青藏高原高寒草原区工程迹地面积对其恢复植物群落特征的影响
2013-12-16郭正刚
毛 亮,周 杰,郭正刚
(兰州大学草地农业科技学院草地农业生态系统国家重点实验室,兰州 730020)
高寒草原是青藏高原草地的重要组分[1],也是我国重要的高山植物基因库和碳汇载体[2],更是流域生态安全屏障和当地畜牧业生产的物质基础[3]。青藏公路和铁路,高压电线和光缆等人类线性工程穿越青藏高原高寒高原时,总会在施工地段或附近地区形成面积大小不同的工程迹地[4],因此工程迹地植被恢复是目前青藏高原受损高寒草原生态系统修复的重要内容。西方国家工程迹地恢复最早始于20世纪40年代的筑路迹地恢复[5],而我国筑路迹地恢复的研究始于高速公路护坡[6],21世纪初期开始逐渐增多。筑路迹地恢复包括人工恢复和自然恢复两种。由于我国筑路迹地恢复始于高速公路建设,因此筑路迹地恢复目前以人工恢复研究较多[4,6-7]。虽然筑路迹地自然恢复是植被恢复的一种主要方式,且具有省时、省力和省财,避免草皮移植对客地原生草原影响等优点,但目前研究相对较少。马世震等[5]通过对比不同恢复时期植物群落及多样性特征,指出青藏高原高寒草原区工程迹地植被恢复至少需要20年;祝广华等[6]通过比较不同草地类型经历等同恢复时间的特征,认为高寒草原和高寒草甸是受工程活动影响最广的草地类型;Guo等[3]通过分析未干扰群落与离工程迹地不同距离处恢复群落的特征,明晰筑路工程对高寒草原的干扰尺度在离路基100 m的范围内。然而目前关于工程迹地自然恢复研究均集中于工程迹地上已经恢复的植物群落,忽略经历相同时期因面积较大至今仍然没有植被恢复的工程迹地,这说明工程迹地面积影响着工程迹地植物群落的自然恢复进程,但工程迹地面积变化与恢复植物群落的关系多限于直观的感受,缺乏科学试验提供佐证。
人类工程迹地主要包括路基周边,取弃土场,沙石料场,施工便道,营地场地及施工地等[7],其不同的受损程度决定了植被恢复进程存在着差异。青藏公路路基主体显著高于周边取土迹地,路基周边土壤因汇集了路面降水而持有较高的含水量,而且容易滞留风媒种子和花粉[8],从斑块恢复学上,路基周边迹地拥有最好的植物群落自然恢复物质基础,从干扰程度上,路基周边植被破坏严重(筑路一般就近取土),分布范围广,具有一定的代表性和典型性[3,7]。本研究以青藏高原平坦地区青藏公路的路基周边迹地为例,通过调查经历相同恢复时期而面积不同工程迹地上的植物群落特征,明确工程迹地面积对青藏高原高寒草原区植被恢复进程的影响。
1 材料与方法
1.1 研究区概况
研究地区位于青藏公路沿线沱沱河北岸,距沱沱河20 km,(即国道109线3144号路基桩和3126号路基桩之间),地理坐标为92°37′E,34°22′N,海拔4550—4560 m。气候属高原大陆性气候,全年冰冻期 331 d,无霜期9—50 d,平均气温-4.2℃,最低气温-14.8℃,最高气温6.7℃,年均降水量275.92 mm,夏季降水占全年降雨70%[9]。土壤以高寒草原土为主。未受工程干扰的草原为青藏苔草(Carex moorcroftii)紫花针茅(Stipa purpurea)草原,优势种为紫花针茅和青藏苔草,主要伴生种有草地早熟禾(Poa pratensis)、短穗兔耳草(Lagotis brachystachya)和矮火绒草(Leontopodium nanum)等[10]。
1.2 研究方法
1.2.1 样地设置与调查
