松果梢斑螟对虫害诱导寄主防御的抑制作用
2013-12-16李秀玲李新岗杨立军
张 晓,李秀玲,李新岗,杨立军,陈 辉
(西北农林科技大学林学院/国家林业局黄土高原重点实验室,杨凌 712100)
松果梢斑螟(Dioryctria pryeri Ragonot)和油松球果小卷蛾(Gravitarmata margarotana(Heinemann))是我国松属针叶树重要的球果和枝梢害虫,一年一代,以幼虫钻蛀危害,长期以来对我国针叶树的正常生长、自然更新和种子生产造成很大影响[1]。特别是北方油松(Pinus tabuleaformis Mill.)良种基地,受害更为严重[2-3]。由于球果小卷蛾对油松球果的先期危害,引诱松果梢斑螟趋向虫害球果产卵和取食,即诱导负防御[3-5]或诱导敏感性[6]。小卷蛾幼虫危害时间比梢斑螟幼虫早7—10 d,并且小卷蛾幼虫的先期危害诱导了油松球果的直接防御和间接防御[2-4];如显著增加了蛋白酶抑制剂(PIs)、多酚氧化酶(PPO)和松脂含量,特别是双萜松脂酸[2-3]。梢斑螟幼虫在油松上,发育快、个体较大、活跃、竞争能力强,可在虫害球果中正常发育[1]。但高含量松脂(松脂酸)球果(如华山松球果)严重影响梢斑螟幼虫的生长与存活,松脂酸也成为梢斑螟幼虫的直接防御物质[1,3,5]。
近年来,关于虫害诱导负防御的研究已经很多,主要是引诱成虫产卵或幼虫取食的报道,并集中在虫害诱导挥发物的引诱方面[6-9]。虫害诱导负防御是植物诱导防御的一种形式[5-6],不但松果梢斑螟趋向虫害球果,梢斑螟属(Dioryctria)和其它类群的昆虫也都有趋向虫害寄主或组织的习性[6-8]。但趋向虫害寄主取食的害虫,如何克服或适应虫害的诱导防御,可能涉及一些科学问题,如转基因抗虫植物被更严重的害虫适应或克服、诱抗剂应用后引起潜在害虫的严重危害等。Karban和Baldwin认为,虫害寄主(诱导挥发物)暗示着竞争者和天敌的存在、植物营养质量的下降,也代表着潜在寄主的存在[6]。
害虫取食危害与其激发子一起,激活植物防御基因,使植物产生诱导防御物质对付昆虫。但葡萄糖氧化酶(glucose oxidase,GOX)是多食性或具强竞争性鳞翅目幼虫下唇腺中抑制植物诱导防御的“激发子”,可抑制植物的防御,提高害虫自身的适应性[6,10-18]。采用分光光度计法可测定昆虫下唇腺GOX活性[18-20],且同一昆虫不同寄主组织或不同质量的饲料对昆虫GOX活性影响很大[14,18-19]。据生长率理论,昆虫的生长率和虫体RNA含量与P含量高度耦合[21],测定同一龄期不同组织梢斑螟幼虫RNA和P含量[21-22],就可反映取食组织对梢斑螟幼虫生长发育的影响。为了进一步探讨小卷蛾危害引起油松球果的负防御机制,以及梢斑螟幼虫对寄主防御的适应和抑制作用[3],本文研究了不同组织梢斑螟幼虫GOX活性和虫害组织中的萜类防御物质变化,以及不同组织中梢斑螟幼虫的RNA和P含量。
1 材料与方法
1.1 供试材料
1.1.1 材料来源
油松球果和新梢采自甘肃省正宁县中湾油松良种基地,位于子午岭西部,海拔1300—1500 m,1983—1985 年营建,树龄30a,树高约6 m,405 株/hm2,郁闭度0.5—0.6,坡度10—20°。采样时间为2009 和2010 年5月,正是松果梢斑螟幼虫的危害期,通过田间接虫标记,并选择虫害第1天、第5天、第10天和第15天(分别为3、4、5龄和老熟幼虫)的球果和新梢,以健康球果和新梢为对照,每个处理3个重复。球果的取样部位为2年生球果的鳞片,分别从蛀孔左侧、右侧及后方取样;新梢的取样部位为当年生新梢顶部,约2 cm,三分法取样。球果、新稍取样均为 0.3g,置于-80℃用于防御物质的分析[3,23]。
