaceae,Diversisporaceae,Paraglomeraceae,Archaeosporaceae,Geosiphonaceae)9 个属(Glomus, Gigaspora, Scutellospora, Acaulospora, Entrophospora, Diversispora, Paraglomus, Archaeospora,Geosiphon);此后经过不断补充和调整[6]又新增了 4个科(Claroideoglomeraceae,Entrophosporaceae,Pacisporaceae,Ambisporaceae)9 个 属 (Funneliformis,Rhizophagus,Sclerocystis,Claroideoglomus,Racocetra,Pacispora,Otospora,Redeckera,Ambispora),使AMF(球囊霉门)分类系统包含了1个纲4个目11个科18个属(见 http://www.amf-phylogeny.com)。最近,Oehl等[7]根据对 AMF 的 DNA 序列(包括 rDNA 和 β-微管蛋白基因序列)以及形态学特征的综合分析,进一步对球囊霉门分类系统进行了调整,形成目前包含3个纲5个目14个科26个属的最新AMF分类系统(表1)[7]。
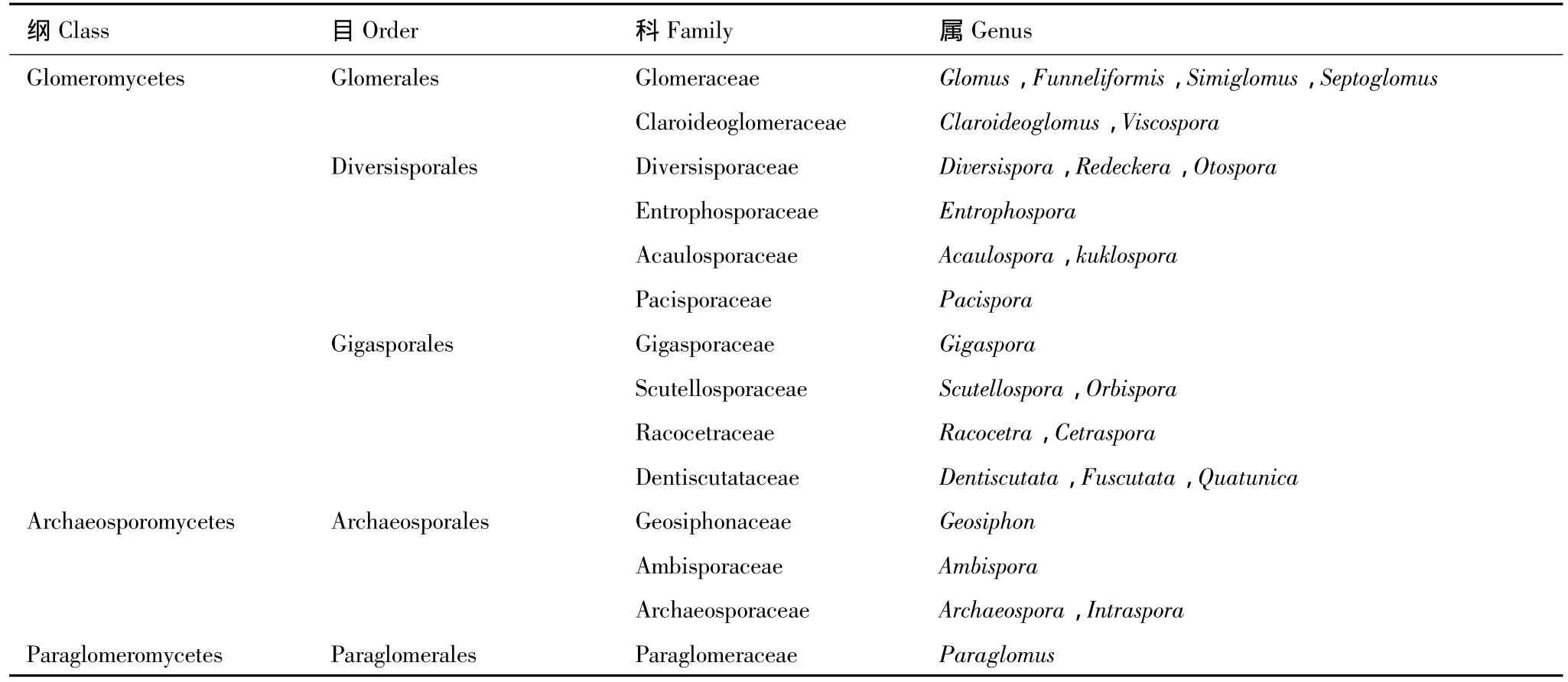
表1 丛枝菌根真菌(arbuscular mycorrhizal fungi(AMF),球囊霉门)最新的分类系统[7]Table 1 Updated classification system of arbuscular mycorrhizal fungi(AMF,Glomeromycota)[7]
1.2 AMF的全球物种数量
地球上大约有70%—80%的高等植物种类能够与AMF形成共生关系[1]。到目前为止,在从热带雨林到极地冰原绝大多数类型的生境中均发现了 AMF的存在[8]。大量研究分别报道了森林[9]、草原[10]、农田[11]、高原[12-13]、高山草甸[14]、沙漠[15]、干热河谷[16]、盐碱地[17]、工业污染区[18]、滨海红树林[19-20]等生态系统中AMF与寄主植物的共生关系。AMF丰富的寄主和生境多样性充分证明它们对不同生境类型的适应性,以及它们对部分胁迫环境的耐受能力。
尽管AMF具有极高的寄主和生境类型多样性,迄今为止全球范围内报道的AMF物种数量要少于250个种(http://www.amf-phylogeny.com),远远低于其寄主植物的种类多样性。一个通常的解释是AMF具有较低的寄主偏好性和专一性;不少研究的结果在一定程度上支持了这一观点[10,21]。然而也有明确的证据显示,至少部分AMF种类与寄主植物之间存在着明显的寄主偏好性[22];还有报道称,AMF在寄主植物根内的生长模式[23]以及它们对寄主植物的生态效应[24]与两者的种类(株系)密切相关,进一步证实了AMF寄主专一性和偏好性的存在。因此,用AMF较低的寄主偏好性和专一性来解释AMF种类多样性远远低于其寄主的种类多样性目前还存在较大的争议。
近年来,有越来越多的证据表明AMF在全球的种类数量可能被严重低估[20,25]。Börstler等[14]根据在两个高山草甸生态系统中AMF的多样性调查结果推测,全球范围内AMF种类至少要超过1250种。Öpik等[26]采用整合分析的方法探讨了AMF在全球的群落多样性,发现在已调查的52种寄主植物根内一共定殖有95个AMF的分类单元。Wang等[20]甚至在高盐、高湿的滨海红树林生境中也检测到很高的AMF物种多样性:他们从在不同潮间带均有分布的3种寄主植物根内一共检测到23个AMF分类单元。还有不少调查试验的结果[27-31]也表明,在许多类型的生态系统中都存在较高的AMF种类多样性(表2);尤其是在运用分子学手段进行AMF物种多样性的调查当中,往往出现较大比例未知种类的AMF序列[3,20,25];这些结果进一步支持了AMF在全球范围内的物种多样样可能远远超过目前所发现种类的观点。有学者甚至提出,全球范围内AMF的种类多样性可能与其寄主植物种类的多样性处于一个相当的水平[9,32]。尽管AMF在全球范围内的物种多样性水平还需要进一步的研究才能明确,AMF的物种多样性很可能要远高于目前所发现的AMF种类。

表2 检测到较高丛枝菌根真菌(arbuscular mycorrhizal fungi,AMF)物种数量的典型研究Table 2 Typical reports in which high level of arbuscular mycorrhizal fungi(AMF)species richness were detected
2 AMF物种多样性的影响因素
在生态系统中能够影响AMF物种多样性的因素很多;每一处生境中AMF的物种多样性及其分布特征都是由各种生物和非生物因素共同作用的结果。
2.1 寄主植物
作为生活在植物根内的专性共生真菌,AMF的物种多样性水平受到寄主植物的影响是显而易见的。由于不同植物在生理代谢、根系形态结构与分泌物等方面可以存在较大差异,这必然会影响到AMF对寄主植物的识别和感染,进而影响到AMF的生长发育、产孢、分布、群落组成、多样性等[33]。