在1994年扩建和修缮青藏公路时遗留的路基周边迹地上布设样地。为了选择研究样地,首先实测56个单独连片的工程迹地,记录各个工程迹地恢复植物群落的高度、盖度,观察紫花针茅是否出现。筑路迹地形状多为狭长型,中间为长方形,两边为半圆形,在测定其面积时,采用分割法测定求和。然后以恢复植物群落的高度、盖度、紫花针茅出现否作为变量,对56个工程迹地聚类,聚类结果表明,56个工程迹地可划分为4个类群,4个类群工程迹地面积的平均值分别为55,156,254,583 m2。根据4个类群工程迹地面积平均值设置样地,工程迹地面积梯度分别为(55±10)m2,(156±20)m2,(254±30)m2,(583±40)m2,形成4 个面积梯度处理,然后每个梯度选择3个样地作为研究对象(3次重复),共计12个,12个样地的周长和面积基本接近(表1)。同时在工程迹地邻近、未受工程干扰的青藏苔草紫花针茅草原布设3个10 m×10 m的对照样地。共计15个样地,各个样地的海拔、坡向、地表基况基本保持一致。在每个样地对角线上布设4个1 m×1 m的样方,分别统计样方内的物种组成、群落盖度、高度、频度,然后收获期地上生物量,带回实验室烘干称重,共计调查样方60个。野外调查于2011年7—8月进行。

表1 12个筑路工程迹地样地的周边和面积Table 1 Total circumferences and areas of 12 plots from land used for engineering construction
1.2.2 α多样性的测度
分别选用Shannon-Wiener指数、Pielou指数、Simpson指数、样方内出现的物种数,测度多样性指数(H)、均匀度指数(J)、优势度指数(D)和丰富度指数(S)[11],其计算公式分别为:

式中,Pi为种i的重要值,即Pi=(相对盖度+相对密度+相对频度)/3,S以样方内出现的物种数量度量。
β多样性选用Whittaker指数(βws)测度[3],其计算公式分别为:
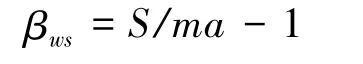
式中,S为系统的总物种数;ma为样方的平均物种数。
1.2.3 植被恢复能力评价
评价工程迹地与自然地植被恢复能力的综合指标用SL[12-13],计算公式为:

式中,H代表工程迹地Shannon-Wiener指数;H0代表未干扰地Shannon-Wiener指数;ΔF指相同地点工程迹地与未干扰草地植被覆盖度差值;I是顶级物种侵入指数,计算方法为:

式中,Pi表示原优势建群物种出现的频度(即优势建群物种的重要值)。
1.3 数据处理
采用SPSS 17.0软件进行ANOVA方差分析,Excel制图。
2 结果
2.1 工程迹地面积对恢复植物群落组分的影响
工程迹地上恢复植物群落的优势种和主要伴生种随着工程迹地面积变化而发生明显变化(表2)。工程迹地面积为583 m2时,群落优势种为短穗兔耳草,虽然青藏苔草和草地早熟禾等草原植物成功定居,但没有成为群落优势种群。工程迹地面积为254 m2时,优势种被青藏苔草替代,而短兔耳草沦落为伴生种,此时定居的伴生种还有矮火绒草和草地早熟禾等。工程迹地面积缩至156 m2时,群落优势种演变为青藏苔草和草地早熟禾,伴生种出现沙生凤毛菊(Saussurea arenaria)、羽叶点地梅(Pomatosace filicula)和紫花针茅。工程迹地面积缩小到55 m2,优势种除了青藏苔草以外,还有矮火绒草,此时高寒草原指示植物紫花针茅作为首要伴生种出现。未受筑路工程干扰的高寒草原主要以紫花针茅和青藏苔草为优势种,伴生种种类丰富。这说明随着工程迹地面积的增加,植物群落恢复速率越慢,环境旱生化,恢复植物群落组分结构越单一,草原指示物种出现越困难,而工程迹地面积越小,恢复植物群落越接近顶级群落,在工程迹地156 m2内出现了高寒草原指示物种紫花针茅,然后随着工程迹地面积缩小,群落结构趋向复杂。

表2 青藏高原高寒草原区不同工程迹地面积上恢复植物群落的组分Table 2 Component of restorable plant community at different land areas used for engineering construction in the alpine steppe of the Qinghai-Tibetan Plateau
2.2 工程迹地面积对恢复植物群落空间结构的影响
2.2.1 群落高度