1.1.2 供试昆虫
松果梢斑螟幼虫采样地点和时间同上。采集正在取食球果和新梢的梢斑螟幼虫,每两天采样1次。按取食部位,将其分为危害球果的1果1虫、1果多虫、与小卷蛾共生和危害新梢4个处理。每个处理分别取3龄、4龄和5龄取食活跃期和脱皮期幼虫,以及老熟幼虫各30—60头,共3份材料,置于-80℃分别用于测试幼虫下唇腺GOX活性、RNA和P含量。
1.2 梢斑螟幼虫葡萄糖氧化酶活性测定
1.2.1 幼虫下唇腺提取液的制备
分别解剖1果1虫、1果多虫、与小卷蛾共生和危害新梢4个处理的梢斑螟幼虫,取出下唇腺,在pH 7.0的磷酸缓冲液(0.05 mol/L)中冰浴研磨,4℃ 12000×g离心15 min,取上清液为下唇腺提取液。3龄幼虫每样品中有10头幼虫的下唇腺,其他龄期每样品中有5头幼虫下唇腺,重复6次[13,19]。
1.2.2 下唇腺葡萄糖氧化酶活性测定
GOX活性测定参照Bergmeyer等方法[24],用UV-1240(uv mini)分光光度计测定。35℃条件下,反应混合物包括:0.17 mmol/L 邻联茴香胺(Sigma),1.72%D-葡萄糖缓冲溶液(pH 7.0)2.9 mL,60 U/mL 辣根过氧化物酶(Roche)溶液和下唇腺提取液各0.1mL。用0.1 mL缓冲液代替下唇腺提取液为对照。在500 nm下连续记录5 min反应混合物的光吸收,计算每分钟的光吸收变化并作图,取线性变化范围内每分钟变化值。蛋白质浓度测定以牛血清蛋白为标准蛋白,用考马斯亮蓝染色法测定[19,25]。
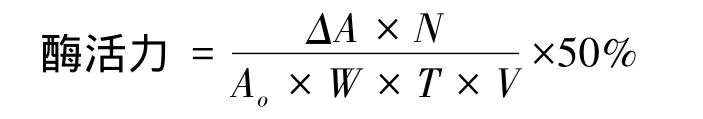
式中,△A表示线性变化范围内每分钟变化值;N表示酶液总体积;W表示蛋白质含量(mg/mL);T表示光反应时间(min);V表示加入的酶液体积。Ao表示邻联茴香胺在500 nm下的摩尔消光系数,为7.5。
1.3 寄主萜类化合物的提取与分析
1.3.1 萜类成分的提取
根据Miller等的叔丁基甲基醚(TBME)浸提并甲酯化法(简称甲酯化法),选取0.3g球果或新梢样品,加入1.5 mL叔丁基甲基醚(加150 μg/mL异丁基苯为内标),震荡浸提14 h,过滤,用0.3 mL 0.1 mol/L的(NH4)2CO3(pH8.0)洗脱,取0.4 mL洗脱后的醚提取物,加0.16 mL甲醇和0.15 mL三甲基硅基重氮甲烷、密封,在室温下放置30 min,再加入0.6 mL洗脱提取物,用装有0.3g硅胶和0.2g无水硫酸镁的巴斯德滴管过滤,用1 mL乙醚洗脱硅胶柱,所得样品置于-20℃冰箱中待分析[23]。
1.3.2 GC-MS 分析
萜类分析利用美国Thermo-Finnigan公司的TraceDSQ气相色谱-质谱仪(GC-MS),色谱柱利用DB-WAX毛细管柱(30 m×0.25 mm ID,膜厚0.25 μm),载气氢气;流速 2 mL/min,FID 温度300℃,进样口温度 220℃,进样1μL,分流比10∶1。起始温度40℃,停留3 min,以3℃/min速率升温至110℃,然后以10℃/min速率升温至180℃,再以15℃/min速率升温至250℃,保留10 min。质谱条件:EI离子源,电离能70ev。各成分通过与谱库(NISTZOOZ版)标准化合物的质谱图核对并分析后,进行定性,根据内标法进行定量[2-4,23]。
