越冬过程中刺柏属4树种叶片生理指标变化及适应性综合评价
2013-12-14孙敬爽贾桂霞陶霞娟孙长忠
孙敬爽,贾桂霞,陶霞娟,孙长忠,①
(1.中国林业科学研究院华北林业实验中心,北京102300;2.北京林业大学园林学院,北京100083)
常绿针叶树是中国北方地区重要的城市绿化森林树种,通过引进国外观赏价值高的针叶树品种,不仅可以增加国内针叶树种质资源,而且可以进一步提升中国北方地区城市冬季绿化的景观效果、改善城市生态环境。
冬末和早春的低温、干旱及多风等逆境是影响中国北方地区常绿针叶树安全越冬的最主要生态因子。据报道[1],在北京、大连、沈阳和长春等北方城市的道路绿化中,约有10% ~30%的常绿针叶树受到生理干旱的伤害。目前,植物抗性研究多以低温、干旱等单一因素作为逆境因子,但植物在越冬过程中会受到环境条件的复杂性和植物对逆境因子的适应性等多种因素的影响,因而,通过科学有效的方法对引进观赏针叶树进行抗逆性综合评价,可为其成功引种与进一步推广应用提供理论依据。
主成分分析法是一种多变量的数学分析方法,目前已逐渐应用于植物的抗逆性综合评价[2]。针叶树的抗逆性较强,但有关针叶树抗逆性的研究却极少,且多采用单一指标或多个指标直接进行抗逆性评价[3-5],研究结果具有一定的局限性。
作者对越冬过程中4个引进的刺柏属(Juniperus Linn.)观赏针叶树品种的9项生理指标进行测定,并采用主成分分析法和隶属函数法对其越冬适应性进行综合评价,以期为观赏针叶树越冬适应性的鉴定、评价和预测提供实验依据。
1 材料和方法
1.1 实验地概况和实验材料
实验在华北林业实验中心苗圃进行。实验地位于北京市门头沟区,地理坐标为北纬39°28'~41°05'、东经 115°20'~117°30'。属于温带大陆性气候,年平均气温14.0℃,极端最低温-27.4℃,极端最高温42℃以上;年均日照总时数2 671.4 h,全年无霜期190~195 d,年平均降雨量640 mm。土壤为沙壤土-中壤,四周无建筑物遮挡。
供试的4个观赏树种为高山桧品种‘蓝星’(Juniperus squamata‘Blue Star’)、杂种桧品种‘黄鹿角’(J.×media‘Pfitzeriana Aurea’)、桧品种‘金羽’(J.chinensis‘Plumosa Aurea’)和欧洲桧品种‘金锥’(J.communis‘Gold Cone’),均 为 观 赏 矮 生 型 针叶树。均于2002年引进,苗高为10~15 cm、冠幅为20~30 cm2。经过3 a拱棚越冬保护后,于2006年在实验地露地越冬栽培;各品种15~20株苗木,3次重复。实验期间(2006年10月至2007年3月)各月的平均最低温度和空气相对湿度见表1。

表1 越冬期间各月平均最低温度和空气相对湿度Table 1 Mean minimum temperature and air relative humidity of each month during the overwintering period