青藏高原高寒草原区,恢复植物群落的高度随着工程迹地面积减小而逐渐增加(图1)。当工程迹地面积小于或等于156 m2时,虽然恢复群落间或者恢复群落与未干扰群落间的高度有所差异,但并不显著,但其显著大于工程迹地面积为583 m2时恢复植物群落的高度(P<0.05)。工程迹地面积为55、156、254、583 m2时恢复植物群落的高度分别为未干扰群落高度的65%、69%、73%和83%,反映了工程迹地面积越小,恢复植物群落高度约接近于未干扰群落的高度。
2.2.2 群落总盖度

图1 青藏高原高寒草原区不同工程迹地面积上恢复植物群落高度Fig.1 Height of restorable plant community at different land areas used for engineering construction in the alpine steppe of the Qinghai-Tibetan Plateau

图2 青藏高原高寒草原区不同面积工程迹地上恢复群落总盖度Fig.2 Covers of restorable plant community at different land areas used for engineering construction in the alpine steppe of the Qinghai-Tibetan Plateau
随着工程迹地面积增加,恢复群落总盖度逐渐减少(图2),当工程迹地面积为55 m2,恢复群落盖度和未干扰群落盖度没有显著差异,而他们显著大于工程迹地面积大于或等于156 m2时恢复植物群落的总盖度(P<0.05),工程迹地面积为55、156、254、583 m2时恢复植物群落的总盖度分别为未干扰群落盖度的47%、62%、65%和87%,说明工程迹地面积越小,植物群落盖度越易恢复,反之亦然。
2.3 工程迹地面积对植物多样性的影响
2.3.1 α多样性
随着工程迹地面积减小,植物物种多样性指数和丰富度指数均逐渐增大(表3),其中多样性指数在工程迹地面积55 m2时与未干扰草地没有显著差异,但当工程迹地面积大于156 m2,其恢复群落的多样性指数显著小于未干扰草地的多样性指数(P<0.05),说明工程迹地面积为156 m2,工程迹地恢复群落的物种多样性指数已接近于未干扰地群落的多样性指数。植物物种丰富指数和多样性指数随着工程迹地面积变化过程基本趋同。随着工程迹地面积减小,植物均匀度指数变化较为复杂,其中均匀度指数的最大值和最小值分别出现在工程迹地面积最大的样地和未干扰地,在恢复植物群落序列中,均匀度指数在程迹地面积为254 m2时最小,此时均匀度指数和未干扰群落均匀度指数差异不显著。优势度指数随着工程迹地面积变化虽然存在分异,但其在恢复群落之间,以及恢复群落和未干扰群落间均差异不显著,说明工程迹地面积变化对优势度指数没有显著的影响。

表3 青藏高原高寒草原区不同面积工程迹地上恢复植物群落的丰富度指数、均匀度指数、多样性指数和优势度指数Table 3 Richness index,evenness index,diversity index and dominant index of restorable plant community at different land areas used for engineering construction in the alpine steppe of the Qinghai-Tibetan Plateau
2.3.2 β多样性
植物群落β多样性指数随工程迹地面积减小而逐渐减小(图3),工程迹地面积为583 m2时群落的β多样性指数显著大于工程迹地面积156 m2和254 m2的β多样性指数(P<0.05),而后者显著大于工程迹地面积为55 m2时的恢复群落的β多样性指数(P<0.05),所有恢复群落的β多样性指数显著大于未干扰群落的β多样性指数(P<0.05),说明工程迹地面积越小,环境异质性越小,群落内物种替代速率降低。