1.4 梢斑螟幼虫RNA和P含量测定
1.4.1 RNA 提取和含量测定
采用显微荧光测定法测试RNA含量。取3龄、4龄和5龄取食活跃期幼虫各30头(见1.1.2虫源),每组虫体的RNA用Trizol Reagent(Invitrogen)提取。RNA含量测定以酵母RNA为标准,采用对RNA有高度特异性的荧光试剂Ribo-Green测定。具体方法(Ribo Green RNA quantitation reagent and Minicell Adaptor Kit(P/N 3800-928))为:将RNA样品用TE稀释后取1 mL与等体积1∶2000稀释的RiboGreen试剂混匀,黑暗中放置5 min后在激发和发射波长分别为500 nm和525 nm条件下测定并记录。所有RNA的含量均为占虫体干重的百分比[21]。
1.4.2 P 含量测定
标准曲线的制作:取磷酸二氢钾0.4407g配置标准品溶液(每1mL含磷10.05μg)。取标准品溶液0、0.3、0.6、0.9、1.2 和 1.5 mL 分别置试管中,各加水至 3 mL,摇匀;再分别加入定磷试剂(2.5% 钼酸铵溶液∶3 mol/L 硫酸溶液∶水∶10%抗坏血酸=1∶1∶2∶1,用前配制)3 mL,摇匀;在 50℃水浴中反应 25 min。以0 号管作为空白对照,用UV-1240(uv mini)分光光度计在650 nm测定各样品液的吸光度。以溶液吸光度对其浓度作图,得标准曲线。
P含量的测定:采用抗坏血酸-钼蓝显色定磷法测定虫体中总P含量。取3龄、4龄和5龄取食活跃期幼虫各30头,将虫体在50℃烘48 h后经浓硫酸、浓硝酸硝化。样品溶液与定磷试剂混合摇匀,测定样品溶液的吸光度。利用磷标准曲线计算总磷含量,样品P含量为其占虫体干重的百分比[21-22]。
1.5 数据统计
数据用SPSS 10.0单因素方差(ANOVA)分析样品间的差异显著性,用a、b表示差异显著(P<0.05);用A、B表示差异极显著(P<0.01)。采用Sigmaplot 10.0作图。
2 结果与分析
2.1 松果梢斑螟下唇腺葡萄糖氧化酶活性变化
图1为不同组织中松果梢斑螟幼虫下唇腺GOX活性。可以看出,幼虫发育期内GOX活性呈动态变化,1果1虫、1果多虫、与小卷蛾共生和新梢4个处理梢斑螟幼虫GOX活性随虫龄而增加,有相同的变化趋势。但在幼虫脱皮期(每龄第4天)酶活性较低,取食活跃期(每龄第2天)酶活性显著增加。3龄、4龄和5龄取食活跃期GOX活性依次极显著升高(P<0.001),在5龄取食活跃期GOX活性最高,老熟幼虫GOX活性最低。
新梢、1果1虫、1果多虫、与小卷蛾共生等不同部位,其同一龄期梢斑螟幼虫GOX活性不同。3龄幼虫中,与小卷蛾共生的幼虫酶活性 >1果1虫和1果多虫的幼虫酶活性 >取食新梢的幼虫酶活性,但差异不显著。4龄幼虫中,与小卷蛾共生的幼虫酶活性最高(0.62±0.04),取食新梢的幼虫酶活性最低(0.42±0.08),取食球果(1果1虫和1果多虫)居中,三者差异显著。但1果多虫(0.50±0.02)和1果1虫(0.55±0.05)的幼虫酶活性差异不显著。5龄幼虫中,与小卷蛾共生的幼虫酶活性最高(0.81±0.07),显著高于新梢(0.57±0.05),其余之间差异不显著。