到目前为止,关于寄主植物对AMF物种多样性影响的研究通常都是在植物种的水平进行的探讨。不少研究发现,在相同或者类似的生境中,不同种类寄主植物根内或根际的AMF物种多样性通常存在较大的差异[34],证实了寄主植物种类对AMF物种多样性的重要影响。然而,也有研究发现植物种类对AMF多样性并无明显影响[32]。因此,寄主种类对AMF物种多样性的影响应该是与特定的生境、植物种类以及AMF类型密切相关的[20]。在植物种以下水平,关于不同品种(基因型)寄主植物对AMF物种多样性影响的研究则尚不多见。郭绍霞和刘润进[35]调查了同一生境不同牡丹(Paeonia suffruticosa)品种根际土壤中AMF孢子的密度、种属组成、物种多样性指数等,发现不同品种的牡丹能促进或抑制不同AMF种类的生长发育,进而影响根际AMF的多样性和群落特征。Hannula等[36]采用末端限制性长度多态性(T-RFLP)的方法监测了大田控制试验中六个不同土豆(Solanum tuberosum)品种根围AMF的群落结构及多样性,发现寄主品种的差异对根围AMF的多样性和群落结构并无明显的影响。因此,不同品种或基因型的植物对AMF物种多样性是否存在影响、影响的强度及影响机制尚需进一步的研究才能明确。近年来,在群落水平研究寄主植物对AMF物种多样性的影响成为菌根领域受到广泛关注的课题。由于不同种类的寄主植物对AMF的偏好性、专一性及作用方式通常存在较大差异,植物的群落结构及多样性必然会对该生境中AMF的种类组成和物种多样性产生深刻的影响。不少研究发现,在植物多样性高的生境中AMF的物种多样性水平往往也较高[9,16];因此,寄主植物的群落多样性可能是决定AMF物种多样性的重要因素[33,37-38]。然而,对于这一结果也可以有不同的解释,即AMF的物种多样性可能也是影响植物群落多样性的一个重要因子[39]。事实上,AMF的物种多样性和植物群落多样性应该是一种相互影响、相互作用的关系[3]。
2.2 环境因子
环境因子对AMF物种多样性的影响一直是菌根研究领域的重要内容。在影响AMF多样性的各种环境因素当中,报道得最多的是土壤状况对AMF物种多样性的影响。迄今为止,关于土壤理化因子对土壤中AMF孢子种类组成影响的研究已经很多报道[34]。Oehl等[40]调查了16个不同生态样区土壤中AMF孢子的种类组成及多样性,发现土壤类型能强烈影响AMF孢子的群落结构及多样性水平。Antoninka等[41]在一处草地生态系统中开展的研究表明,长期(7a)施用N肥能显著降土壤中AMF孢子的密度及种类多样性。Gosling等[42]的研究发现,向农田生态系统中添加适量有机质能够明显提高农田土壤中AMF孢子种类的多样性。此外,关于土壤理化因子对寄主根内AMF物种多样性影响的研究也有部分报道。Wirsel[43]采用分子学方法调查了一处湿地生态系统中分布在不同类型土壤中的芦苇(Phragmites australis)根内AMF感染强度和物种多样性,发现寄主植物根围土壤的理化特征是芦苇根内AMF感染强度和多样性的决定性因子。van Diepen等[44]在一处硬木森林中开展的研究发现,长期(>12a)施用外源N能够强烈影响该生境中AMF的群落结构,并显著降低寄主植物根内 AMF 的物种多样性。Zarei等[45]和 Martínez-García等[15]的研究也都表明,土壤理化因子是影响寄主植物根内以及根际AMF的群落结构和多样性的重要影响因素。因此,土壤性状无论是对于根际AMF孢子群落还是根内AMF种类的多样性水平均具有重要的影响。
除了土壤理化因子之外,还有不少研究报道了其它环境因素对AMF物种多样性的影响。Wang等[20]采用分子学手段调查了一处红树林湿地生态系统中的AMF多样性,发现淹水强度是影响湿地植物根内AMF多样性的决定性因子。Gai等[46]调查了西藏色拉山在不同海拔分布的寄主植物与AMF的共生状况,发现随着海拔的增加,AMF的感染强度、根际土壤中的孢子密度及种类多样性均呈现下降的趋势。Wu等[47]调查了富士山在不同海拔高度均有分布的四种寄主植物根内以及根际的AMF物种多样性,发现AMF的物种多样性水平呈现随海拔升高而下降的规律;他们认为,这可能是由不同海拔高度的植物群落结构、土壤理化性质存在差异所导致的。还有研究发现,季节变化对寄主植物根内AMF的物种多样性具有显著的影响[9-10];Dumbrell等[31]运用最新的焦磷酸测序技术检测了一处温带草原生境中部分寄主植物根内的AMF群落结构,发现寄主根内AMF的物种多样性及群落组成在冬季和夏季存在明显的差异。季节变化对AMF多样性的影响主要是由于寄主植物根系的物候学特征、寄主植物以及AMF孢子的周边环境因子(如温度、日照时间、水分等)等随季节变化而引起的[9-10,31]。近年来,关于全球气候变化(如气温增加、大气CO2浓度升高等)对AMF多样性影响的研究受到关注。不少研究表明,在一定范围内CO2浓度的升高能够促进寄主植物的光合作用,增加碳水化合物向地下部的供应[48],进而提高根际AMF的侵染强度和物种多样性[41,49]。还有研究发现,温度升高同样能够增加寄主植物根围AMF的侵染强度和物种多样性[50];一方面是由于温度升高直接促进了AMF根外菌丝的分枝与伸长[51],另一方面则是由于温度升高促进了寄主植物的光合作用[49,51]。不过也有报道称,温度升高能够提高土壤中N、P营养的有效性[52-53],进而减少寄主根围AMF的物种多样性。总体上,关于全球气候变化对AMF物种多样性的影响尚未阐明;通过较大的时间和空间尺度来探讨全球气候变化对AMF物种多样性的影响,将是今后AMF物种多样性研究领域值得关注的研究方向。