图3 青藏高原高寒草原区不同面积工程迹地恢复群落的β多样性指数Fig.3 β diversity index of restorable plant community at different land areas used for engineering construction in the alpine steppe of the Qinghai-Tibetan Plateau
2.4 工程迹地面积对植物地上生物量的影响
地上生物量是植物地上生产能力的综合测度指标,更是退化草原恢复生产流程的物质基础。研究结果表明,经过19a的恢复,工程迹地上植物地上生物量从无到有,但增加量随着工程迹地面积变化而明显不同(图4),当工程迹地面积小于或等于156 m2时,其地上生物量和未干扰群落的生物量差异不显著,说明已经基本恢复到未干扰的水平,而当工程面积大于或等于254 m2后,经历相同时期恢复群落的生物量仍然显著小于未干扰群落的生物量(P<0.05),工程迹地面积为583 m2和254 m2时恢复植物群落的生物量仅为未干扰群落的48%和49%,说明植物群落生物量的恢复存在一个工程迹地面积的临界值。

图4 青藏高原高寒草原不同面积工程迹地上的生物量Fig.4 Biomass of restorable plant community at different land areas used for engineering construction in the alpine steppe of the Qinghai-Tibetan Plateau
2.6 工程迹地面积对群落恢复度的影响
群落恢复度是反应干扰迹地上植被恢复能力的综合指标,综合了植物多样性,初级生产力和群落内种群分布特征的信息。试验结果表明,经历相同恢复时期,不同工程迹地面积植被恢复能力存在明显差异(图5),表现为随工程迹地面积增加,SL值逐渐显著减小(P<0.05),说明植物恢复能力越弱。

图5 青藏高原高寒草原区不同面积工程迹地植被恢复能力Fig.5 Plant Restoration capability at different land areas used for engineering construction in the alpine steppe of the Qinghai-Tibetan Plateau
3 讨论
3.1 工程迹地面积变化对高寒草原区恢复植物群落组分、结构和生产力的影响
受损草地生态系统在自然恢复过程中,通过逐渐优化和改善群落组分和结构,正向演替,这种过程贯穿植被恢复演替进程[14-15]。工程迹地产生过程具有突发特性,其植被恢复过程较不同于逐渐退化草地的恢复过程[3],这是因为工程迹地土壤内植物繁殖体较逐渐退化草地要少的多,而植物繁殖体萌发后是否健康生长,与工程迹地面积大小密切相关。本研究结果表明,经历相同恢复时期,不同工程迹地面积上恢复植物群落的组分,盖度,高度和生物量均存在明显差异,表现为工程迹地面积越小,恢复植物群落的组分、空间结构和生产力指标越接近于未干扰群落的指标,说明其恢复较好,而面积越大,恢复程度越慢,这是因为工程迹地面积较小时,生境相对温和,此时土壤内残存成熟植物种子或其它繁殖构件,或者其他入侵物种的繁殖体,在适宜环境下萌发或繁殖,新生植物根系容易获取土壤养分和水分,容易存活,正向演替,经过一定时期后逐渐形成稳定植物群落[16],而工程迹地面积较大时,很多植物繁殖体萌发后,由于裸斑面积太大,土壤蒸发大,特别是土壤浅层含水量较低,新生植物因不易获取土壤养分和水分,在严酷环境中很快死亡,只有抗逆性较强的植物能够残活[3,17],但因其环境恶劣,生长过程较其在面积较小工程迹地上的过程要慢,这客观上减缓了面积较大工程迹地上植物群落的自然恢复过程。