1.2 方法
实验期间于每月的13日至15日,随机采集各品种供试植株顶端侧枝的成熟叶片,混合后用于各项生理指标测定。
膜电解质外渗率采用组织液外渗电导法[6]261测定,并略加改进。称取鲜叶0.2 g,加去离子水20 mL,用循环真空泵抽气5次,室温浸提1.5 h后测定电导值(R);然后置于沸水浴中处理20 min,冷却后测定电导值(R0),按照公式“膜电解质外渗率=(R/R0)×100%”计算膜电解质外渗率。
采用羟胺氧化法[7]测定超氧阴离子自由基(O-·2)产生速率。参照文献[8]的方法测定可溶性蛋白质含量。采用硫代巴比妥酸法[9]测定丙二醛(MDA)含量。采用氮蓝四唑(NBT)光化还原法[10]123测定超氧化物歧化酶(SOD)活性,以抑制氮蓝四唑光化还原的50%为1个酶活性单位。采用分光光度法[10]121-122测定过氧化氢酶(CAT)活性,以1 min内A240变化0.1为1个酶活性单位。采用愈创木酚法[6]164-165测定过氧 化 物 酶(POD)活性,以1 min内A470变化0.01为1个酶活性单位。参照Arrigoni[11]的方法测定抗坏血酸过氧化物酶(APX)活性。谷胱甘肽还原酶(GR)活性参照Knorzer等[12]的方法测定。
1.3 数据统计分析
对各指标的原始测定数据进行标准化,然后采用Varimax方法[13]旋转后得到各指标的载荷因子矩阵,建立主成分方程;参照文献[14]计算综合指标值CI(x),参照文献[15]计算各品种各综合指标的隶属函数值U(x)和权重以及各品种越冬适应性的综合评价值(D)。
采用SPSS 13.0和Excel 2007统计分析软件进行数据处理、相关性分析、主成分分析以及隶属函数的计算。
2 结果和分析
2.1 越冬期间4树种叶片生理指标的变化
越冬期间4种观赏针叶树叶片部分生理指标的测定结果见表2,叶片中5种抗氧化酶活性的测定结果见表3。
由表2可以看出:在4个指标中,叶片膜电解质外渗率变化幅度均较小。4树种叶片O-·2产生速率随时间延长均呈现先升高后降低的变化趋势,其中高山桧‘蓝星’、杂种桧‘黄鹿角’和桧‘金羽’O-·2产生速率的峰值在次年1月份(2007年1月)或2月份(2007年2月);而欧洲桧‘金锥’的产生速率峰值提前至12月份(2006年12月)且明显高于其他树种,说明在越冬过程中供试4树种叶片中的产生和清除系统平衡遭到破坏,使产生速率明显提高[16]。越冬过程中4种观赏针叶树叶片的可溶性蛋白质含量均逐渐增加,说明逆境胁迫诱发了可溶性蛋白质的合成以维持正常的生理功能。与10月份(2006年10月)相比,高山桧‘蓝星’、杂种桧‘黄鹿角’和桧‘金羽’3树种叶片的MDA含量在11月份(2006年11月)至次年3月份(2007年3月)期间明显降低,说明在越冬过程中其叶片细胞膜结构和膜脂含量可能发生变化,阻止了膜脂过氧化的发生;而欧洲桧‘金锥’叶片MDA含量在次年1月份达到峰值,明显高于10月份,说明该树种细胞膜受到一定的伤害。