2.3 人类活动
人类活动能够通过直接或者间接的方式对生态系统中AMF的物种多样性产生深刻的影响。有研究报道,人类活动的干扰会显著降低生态系统中AMF的物种多样性[54]。Helgason等[55]的研究结果表明,在农田生态系统中,人为干扰程度大的农田中AMF的物种多样性水平通常较低。Lin等[56]利用454焦磷酸测序技术测定了华北地区长期(>20a)采用不同施肥方式的农田中AMF的多样性及群落结构,发现长期施肥处理(尤其是施用P肥和N肥)明显降低了农田生境中的AMF多样性水平。Schnoor等[57]在一处半人工草地生态系统中的研究发现,对土壤进行机械翻耕强烈影响了该生境中AMF的群落结构,并显著降低了AMF的物种多样性。Daniell等[58]和Rosendahl等[59]的研究结果也表明,频繁的人为干扰对寄主植物根内AMF的多样性和群落结构均存在巨大影响。然而,关于人为干扰对AMF多样性的影响也有不同的报道。Öpik等[29]采用分子学手段调查了一处针叶林生境中5种寄主植物根内AMF的群落结构和多样性,发现一定强度的人为扰动对寄主根内AMF的群落结构和多样性均无明显的影响。Hijri等[60]和Vallino等[61]也报道了类似的结果。显然,人类活动对生态系统中AMF物种多样性的影响应该是与人为干扰的方式和强度密切相关的。
3 AMF物种多样性的研究方法
传统对AMF物种多样性的调查是基于寄主植物根际土壤中AMF孢子的形态学鉴定,包括直接从寄主根际分离或者是利用菌根敏感型植物诱导培养获得的AMF孢子[1]。迄今为止报道的AMF种类均是基于该方法鉴定和发现的。然而,随着对AMF多样性研究的深入,形态学方法逐渐暴露出以下不足。一方面,根际土壤中的AMF孢子无法准确反映寄主根内AMF的群落结构,因而无法对植物根内的AMF群落动态进行实时监测。由于AMF存在部分不产孢子的种类,基于根际土壤中AMF孢子的多样性结果往往低估了AMF真实的多样性水平。此外,土壤中还存在一些取食AMF孢子的生物类群[62]以及以AMF孢子为寄主的寄生生物[63],导致土壤中AMF孢子群落组成与寄主根内的AMF种类组成存在较大差异。另一方面,土壤中AMF孢子的群落结构及多样性水平的调查结果强烈受到采样时间的影响。Tian等[64]调查了我国华北农田生态系统中AMF孢子的群落动态,发现AMF孢子的种类组成及多样性水平明显受到种植作物(小麦,Triticum aestivum)生长发育期的影响:AMF孢子的种类多样性在植物需P量高的生长期要明显高于其它时期。Li等[65]和Cuenca等[66]的研究结果也表明,采样时间对AMF孢子多样性的调查结果具有重要的影响。最后,AMF孢子的形态学鉴定在很大程度上依赖于研究人员的经验和文献资料的获取,对于初学者往往具有较大的难度;以形态特征为依据的AMF鉴定方法在一定程度上具有不一致性、偶然性和局限性[16]。
自Simon等[67]首次运用分子学技术调查寄主根内AMF的多样性以来,以PCR为基础的分子学方法已广泛应用于AMF的种类鉴定、多样性调查以及群落结构的研究。一般情况下,该方法首先对AMF的核糖体小亚基基因、内转录间隔区及/或核糖体大亚基基因片段进行特异性扩增,然后运用变性梯度凝胶电泳[68]、单链构象多态性[69]、限制性片段长度多态性[70]或者末端限制性片段长度多态性[71]等技术对扩增片段进行初步的分型分析,最后测定目的片段的DNA序列,并开展分子系统发育分析。相比基于AMF孢子形态学鉴定的传统手段,分子学方法最大的优点在于能够直接检测寄主根内的AMF群落结构和多样性;并且其实验操作更为标准、结果也更为可信[72]。因此,它们的运用极大拓展了对不同生境中AMF物种多样性的认识,加深了对AMF物种多样性的理解。
另一方面,尽管分子学技术较好的弥补了传统形态学方法的缺陷,它们在应用于AMF物种多样性的研究中同样面临不少问题。首先,尽管目前已经报道了不少用于AMF序列特异性扩增的引物(例如,AM1-NS31[55],AML1-AML2[73]),这些引物大多只能扩增部分的已知 AMF 种类[74],导致分子学调查的结果无法全面反映目标寄主根内的AMF多样性。其次,尽管技术上的改进已经降低了Sanger测序的成本,在样本量较大的情况下AMF多样性的分子学调查仍然价格不菲。因此,在运用分子学技术对寄主植物根内的AMF多样性进行调查时往往只能选取生境中少数代表性的植物种类进行检测[26],难以全面反映寄主植物根内的AMF多样性。最后,AMF多样性的分子学研究最终获得的是AMF的种系型或者分类单元,而不是传统意义上的AMF“种”。由于目前对于AMF种间和种内的基因差异水平尚不清楚[75],用于AMF分子进化分析的目标片段通常也无法在种的水平鉴别AMF[74],导致分子学研究中获得的AMF种系型/分类单元难以同传统意义上的AMF“种”建立明确的对应关系。