当工程迹地面积小于156 m2时,群落优势种由过渡性植物向原有群落演替,大量属于未干扰草原群落的植物成分涌现,草原指示植物紫花针茅已零星出现,当工程迹地面积继续减少到55 m2时,草原指示物种紫花针茅已成为主要伴生种,群落组分多样化,伴生种数量、优势种高度和重要值,群落高度和盖度均明显增加,说明工程迹地面积小于156 m2时,高寒草原受到干扰后具有较强的自我恢复能力[18]。而当工程迹地面积超过254 m2时,只有能够存活于极端耐严酷条件的青藏苔草出现,而其余物种均为杂草,群落组分简单,伴生种种类较少,群落盖度和高度相对较低,优势种高度较低,说明工程迹地面积超过254 m2时,工程迹地植物群落恢复能力较弱。工程迹地面积较大时,植物繁殖体定居及其种群扩张需要的时期可能更长,后者土壤水分和养分条件无法支持新定居种群的需要,实现恢复相对困难。采用综合反映群落盖度、顶级群落物种出现数与频度的植被恢复能力综合评价指标评价的结果再次印证了这个结果,从SL的分异性可以看出,工程迹地面积越小,其值越大,而工程迹地面积越小,其值越大,充分反映了青藏高原高寒草原区,面积较小的工程迹地较面积较大的工程迹地在经历相同恢复期后,更容易形成稳定的群落组分体系[12],逐渐趋向于未受干扰的群落。
3.2 工程迹地面积变化对高寒草原区恢复群落植物多样性的影响
植物多样性是高寒草原维持稳产和生态系统健康的关键所在[3,11],工程迹地恢复群落的植物多样性变化反应了其演替趋向。本研究结果表明,工程迹地面积对恢复群落的植物多样性具有明显影响,多样性指数和丰富度指数随着工程迹地面积减小而逐渐增大,其临界发生在工程迹地面积为254 m2时,当工程迹地面积小于或等于此值时,虽然物种丰富度指数仍然小于未干扰群落,但已经达到未干扰群落的67%以上,面积越小,越接近于未干扰群落的物种丰富度,而多样性指数基本恢复至未干扰群落的水平,群落内种群分布相对均匀[19],主要是工程迹地面积在一定的范围内,小环境一致性增大,伴生种在较一致的环境中能够很好的生长,各个种出现机率差异小。当工程迹地面积大于254 m2时,植物繁殖体因入侵途径延长而变得困难[20],部分新生物种因无法适应土壤水分蒸发快的环境而死亡,只有能够忍耐极端环境的短穗兔耳草、青藏苔草和矮火绒草等植物生长[11],导致群落样方内物种数很少,仅为未干扰群落的30%—40%。
物种多样性反映了群落内部物种数量的变化,而β多样性则反映的是群落间多样性在质方面的分异,其反映了不同群落间共有物种数的多少,揭示群落间物种的替代速率[21],两个群落间共有物种数少,说明其分布地环境差异大[22]。β多样性指数对工程迹地面积变化的响应结果表明,工程迹地面积越小,β多样性指数逐渐减少,说明群落内物种替代速率越低,其和未干扰群落共有的物种数增加,工程迹地的环境条件越趋向于未干扰群落的环境[23],反之亦然。
恢复群落的α多样性和β多样性对工程迹地面积变化的响应过程说明,工程迹地恢复群落的植物多样性恢复过程受控于面积,且面积的拐点出现在254 m2,当工程迹地面积大于此值时,植物多样性恢复较慢,群落间物种替代速率高[24],其生境和未干扰群落的生境差异大[3],而当工程迹地面积小于此值时,经过19年的恢复,工程迹地α多样性已基本接近未受干扰的草原群落,恢复群落和未受干扰群落间共有物种数逐渐增多,生境异质性减弱[15],说明青藏高原高寒草原区工程迹地植物多样性的恢复不仅受时间的调控,而且受工程迹地面积大小的调控,当工程迹地面积小于254 m2时,经历一定时期工程迹地上植物多样性可以恢复,而当工程迹地面积大于254 m2时,植物多样性恢复的时间可能更长或不易恢复。
致谢:兰州大学草地农业科技学院的藕洋、冯成庸、王丹丹、张晓黎、赵旭同学参加野外调查工作,特此致谢。
[1] Ren Z H,Zhang Y G,Li D Q,Xiao Q M,Cai C Y.The soil microbial activities and microbial biomass in Sanjiangyuan alpine grassland.Acta Ecologica Sinica,2011,31(11):3232-3238.