2.2 虫害球果和新梢萜类含量的变化
图2为虫害油松球果和新梢单萜、倍半萜和双萜含量变化。可以看出,单萜、倍半萜和双萜含量呈现先增加后降低的趋势,球果的萜类含量随虫害时间延长而明显下降。虫害1 d时,新梢和球果单萜总含量均显著增加(P<0.01),随虫害时间而逐渐降低,虫害15 d时,单萜含量最低。作为新梢和球果单萜主要成分的1R-α-蒎烯和D-柠檬烯含量与单萜总含量有一致的变化趋势,即虫害后先升后降,最后与对照水平接近(图1,表1,表2)。
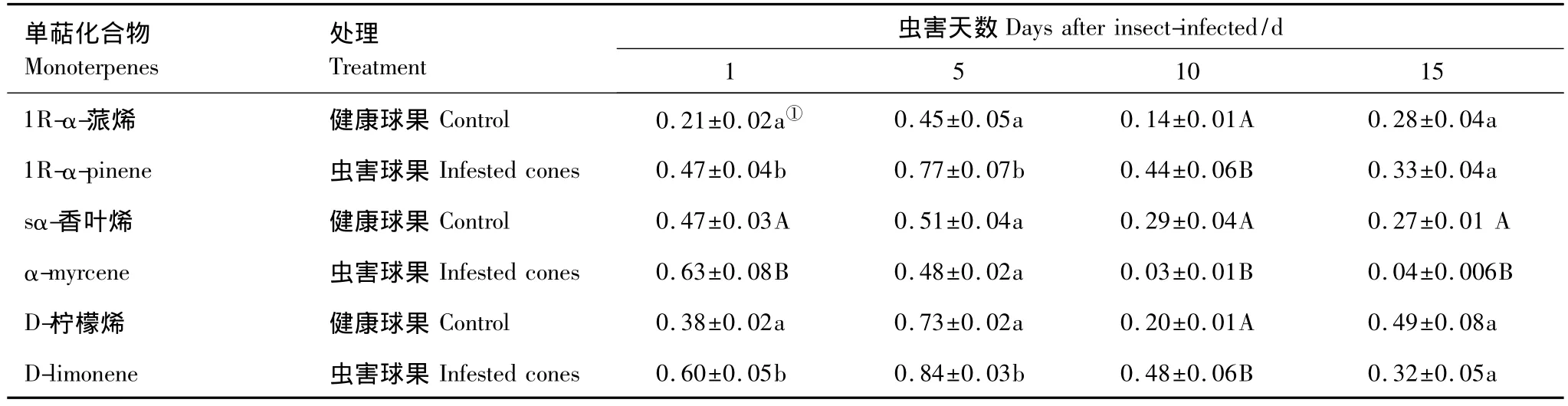
表1 虫害油松球果主要单萜成分含量随时间的变化/(mg/g鲜重)Table 1 Changes of major monoterpene content with time in the infested cones
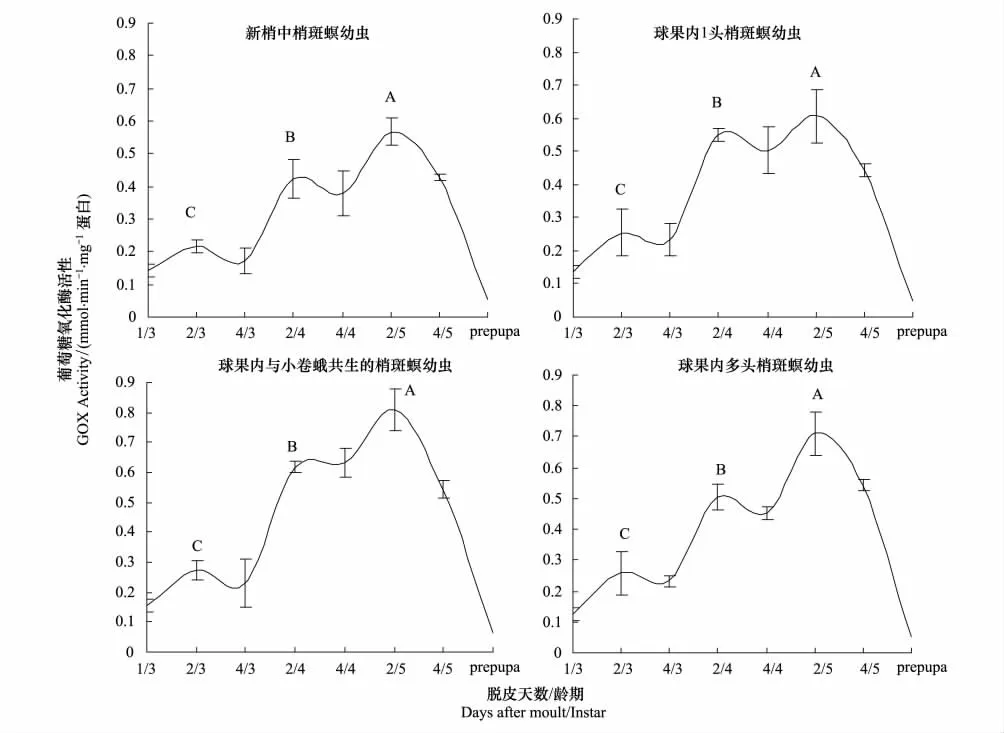
图1 不同组织中梢斑螟幼虫下唇腺葡萄糖氧化酶活性比较Fig.1 Glucose oxidase(GOX)activity in labial salivary glands of D.pryeri developed in four host tissues
油松球果和新梢中倍半萜含量较少,虫害后倍半萜含量明显下降。虫害初期,球果中倍半萜含量没有变化,随后显著下降;而新梢虫害初期倍半萜含量略有增加,随后明显下降。倍半萜总含量,球果略高于新梢(图1)。球果和新梢中主要倍半萜均为石竹烯、α-石竹烯和1-甲基-5-亚甲基-8-(1-异丙基)-1,6-环十二烯,其中石竹烯含量较大。在虫害球果,初期(第1天)石竹烯增加,随后显著下降;而其它2种成分虫害后显著降低(表3)。虫害后,新梢中的石竹烯显著增加,随后呈曲线波动;α-石竹烯虫害后显著增加,随后降低;1-甲基-5-亚甲基-8-(1-异丙基)-1,6-环十二烯在虫害初期含量变化不大,但在虫害10 d后,该成分检测不到(表4)。

表2 虫害新梢主要单萜成分随时间的变化/(mg/g鲜重)Table 2 Changes of major monoterpene content with time in the infested branches

图2 松果梢斑螟幼虫危害后油松球果及新梢萜类含量随时间的变化Fig.2 Changes of terpene content with time in fested cones and branches

表3 虫害球果主要倍半萜成分随时间的变化/(mg/g鲜重)Table 3 Changes of major sesquiterpene content with time in the infested cones
图2可以看出,球果虫害后,双萜含量显著增加,后期明显降低到正常水平。虫害第1天双萜含量达到最大(7.993 mg/g鲜重),是健康球果的1.7倍;虫害5 d时双萜含量是健康球果的1.4倍,第10天双萜含量开始下降,第15天时接近健康球果水平。新梢虫害后,其双萜含量初期显著增加,随后逐渐接近健康新梢,但虫害第15天时双萜含量又显著高于健康新梢(图1)。虫害球果和新梢显著增加双萜成分是松香酸甲酯、1-菲羧酸甲酯类和贝壳杉-16-烯-18-酸酯,新梢中松香酸甲酯含量较少。双萜主要成分的变化趋势与总含量一致:虫害后显著增加,后逐渐降低(表5,表6)。