以上问题的存在一定程度上限制了分子学方法的应用,阻碍了AMF物种多样性的研究进展。最近,运用分子学方法研究AMF的物种多样性取得了明显的进展。Krüger等[74]设计出4组AMF特异性的引物混合物,其目的片段所包含的变异位点具备在种的水平区分AMF的能力,被推荐作为AMF的DNA条形码(DNA barcoding)[76-77]。Wang等[20]首次报道了该组引物应用于野外AMF多样性调查的研究结果,发现除了其扩增片段具有足够的变异位点之外,该组引物具有很高的AMF特异性。如果在今后的研究当中能够进一步确认该序列作为DNA条形码的可行性,无疑能够极大的推进对AMF物种多样性的研究进展。
AMF物种多样性的研究进展在很大程度上依赖于研究手段的不断进步。近年来快速发展的新一代测序技术,如最新的454焦磷酸测序技术(454 pyrosequencing),在对AMF物种多样性的研究当中具备了传统分子学方法无可比拟的巨大优势。首先,新一代测序技术的测序深度在数量级上远远高于第一代测序技术,可以检测到极微量的AMF序列,便于发现寄主根内非优势的AMF种类。其次,新一代测序技术在对AMF特定的DNA序列进行PCR扩增之后直接进行测序,无需对目的片段进行克隆、分型分析等操作,其研究结果相对来讲更接近目标生境中AMF真实的群落结构。目前,国际上有研究已经报道了运用该技术对AMF多样性的调查结果。Öpik等[25]首次报道了运用该技术对一处自然生境中AMF多样性的调查结果,发现454焦磷酸测序技术可以检测到部分传统分子学方法无法检测到的AMF分类单元。Dumbrell等[31]和Gillevet等[78]也报道了类似的研究结果。Moora等[30]运用454测序技术调查了分布于不同地区棕榈树(Trachycarpus fortunei)根内的AMF群落结构;他们从14处棕榈树分布生境(包括4处自然分布的生境以及10处移栽的生境)的目标寄主根内一共检测到73个AMF分类单元。这些报道不仅充分证明了新一代测序技术在AMF物种多样性研究中的巨大应用前景,也在一定程度上支持了全球的AMF物种多样性水平被严重低估的观点。随着近年来454焦磷酸测序技术在序列可读长度(目前可达到700—1000 bp)以及成本控制方面的快速发展,可以预见在数年之内该技术将极大的加深人们对AMF物种多样性的理解。
4 展望
研究方法的改进与新技术的发展为目前AMF物种多样性的研究提供了极好的机遇;有理由相信,AMF物种多样性的研究将在几年之内取得较大的进展。除了新技术的发展值得期待之外,笔者认为在AMF物种多样性研究当中有以下两个方向是需要重点关注的。首先,加强对不同类型生境(尤其是各种特殊的生境类型)中AMF的物种多样性及其生态功能的研究,并深入探讨AMF的种类组成或多样性水平与它们在生态系统中所发挥的生态功能之间的联系。例如,尽管已经有明确的证据显示湿地生态系统中也广泛存在AMF与植物的共生[19-20,43],目前国内外对于各种类型湿地生态系统中AMF的物种多样性水平以及AMF在湿地生境中的生态功能还所知甚少。其次,结合常规研究方法以及最新的技术手段,通过构建大型永久样地或者开展大尺度(包括时间和空间跨度)调查试验的方式系统的研究AMF物种多样性对生境中其它土壤微生物、动物以及植物群落特征的影响机制,以深入理解AMF群落对整个生态系统(乃至全球生态系统)的影响及作用机制。
[1] Smith S E,Read D J.Mycorrhizal Symbiosis.Cambridge,UK:Academic Press,2008.
[2] Heckman D S,Geiser D M,Eidell B R,Kardos N L,Hedges S B.Molecular evidence for the early colonization of land by fungi and plants.Science,2001,293(5532):1129-1133.
[3] van der Heijden M G A,Bardgett R D,van Straalen N M.The unseen majority:soil microbes as drivers of plant diversity and productivity in terrestrial ecosystems.Ecology Letters,2008,11(3):296-310.
[4] Morton J B,Benny G L.Revised classification of arbuscular mycorrhizal fungi(Zygomycetes):a new order,Glomales,two new suborders,Glomineae and Gigasporineae,and two new families,Acaulosporaceae and Gigasporaceae with an emendation of Glomaceae.Mycotaxon,1990,37:471-491.