[2] Han D R,Cao G M,Guo X W,Zhang F W,Li Y K,Lin L,Li J,Tang Y H,Gu S.The potential of carbon sink in alpine meadow ecosystem on the Qinghai-Tibetan Plateau.Acta Ecologica Sinica,2011,31(24):7408-7417.
[3] Guo Z G,Long R J,Niu F J,Wu Q B,Hu Y K.Effect of highway construction on plant diversity of grassland communities in the permafrost regions of the Qinghai-Tibet plateau.The Rangeland Journal,2007,29(2):161-167.
[4] Zhou G Y,Li T C,Xu W H,Sun J,Ma H,Chen G C.Characteristics analysis of mineral elements content in dominant species and soil of artificial restoration vegetation of Qinghai-Tibetan Plateau permafrost region.Resources and Environment in the Yangtze Basin,2011,20(2):191-196.
[5] Spellerberg I F.Ecological Effects of Roads.Plymouth,UK:Science Publishers Inc,2002.
[6] Liu C X,Han L B.Review of researches in vegetation restoration of freeway slopes.Acta Ecologica Sinica,2007,27(5):2090-2098.
[7] Cheng P,Ji W S.Research on the test of vegetation recover in summit area of Tanggula Mountain along Qinghai-Tibet railway.Journal of Railway Engineering Society,2008,25(1):48-52.
[8] Ma S Z,Chen G C,Peng M,Zhou G Y,Zhao Y L.The alpine steppe vegetation restoration process of fountainhead region of Yangtze River.China Environmental Science,2004,24(2):188-191.
[9] Zhu G H,Tao L,Ren J.Evaluation of using land for constructing Qinghai-Tibet railway on native vegetation.Acta Agrestia Sinica,2006,14(2):160-164.
[10] Yang S Z,Jin H J,Ji Y J,Wei Z,He R X.Revegetation in permafrost regions along a linear project.Journal of Glaciology and Geocryology,2008,30(5):875-882.
[11] Norris J E.Root reinforcement by hawthorn and oak roots on a highway cut-slope in Southern England.Plant and Soil,2005,278(12):43-53.
[12] Zhang G S,Shi X H,Li D l,Wang Q C,Dai S.Climate change in Tuotuohe area at the headwaters of Yangtze River.Journal of Glaciology and Geocryology,2006,28(5):678-685.
[13] Liu Y,Hu G,Liang S C,Peng M,Lu X F,Yue P P.Interspecific associations analysis of Stipa purpurea community in Tuotuo River area.Guihaia,2007,27(5):720-724.
[14] Guo Z G,Li X F,Liu X Y,Zhou X R.Response of alpine meadow communities to burrow density changes of plateau pika(Ochotona curzoniae)in the Qinghai-Tibet Plateau.Acta Ecologica Sinica,2012,32(1):44-49.
[15] Wang G X,Yao J Z,Guo Z G,Wu Q B,Wang Y B.Changes in permafrost ecosystem under the influences of human engineering activities and its enlightenment to railway construction.Chinese Science Bulletin,2004,49(16):1741-1750.
[16] Li D M,Guo Z G,An L Z.Assessment on vegetation restoration capacity of several grassland ecosystems under destroyed disturbance in permafrost regions of Qinghai-Tibet Plateau.Chinese Journal of Applied Ecology,2008,19(10):2182-2188.
[17] Huang Z L,Kun G H,He D Q.Plant community diversity in Dinghushan Nature Reserve.Acta Ecologica Sinica,2000,20(2):193-198.
[18] Hou Y,Guo Z G,Long R J.Changes of plant community structure and species diversity in degradation process of Shouqu wetland of Yellow River.Chinese Journal of Applied Ecology,2009,20(1):27-32.
[19] Han L H,Shang Z H,Ren G H,Wang Y L,Ma Y S,Li X L,Long R J.The response of plants and soil on black soil patch of the Qinghai-Tibetan Plateau to variation of bare-patch areas.Acta Prataculturae Sinica,2011,20(1):1-6.