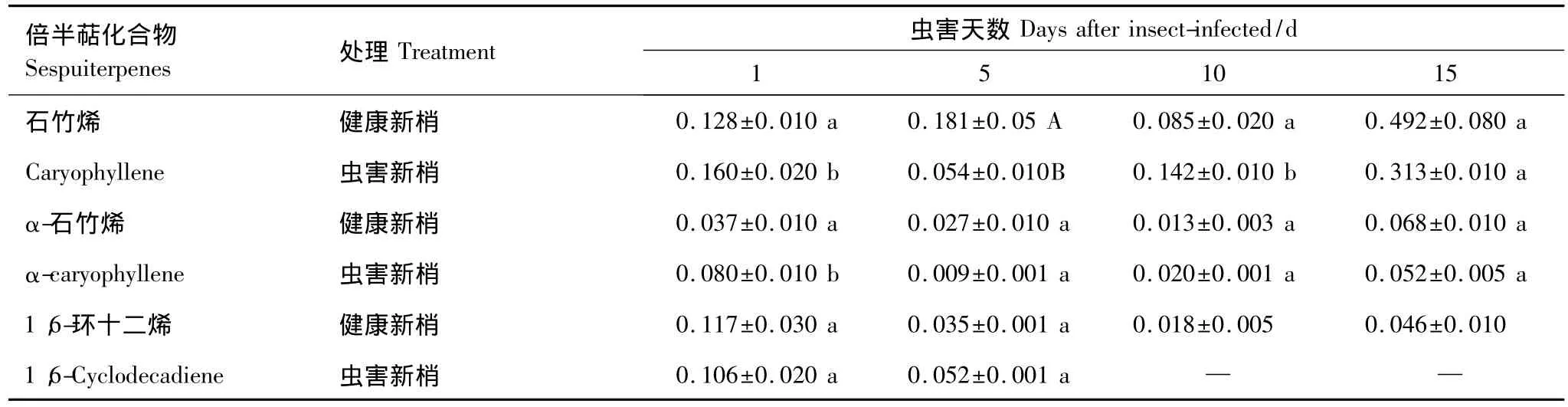
表4 虫害新梢主要倍半萜成分随时间的变化/(mg/g鲜重)Table 4 Changes of major sesquiterpene content with time in the infested branches
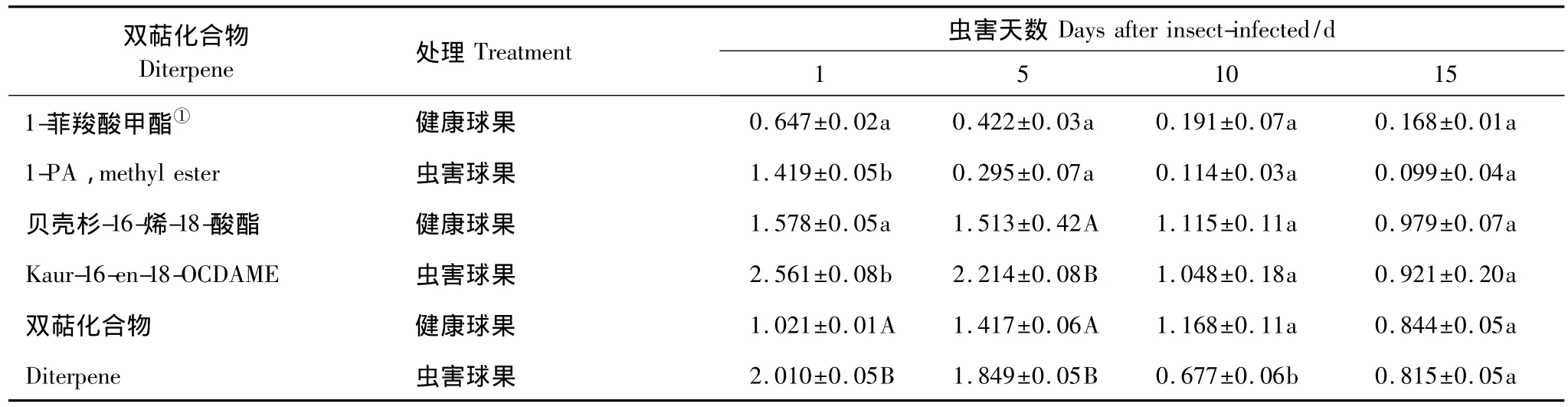
表5 虫害球果主要双萜成分随时间的变化/(mg/g鲜重)Table 5 Changes of major diterpene content with time in the infested cones
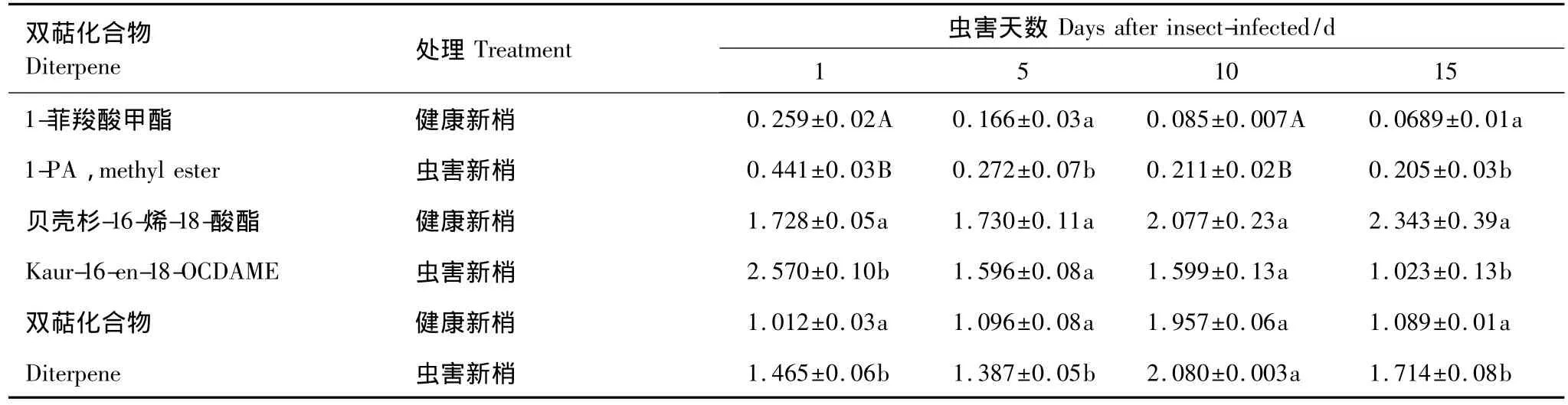
表6 虫害诱导新梢主要双萜成分随时间的变化/(mg/g鲜重)Table 6 Changes of major diterpene content with time in the infested branches
2.3 取食寄主不同部位对梢斑螟幼虫RNA和P含量的影响
根据生长率和虫体RNA与P含量高度耦合的理论[21],测定取食不同处理食物的梢斑螟幼虫RNA与P含量,可间接反映寄主食料对幼虫生长发育的影响。表7可以看出,取食3种不同处理寄主食物的梢斑螟幼虫,都以3龄幼虫的RNA含量和P含量最高,其次是4龄幼虫和5龄幼虫,差异极显著(P<0.01)。说明梢斑螟3龄幼虫的生长率较快,之后逐渐变缓。
就取食部位来看,3个龄期幼虫RNA含量均为:取食新梢幼虫 <取食小卷蛾危害球果幼虫<取食球果(1果1虫和1果多虫)幼虫,但无显著差异。就P含量来看,除4龄幼虫外,取食球果的梢斑螟幼虫P含量较高,取食新梢的幼虫P含量较低,差异不显著。说明3种处理食物对梢斑螟幼虫生长率影响无显著差异。
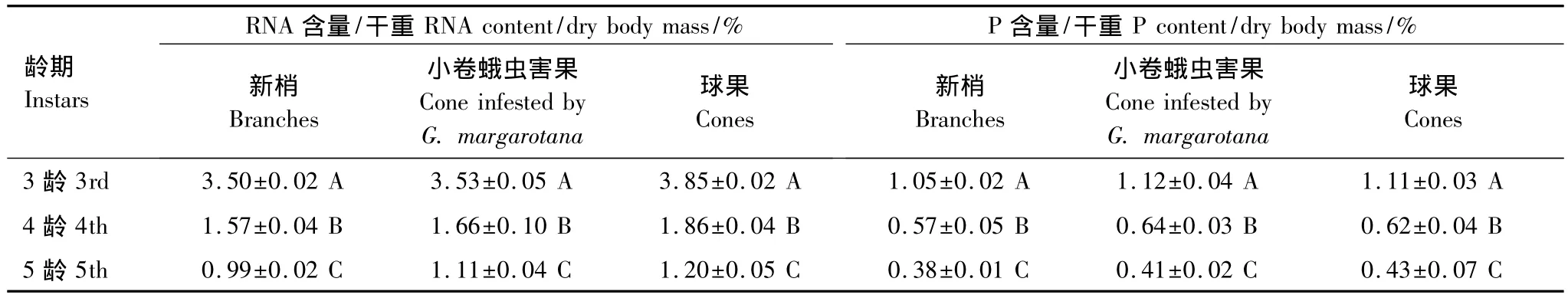
表7 取食寄主不同部位梢斑螟幼虫RNA和P含量比较Table 7 RNA and P content comparison of pine comeworm larvae in different host tissue
3 讨论
在北方油松良种基地,每年7月,羽化的松果梢斑螟成虫趋向虫害(干枯的)球果附近产卵;翌年5月,小卷蛾幼虫先期危害,引诱梢斑螟越冬幼虫趋向正在发育的虫害球果;同时,梢斑螟幼虫也危害健康球果和新梢[1-2,4]。在林区条件下,本研究选择梢斑螟与小卷蛾一起危害(与小卷蛾共生)的球果、有1头梢斑螟幼虫(1果1虫)的球果、有2—3头梢斑螟幼虫(1果多虫)的球果和梢斑螟危害新梢等4种处理,取食不同食料的梢斑螟幼虫下唇腺GOX活性测试表明,4种食料均以5龄幼虫GOX活性最高,显著高于4龄和3龄幼虫。并且,3个龄期的幼虫均表现为:球果中与小卷蛾共生的梢斑螟幼虫GOX活性 >球果中单一梢斑螟幼虫酶活性>新梢中幼虫酶活性,其中与小卷蛾共生的4龄和5龄幼虫GOX活性显著高于新梢内的幼虫。研究发现,多食性鳞翅目幼虫下唇腺GOX可显著抑制植物的防御。并且,人工饲料饲养的幼虫GOX活性显著高于寄主植物上发育的幼虫,同一昆虫不同寄主上幼虫GOX活性也显著不同,反映了食料质量(碳水化合物、蛋白等)影响幼虫GOX活性水平[14,18-19],这与本研究结果一致。