[5] Schüβler A,Schwarzott D,Walker C.A new fungal phylum,the Glomeromycota:Phylogeny and evolution.Mycological Research,2001,105(12):1413-1421.
[6] Stürmer S L.A history of the taxonomy and systematics of arbuscular mycorrhizal fungi belonging to the phylum Glomeromycota.Mycorrhiza,2012,22(4):247-258.
[7] Oehl F,da Silva G A,Goto B T,Sieverding E.Glomeromycota:three new genera and glomoid species reorganized.Mycotaxon,2011,116(1):75-120.
[8] Gai J P,Christie P,Feng G,Li X L.Twenty years of research on community composition and species distribution of arbuscular mycorrhizal fungi in China:a review.Mycorrhiza,2006,16(4):229-239.
[9] Husband R,Herre E A,Turner S L,Gallery R,Young J P W.Molecular diversity of arbuscular mycorrhizal fungi and patterns of host association over time and space in a tropical forest.Molecular Ecology,2002,11(12):2669-2678.
[10] Santos J C,Finlay R D,Tehler A.Molecular analysis of arbuscular mycorrhizal fungi colonizing a semi-natural grassland along a fertilization gradient.New Phytologist,2006,172(1):159-168.
[11] van der Gast C J,Gosling P,Tiwari B,Bending G D.Spatial scaling of arbuscular mycorrhizal fungal diversity is affected by farming practice.New Phytologist,2011,13(1):241-249.
[12] Cai X B,Peng Y L,Yang M N,Gai J P.Species diversity of arbuscular mycorrhizal fungi of Stipa L.in alpine grassland in northern Tibet in China.Acta Ecologica Sinica,2011,31(20):6029-6037.
[13] Liu Y J,He J X,Shi G X,An L Z,Öpik M,Feng H Y.Diverse communities of arbuscular mycorrhizal fungi inhabit sites with very high altitude in Tibet Plateau.FEMS Microbiology Ecology,2011,78(2):355-365.
[14] Börstler B,Renker C,Kahmen A,Buscot F.Species composition of arbuscular mycorrhizal fungi in two mountain meadows with differing management types and levels of plant biodiversity.Biology and Fertility of Soils,2006,42(4):286-298.
[15] Rodríguez-Echeverría S,Freitas H.Diversity of AMF associated with Ammophila arenaria ssp.arundinacea in Portuguese sand dunes.Mycorrhiza,2006,16(8):543-552.
[16] Li L F,Li T,Zhang Y,Zhao Z W.Molecular diversity of arbuscular mycorrhizal fungi and their distribution patterns related to host-plants and habitats in a hot and arid ecosystem,southwest China.FEMS Microbiology Ecology,2010,71(3):418-427.