[20] Wheeler A P,Angermeier P L,Rosenberger A E.Impacts of new highways and subsequent landscape urbanization on stream habitat and biota.Reviews in Fisheries Science,2005,13(3):141-164.
[21] Trombulak S C,Frissel C A.Review of ecological effects of roads on terrestrial and aquatic communities.Conservation Biology,2000,14(1):18-30.
[22] Guo Z G,Niu F J,Zhan H,Wu Q B.Changes of grassland ecosystem due to degradation of permafrost frozen soil in the Qinghai-Tibet Plateau.Acta Ecologica Sinica,2007,27(8):3294-3301.
[23] Katopodis C,Aadland L P.Effective dam removal and river channel restoration approaches.International Journal of River Basin Management,2006,4(3):153-168.
[24] Guo Z G,Liu H X,Wang G X,Cheng G D.Effect of the Qinghai-Tibetan Highway on the β diversity of grassland plant communities in the northern region of the Qinghai-Tibetan Plateau.Acta Ecologica Sinica,2004,24(2):384-388.
参考文献:
[1] 任佐华,张于光,李迪强,肖启明,蔡重阳.三江源地区高寒草原土壤微生物活性和微生物量.生态学报,2011,31(11):3232-3238.
[2] 韩道瑞,曹广民,郭小伟,张法伟,李以康,林丽,李婧,唐艳鸿,古松.青藏高原高寒草甸生态系统碳增汇潜力.生态学报,2011,31(24):7408-7417.
[4] 周国英,李天才,徐文华,孙菁,马海,陈桂琛.多年冻土区工程迹地人工恢复区植物和土壤的矿质元素含量特征.长江流域资源与环境,2011,20(2):191-196.
[6] 刘春霞,韩烈保.高速公路边坡植被恢复研究进展.生态学报,2007,27(5):2090-2098.
[7] 陈鹏,季维生.青藏铁路唐古拉山越岭地段植被恢复试验研究.铁道工程学报,2008,25(1):48-52.
[8] 马世震,陈桂琛,彭敏,周国英,赵以莲.青藏公路取土场高寒草原植被的恢复进程.中国环境科学,2004,24(2):188-191.
[9] 祝广华,陶玲,任珺.青藏铁路工程迹地对植被的影响评价.草地学报,2006,14(2):160-164.
[10] 杨思忠,金会军,吉延峻,魏智,何瑞霞.寒区线性工程沿线冻土区的植被恢复.冰川冻土,2008,30(5):875-882.
[12] 张国胜,时兴合,李栋梁,汪青春,戴升.长江源沱沱河区45a来的气候变化特征.冰川冻土,2006,28(5):678-685.
[13] 刘洋,胡刚,梁士楚,彭敏,卢学峰,岳鹏鹏.沱沱河地区紫花针茅群落种间联结性分析.广西植物,2007,27(5):720-724.
[16] 李东明,郭正刚,安黎哲.青藏高原多年冻土区不同草地生态系统恢复能力评价.应用生态学报,2008,19(10):2182-2188.
[17] 黄忠良,孔国辉,何道泉.鼎湖山植物群落多样性的研究.生态学报,2000,20(2):193-198.
[18] 后源,郭正刚,龙瑞军.黄河首曲湿地退化过程中植物群落组分及物种多样性的变化.应用生态学报,2009,20(1):27-32.
[19] 韩立辉,尚占环,任国华,王彦龙,马玉寿,李希来,龙瑞军.青藏高原“黑土滩”退化草地植物和土壤对秃斑面积变化的响应.草业学报,2011,20(1):1-6.
[22] 郭正刚,牛富俊,湛虎,吴青柏.青藏高原北部多年冻土退化过程中生态系统的变化特征.生态学报,2007,27(8):3294-3301.
[24] 郭正刚,刘慧霞,王根绪,程国栋.人类工程对青藏高原北部草地群落β多样性的影响.生态学报,2004,24(2):384-388.