双萜(松脂酸)是针叶树组成和诱导防御的主要物质,可作为植食性昆虫的有毒成分而起到直接防御作用[26-28]。松脂萜类聚集在针叶树特化的松脂道中,虫害引起松脂道受损,伴有单萜挥发物的松脂在受害部位形成物理和化学障碍。而倍半萜在松脂中所占比例较小[5,28]。本研究结果显示,双萜(松脂酸)的基础量最大(5.0 mg/g 鲜重左右),而倍半萜则很少(0.3—0.5 mg/g 鲜重),单萜在1.5 mg/g 鲜重左右;虫害后,松脂酸变化在±2.5 mg/g鲜重,虫害球果随时间逐渐减少,第10天后低于2.7 mg/g鲜重;而新梢虫害后,松脂酸初期显著增加,随后降低,最后显著高于健康梢。虫害后,单萜的情况类似双萜,而倍半萜在虫害球果和新梢中,总体是逐渐减少的。与球果比较,新梢的防御物质(松脂酸)逐渐增加。以前研究中,比较了油松球果小卷蛾幼虫危害后,虫害球果萜类的变化,以及其它防御物质的增加[2-3],在本研究中没有比较2种害虫危害油松球果防御反应的差异。
与小卷蛾共生、油松球果和新梢3种食料的梢斑螟幼虫RNA和P含量显示,3龄幼虫RNA和P含量最高,随后逐渐减少,差异极显著,说明前期幼虫发育快,后期幼虫发育慢,符合生长率与虫体RNA和P含量高度耦合的生长率理论[21]。在3种食料中,3个龄期幼虫RNA和P含量与食料密切相关,但3种食料间幼虫RNA和P含量(即生长率)无显著差异,说明梢斑螟幼虫可通过GOX调节和抑制植物的防御[10-15,18-20]。但3种食料对梢斑螟存活、蛹重和生殖的影响还不清楚[19]。下唇腺GOX的高抑制作用是多食性鳞翅目幼虫应对寄主诱导防御的对策[18],而GOX及其产物H2O2通过激发水杨酸(SA)途径而抑制茉莉酸-乙烯(JA-ET)调控的植物防御,改变了植物基因表达,所以GOX能抑制寄主的直接和间接防御[12,20]。小卷蛾幼虫的先期危害,激发了JA途径调控的诱导防御,降低了食物质量[3,5],但后来的梢斑螟幼虫用GOX抑制松脂酸等防御物质[20],使其生长率与健康球果、新梢中的幼虫基本一致。
油松球果小卷蛾幼虫的先期危害,引诱梢斑螟越冬幼虫趋向球果取食[2],造成2种害虫对食料的竞争。在竞争中,小卷蛾幼虫处于劣势而部分死亡,梢斑螟幼虫的死亡率却很低[1]。但是,由于小卷蛾幼虫提前脱果下地,通过虫害球果挥发物(间接防御)而引诱寄生蜂,使受害球果内的梢斑螟幼虫寄生率提高,说明趋向受害球果的梢斑螟幼虫,面临着天敌的威胁[1-2,6]。球果受害后,抗营养、抗消化酶类和防御物质增加,营养质量下降[2-3],但本研究表明,梢斑螟幼虫通过GOX活性抑制植物的防御,使不同食料的幼虫生长率基本一致。后来羽化的梢斑螟成虫,通过虫害挥发物,趋向虫害球果附近产卵[4]。进一步证明,虫害寄主(诱导挥发物)暗示着竞争者和天敌的存在,寄主营养质量的下降,也代表着潜在寄主的存在[6],说明后来者具有特殊的克服机制。
[1] Li X G.Research on the damage mechanism of the cone insects pests of Chinese pine in Shaanxi,China.Journal of Northwest Sci-Tech University of Agriculture and Forestry:Natural Science Edition,2002,30(2):78-82.