[17] Wilde P,Manal A,Stodden M,Sieverding E,Hildebrandt U,Bothe H.Biodiversity of arbuscular mycorrhizal fungi in roots and soils of two salt marshes.Environmental Microbiology,2009,11(6):1548-1546.
[18] Zarei M,König S,Hempel S,Nekouei M K,Savaghebi G,Buscot F.Community structure of arbuscular mycorrhizal fungi associated to Veronica rechingeri at the Anguran zinc and lead mining region.Environmental Pollution,2008,156(3):1277-1283.
[19] Wang Y T,Qiu Q,Yang Z Y,Hu Z J,Tam N F Y,Xin G R.Arbuscular mycorrhizal fungi in two mangroves in South China.Plant and Soil,2010,331(1/2):181-191.
[20] Wang Y T,Huang Y L,Qiu Q,Xin G R,Yang Z Y,Shi S H.Flooding greatly affects the diversity of arbuscular mycorrhizal fungi communities in the roots of wetland plants.PLoS ONE,2011,6(9):e24512.
[21] Martínez-García L B,Armas C,Miranda J D,Padilla F M,Pugnaire F I.Shrubs influence arbuscular mycorrhizal fungi communities in a semi-arid environment.Soil Biology and Biochemistry,2011,43(3):682-689.
[22] BidartondoM I,Redecker D,Hijri I,Wiemken A,Bruns T D,Domínguez L,Sérsic A,Leake J R,Read D J.Epiparasitic plants specialized on arbuscular mycorrhizal fungi.Nature,2002,419(6905):389-392.
[23] Bever J D.Soil community feedback and the coexistence of competitors:conceptual frameworks and empirical tests.New Phytologist,2003,157(3):465-473.
[24] Klironomos J N.Variation in plant response to native and exotic arbuscular mycorrhizal fungi.Ecology,2003,84(9):2292-2301.
[25] Öpik M,Metsis M,Daniell T J,Zobel M,Moora M.Large-scale parallel 454 sequencing reveals host ecological group specificity of arbuscular mycorrhizal fungi in a boreonemoral forest.New Phytologist,2009,184(2):424-437.
[26] Öpik M,Moora M,Liira J,Zobel M.Composition of root-colonizing arbuscular mycorrhizal fungal communities in different ecosystems around the globe.Journal of Ecology,2006,94(4):778-790.
[27] Li L F,Li T,Zhao Z W.Differences of arbuscular mycorrhizal fungal diversity and community between a cultivated land,an old field,and a nevercultivated field in a hot and arid ecosystem of southwest China.Mycorrhiza,2007,17(8):655-665.
[28] Vandenkoornhuyse P,Husband R,Daniell T J,Watson I J,Duck J M,Fitter A H,Young J P W.Arbuscular mycorrhizal community composition associated with two plant species in a grassland ecosystem.Molecular Ecology,2002,11(8):1555-1564.
[29] Öpik M,Moora M,Zobel M,Saks Ü,Wheatley R,Wright F,Daniell T.High diversity of arbuscular mycorrhizal fungi in a boreal herb-rich coniferous forest.New Phytologist,2008,179(3):867-876.
[30] Moora M,Berger S,Davison J,Öpik M,Bommarco R,Bruelheide H,Kühn I,Kunin K E,Metsis M,Rortais A,Vanatoa A,Vanatoa E,Stout J C,Truusa M,Westphal C,Zobel M,Walther G.Alien plants associate with widespread generalist arbuscular mycorrhizal fungal taxa:evidence from a continental-scale study using massively parallel 454 sequencing.Journal of Biogeography,2011,38(7):1305-1317.
[31] Dumbrell A J,Ashton P D,Aziz N,Feng G,Nelson M,Dytham C,Fitter A H,Helgason T.Distinct seasonal assemblages of arbuscular mycorrhizal fungi revealed by massively parallel pyrosequencing.New Phytologist,2011,190(3):794-804.
[32] Bever J D,Schultz P A,Pringle A,Morton J B.Arbuscular mycorrhizal fungi:more diverse than meets the eye,and the ecological tale of why.Bioscience,2001,51(11):923-931.
[33] Liu R J,Wang F Y.Selection of appropriate host plants used in trap culture of arbuscular mycorrhizal fungi.Mycorrhiza,2003,13(3):123-127.
[34] Liu R J,Chen Y L.Mycorrhizology.Beijing:Science Press,2007.
[35] Guo S X,Liu R J.Effects of different peony cultivars on community structure of arbuscular mycorrhizal fungi in rhizosphere soil.Chinese Journal of Applied Ecology,2010,21(8):1993-1997.
[36] Hannula S E,de Boer W,van Veen J A.In situ dynamics of soil fungal communities under different genotypes of potato,including a genetically modified cultivar.Soil Biology and Biochemistry,2010,42(12):2211-2223.
[37] Burrows R,Pfleger F.Arbuscular mycorrhizal fungi respond to increasing plant diversity.Canadian Journal of Botany,2002,80(2):120-130.
[38] Liu R J,Jiao H,Li Y,Li M,Zhu X C.Research advances in species diversity of arbuscular mycorrhizal fungi.Chinese Journal of Applied Ecology,2009,20(9):2301-2307.
[39] Maherali H,Klironomos J N.Influence of phylogeny on fungal community assembly and ecosystem functioning.Science,2007,316(5832):1746-1748.
[40] Oehl F,Laczko E,Bogenrieder A,Stahr K,Bösch R,van der Heijden M,Sieverding E.Soil type and land use intensity determine the composition of arbuscular mycorrhizal fungal communities.Soil Biology and Biochemistry,2010,42(5):724-738.
[41] Antoninka A,Reich P B,Johnson N C.Seven years of carbon dioxide enrichment,nitrogen fertilization and plant diversity influence arbuscular mycorrhizal fungi in a grassland ecosystem.New Phytologist,2011,192(1):200-214.
[42] Gosling P,Ozaki A,Jones J,Turner M,Rayns F,Bending G D.Organic management of tilled agricultural soils results in a rapid increase in colonisation potential and spore populations of arbuscular mycorrhizal fungi.Agriculture,Ecosystems and Environment,2010,139(1/2):273-279.
[43] Wirsel S G R.Homogenous stands of a wetland grass harbour diverse consortia of arbuscular mycorrhizal fungi.FEMS Microbiology Ecology,2004,48(2):129-138.
[44] van Diepen L T A,Lilleskov E A,Pregitzer K S.Simulated nitrogen deposition affects community structure of arbuscular mycorrhizal fungi in northern hardwood forests.Molecular Ecology,2011,20(4):799-811.
[45] Zarei M,Hempel S,Wubet T,Schäfer T,Savaghebi G,Jouzani G S,Nekouei M K,Buscot F.Molecular diversity of arbuscular mycorrhizal fungi in relation to soil chemical properties and heavy metal contamination.Environmental Pollution,2010,158(8):2757-2765.
[46] Gai J P,Tian H,Yang F Y,Christie P,Li X L,Klironomos J N.Arbuscular mycorrhizal fungal diversity along a Tibetan elevation gradient.Pedobiologia-International Journal of Soil Biology,2012,55(3):145-151.
[47] Wu B Y,Hogetsu T,Isobe K,Ishii R.Community structure of arbuscular mycorrhizal fungi in a primary successional volcanic desert on the southeast slope of Mount Fuji.Mycorrhiza,2007,17(6):495-506.
[48] Adair C E,Reich P B,Hobbie S E,Knops J M H.Interactive effects of time,CO2,N and diversity on total belowground carbon allocation and ecosystem carbon storage in a grassland community.Ecosystems,2009,12(6):1037-1052.
[49] Zavalloni C,Vicca S,Büscher M,de la Providencia I E,de Boulois H D,Declerck S,Nijs I,Ceulemans R.Exposure to warming and CO2enrichment promotes greater above-ground biomass,nitrogen,phosphorus and arbuscular mycorrhizal colonization in newly established grasslands.Plant and Soil,2012,doi:10.1007/s11104-012-1190-y.
[50] Olsrud M,Carlsson B Å,Svensson B M,Michelsen A,Mellillo J M.Responses of fungal root colonization,plant cover and leaf nutrients to longterm exposure to elevated atmospheric CO2and warming in a subarctic birch forest understory.Global Change Biology,2010,16(6):1820-1829.
[51] Hawkes C V,Hartley I P,Ineson P,Itter A F.Soil temperature affects carbon allocation within arbuscular mycorrhizal networks and carbon transport from plant to fungus.Global Change Biology,2008,14(5):1181-1190.
[52] Emmett B A,Beier C,Estiarte M,Tietema A,Kristensen H L,Williams D,Penuelas J,Schmidt I,Sowerby A.The response of soil processes to climate change:results from manipulation studies of shrublands across an environmental gradient.Ecosystems,2004,7(6):625-637.
[53] Jonasson S,Castro J,Michelsen A.Litter,warming and plants affect respiration and allocation of soil microbial and plant C,N and P in arctic mesocosms.Soil Biology and Biochemistry,2004,36(7):1129-1139.
[54] Whitfield L,Richards A J,Rimmer D L.Relationships between soil heavy metal concentration and mycorrhizal colonisation in Thymus polytrichus in northern England.Mycorrhiza,2004,14(1):55-62.
[55] Helgason T,Daniell T J,Husband R,Fitter A H,Young J P W.Ploughing up the wood-wide web?Nature,1998,394(6692):431-431.
[56] Lin X G,Feng Y Z,Zhang H Y,Chen R R,Wang J H,Zhang J B,Chu H Y.Long-term balanced fertilization decreases arbuscular mycorrhizal fungal diversity in an arable soil in north China revealed by 454 pyrosequencing.Environmental Science and Technology,2012,46(11):5764-5771.
[57] Schnoor T K,Lekberg Y,Rosendahl S,Olsson P A.Mechanical soil disturbance as a determinant of arbuscular mycorrhizal fungal communities in semi-natural grassland.Mycorrhiza,2011,21(3):211-220.
[58] Daniell T L,Husband R,Fitter A H,Young J P W.Molecular diversity of arbuscular mycorrhizal fungi colonising arable crops.FEMS Microbiology Ecology,2001,36(2/3):203-209.
[59] Rosendahl S,Matzen H.Genetic structure of arbuscular mycorrhizal populations in fallow and cultivated soils.New Phytologist,2008,179(4):1154-1161.