[2] Li X G,Liu H X,Liu L P,Ma Y M.Study on host-plant volatiles affecting the host selection of Dioryctria pryeri.Scientia Silvae Sinicae,2006,42(3):71-78.
[3] Li X G,Liu H X,Hou H B,Gao W H.Response of Chinese pine cones to induction of exogenous methyl jasmonate and Gravitarmata margarotana larvae.Scientia Silvae Sinicae,2007,43(6):66-72.
[4] Li X G,Yang L J,Liu L P,Liu H X.Host selection of adult Dioryctria pryeri.Scientia Silvae Sinicae,2009,45(2):75-81.
[5] Li X G,Liu H X,Huang J.Molecular mechanisms of insect pests-induced plant defense.Chinese Journal of Applied Ecology,2008,19(4):893-900.
[6] Karban R,Baldwin I.Induced Responses to Herbivory.Chicago:University of Chicago Press,1997:104-165.
[7] Carroll M J,Schmelz E A,Meagher R L,Teal P E A.Attraction of Spodoptera frugiperda larvae to volatiles from herbivore-damaged maize seedlings.Journal of Chemical Ecology,2006,32(9):1911-1924.
[8] Fidgen L L,Sweeney J D.Fir coneworm,Dioryctria abietivorella(Grote)(Lepidoptera:Pyralidae),prefer cones previously exploited by the spruce cone maggots Strobilomyia neanthracina Michelsen and Strobilomyia appalachensis Michelsen(Diptera:Anthomyiidae).Canadian Entomologist,1996,128(6):1221-1224.
[9] Landolt P J,Brumley J A,Smtthhisler C L,Biddick L L,Hofstetter R W.Apple fruit infested with codling moth are more attractive to neonate codling moth larvae and possess increased amounts of(E,E)-α-farnesene.Journal of Chemical Ecology,2000,26(7):1685-1699.
[10] Musser R O,Hum-Musser S M,Eichenseer H,Peiffe M R,Ervin G,Murphy J B,Felton G W.Herbivory:Caterpillar saliva beats plant defenses:A new weapon emerges in the co-evolutionary arms race between plants and herbivores.Nature,2002,416(6881):599-600.
[11] Musser R O,Cipollini D,Hum-Musser S M,Williams S A,Brown J K,Felton G W.Evidence that the caterpillar salivary enzyme glucose oxidase provides herbivore offense in Solanaceous plants.Archives of Insect Biochemistry and Physiology,2005,58(2):128-137.
[12] Musser R O,Hum-Musser S M,Lee H K,DesRochers B L,Williams S A,Vogel H.Caterpillar labial saliva alters tomato plant gene expression.Journal of Chemical Ecology,2012,38(11):1387-1404.
[13] Bede J C,Musser R O,Felton G W,Korth K L.Caterpillar herbivory and salivary enzymes decrease transcript levels of Medicago truncatula genes encoding early enzymes in terpenoid biosynthesis.Plant Molecular Biology,2006,60(4):519-531.
[14] Hu Y H,Leung D W M,Kang L,Wang C Z.Diet factors responsible for the change of the glucose oxidase activity in labial salivary glands of Helicoverpa armigera.Archives of Insect Biochemistry and Physiology,2008,68(2):113-121.
[15] Peiffer M,Felton G W.The host plant as a factor in the synthesis and secretion of salivary glucose oxidase in larval Helicoverpa zea.Archives of Insect Biochemistry and Physiology,2005,58(2):106-113.
[16] Ferry N,Edwards M G,Gatehouse J A,Gatehouse A M R.Plant-insect interactions:molecular approaches to insect resistance.Current Opinion in Biotechnology,2004,15(2):155-161.
[17] Roda A L,Baldwin I T.Molecular technology reveals how the induced direct defenses of plants work.Basic and Applied Ecology,2003,4(1):15-26.
[18] Eichenseer H,Mathews M C,Powell J S,Felton G W.Survey of a salivary effector in caterpillars:glucose oxidase variation and correlation with host range.Journal of Chemical Ecology,2010,36(8):885-897.
[19] Babic B,Poisson A,Darwish S,Lacasse J,Merkx-Jacques M,Despland E,Bede J C.Influence of dietary nutritional composition on caterpillar salivary enzyme activity.Journal of Insect Physiology,2008,54(1):286-296.
[20] Diezel C,Von Dahl C,Gaqurrel E,Baldwin I T.Different lepidopteran elicitors account for cross-talk in herbivory-induced phytohormone signaling.Plant Physiology,2009,150(3):1576-1586.
[21] Elser J J,Watts T,Bitler B,Markow T A.Ontogenetic coupling of growth rate with RNA and P contents in five species of Drosophila.Functional Ecology,2006,20(5):846-856.
[22] Kyle M,Watts T,Schade J,Elser J J.A microfluorometric method for quantifying RNA and DNA in terrestrial insects.Journal of Insect Science,2003,3(1):1-7.
[23] Miller B,Madilao L L,Ralph S,Bohlmann J.Insect-induced conifer defense.White pine weevil and methyl jasmonate induce traumatic resinosis,de novo formed volatile emissions,and accumulation of terpenoid synthase and putative octadecanoid pathway transcripts in sitka spruce.Plant Physiology,2005,137(1):369-382.
[24] Bergmeyer H U,Gawehn K,Grassl M.Methods of Enzymatic Analysis.2nd ed.Vol.1.New York:Academic Press,1974:457-458.
[25] Bradford M M.A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding.Analytical Biochemistry,1976,72(7):248-254.
[26] Martin D,Tholl D,Gershenzon J,Bohlmnan J.Methyl jasmonate induces traumatic resin ducts,Terpenoid resin biosynthesis,and terpenoid accumulation in developing Xylem of Norway spruce stems.Plant Physiology,2002,129(3):1003-1018.
[27] Byun-McKay A,Godard K A,Toudefallah M,Martin D M,Alfaro R,King J,Bohlmann J,Aine L.Wound-induced terpene synthase gene expression in Sitka spruce that exhibit resistance or susceptibility to attack by the white pine weevil.Plant Physiology,2006,140(3):1009-1021.
[28] Martin D,Fäldt J,Bohlmann J.Functional characterization of nine Norway spruce TPS genes and evolution of Gymnosperm terpene synthases of the TPS-d subfamily.Plant Physiology,2004,135(4):1908-1927.
参考文献:
[1] 李新岗.油松球果害虫的危害机理研究.西北农林科技大学学报:自然科学版,2002,30(2):78-82.
[2] 李新岗,刘惠霞,刘拉平,马养民.影响松果梢斑螟寄主选择的植物挥发物成分研究.林业科学,2006,42(6):71-78.
[3] 李新岗,刘惠霞,侯慧波,高文海.油松球果对外源茉莉酸甲酯和虫害诱导的生化反应.林业科学,2007,43(3):66-72.
[4] 李新岗,杨立军,刘拉平,刘惠霞.松果梢斑螟成虫的寄主选择.林业科学,2009,45(2):75-81.
[5] 李新岗,刘惠霞,黄建.虫害诱导植物防御的分子机理研究进展.应用生态学报,2008,19(4):893-900.