[60] Hijri I,Sykorova Z,Oehl F,Ineichen K,Mäder P,Wiemken A,Redecker D.Communities of arbuscular mycorrhizal fungi in arable soils are not necessarily low in diversity.Molecular Ecology,2006,15(8):2277-2289.
[61] Vallino M,Massa N,Lumini E,Bianciotto V,Berta G,Bonfante P.Assessment of arbuscular mycorrhizal fungal diversity in roots of Solidago gigantea growing in a polluted soil in Northern Italy.Environmental Microbiology,2006,8(6):971-983.
[62] Mangan S A,Adler G H.Consumption of arbuscular mycorrhizal fungi by spiny rats(Proechimys semispinosus)in eight isolated populations.Journal of Tropical Ecology,1999,15(6):779-790.
[63] Roesti D,Ineichen K,Braissant O,Redecker D,Wiemken A,Aragno M.Bacteria associated with spores of the arbuscular mycorrhizal fungi Glomus geosporum and Glomus constrictum.Applied and Environmental Microbiology,2005,71(11):6673-6679.
[64] Tian H,Drijber R A,Niu X S,Zhang J L,Li X L.Spatio-temporal dynamics of an indigenous arbuscular mycorrhizal fungal community in an intensively managed maize agroecosystem in North China.Applied Soil Ecology,2011,47(3):141-152.
[65] Li Y P,He X L,Zhao L L.Tempo-spatial dynamics of arbuscular mycorrhizal fungi under clonal plant Psammochloa villosa Trin.Bor in Mu Us sandland.European Journal of Soil Biology,2010,46(5):295-301.
[66] Cuenca G,Lovera M.Seasonal variation and distribution at different soil depths of arbuscular mycorrhizal fungi spores in a tropical sclerophyllous shrubland.Botany,2010,88(1):54-64.
[67] Simon L,Lalonde M,Bruns T D.Specific amplification of 18S ribosomal genes from VA endomycorrhizal fungi colonizing roots.Applied and Environmental Microbiology,1992,58:291-295.
[68] Oliveira C A,Sá N M H,Gomes E A,Marriel I E,Scotti M R,Guimarães C T,Schaffert R E,Alves V M C.Assessment of the mycorrhizal community in the rhizosphere of maize(Zea mays L.)genotypes contrasting for phosphorus efficiency in the acid savannas of Brazil using denaturing gradient gel electrophoresis(DGGE).Applied Soil Ecology,2009,41(3):249-258.
[69] Kjøller R,Rosendahl S.Detection of arbuscular mycorrhizal fungi(Glomales)in roots by nested PCR and SSCP(Single Stranded Conformation Polymorphism).Plant and Soil,2000,226(2):189-196.
[70] Zarei M,König S,Hempel S,Nekouei M K,Savaghebi G,Buscot F.Community structure of arbuscular mycorrhizal fungi associated to Veronica rechingeri at the Anguran zinc and lead mining region.Environmental Pollution,2008,156(3):1277-1683.
[71] Mummey D L,Rillig M C.Evaluation of LSU rRNA-gene PCR primers for analysis of arbuscular mycorrhizal fungal communities via terminal restriction fragment length polymorphism analysis.Journal of Microbiological Methods,2007,70(1):200-204.
[72] Scheublin T R,Ridgway K P,Young J P W,van der Heijden M G A.Nonlegumes,legumes,and root nodules harbor different arbuscular mycorrhizal fungal communities.Applied and Environmental Microbiology,2004,70(10):6240-6246.
[73] Lee J,Lee S,Young J P W.Improved PCR primers for the detection and identification of arbuscular mycorrhizal fungi.FEMS Microbiology Ecology,2008,65(2):339-349.
[74] Krüger M,Stockinger H,Krüger C,Schüβler A.DNA-based species level detection of Glomeromycota:one PCR primer set for all arbuscular mycorrhizal fungi.New Phytologist,2009,183(1):212-223.
[75] Börstler B,Raab P A,Thiéry O,Morton J B,Redecker D.Genetic diversity of the arbuscular mycorrhizal fungus Glomus intraradices as determined by mitochondrial large subunit rRNA gene sequences is considerably higher than previously expected.New Phytologist,2008,180(2):452-465.
[76] Stockinger H,Krüger M,Schüβler A.DNA barcoding of arbuscular mycorrhizal fungi.New Phytologist,2010,187(2):461-476.
[77] Krüger M,Krüger C,Walker C,Stockinger H,Schüβler A.Phylogenetic reference data for systematics and phylotaxonomy of arbuscular mycorrhizal fungi from phylum to species level.New Phytologist,2012,193(4):970-984.
[78] Gillevet P M,Sikaroodi M,Torzilli A P.Analyzing salt-marsh fungal diversity:comparing ARISA fingerprinting with clone sequencing and pyrosequencing.Fungal Ecology,2009,2(4):160-167.
参考文献:
[12] 蔡晓布,彭岳林,杨敏娜,盖京苹.藏北高寒草原针茅属植物AM真菌的物种多样性.生态学报,2011,31(20):6029-6037.
[34] 刘润进,陈应龙.菌根学.北京:科学出版社,2007.
[35] 郭绍霞,刘润进.不同品种牡丹对丛枝菌根真菌群落结构的影响.应用生态学报,2010,21(8):1993-1997.
[38] 刘润进,焦惠,李岩,李敏,朱新产.丛枝菌根真菌物种多样性研究进展.应用生态学报,2009,20(9):2301-2307.