表2 越冬期间刺柏属4种观赏针叶树种叶片部分生理指标的测定结果(±SD)1)Table 2 Determination result of some physiological indexes of leaves of four ornamental conifers of Juniperus Linn.during the overwintering period(±SD)1)

表2 越冬期间刺柏属4种观赏针叶树种叶片部分生理指标的测定结果(±SD)1)Table 2 Determination result of some physiological indexes of leaves of four ornamental conifers of Juniperus Linn.during the overwintering period(±SD)1)
?
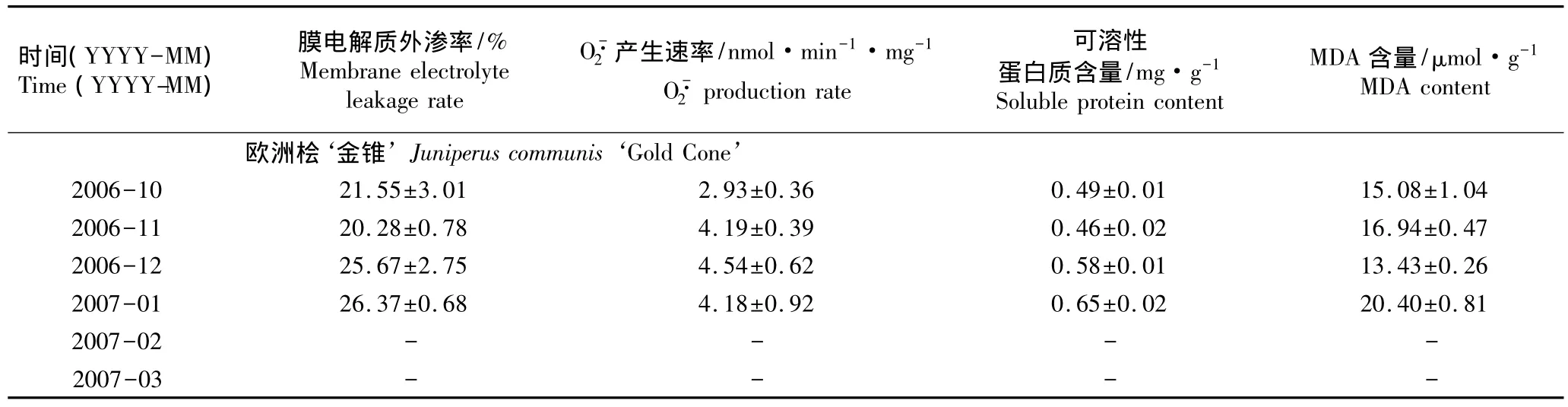
续表2 Table 2(Continued)
表3 越冬期间刺柏属4种观赏针叶树种叶片5种抗氧化酶活性的测定结果(±SD)1)Table 3 Determination result of activity of five antioxidant enzymes of leaves of four ornamental conifers of Juniperus Linn.during the overwintering period(±SD)1)
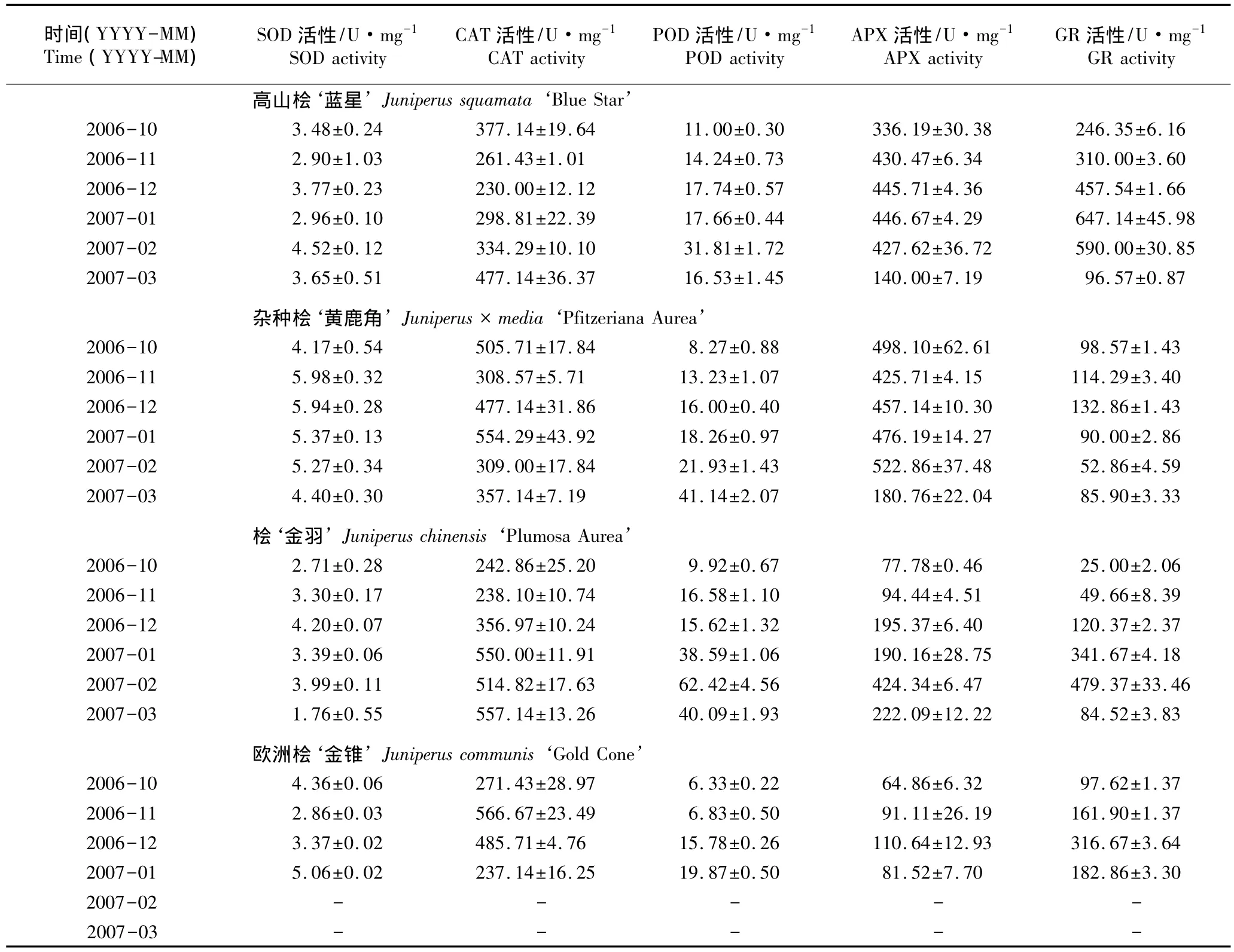
表3 越冬期间刺柏属4种观赏针叶树种叶片5种抗氧化酶活性的测定结果(±SD)1)Table 3 Determination result of activity of five antioxidant enzymes of leaves of four ornamental conifers of Juniperus Linn.during the overwintering period(±SD)1)
1)-:因苗木死亡,无测定数据Without determination datum because of seedling dead.
?
由表3可以看出:在越冬过程中4树种叶片中5种抗氧化酶活性的变幅有较大差异,其中SOD、CAT以及GR活性的变化无明显规律,而10月份至次年2月份POD和APX活性则总体上呈增加的趋势。
由此可见,在越冬过程中,由于受多种逆境因子的综合影响,供试4树种叶片的各项生理指标变幅明显不同,因此不能采用单一指标评价供试4树种的越冬适应能力。
2.2 越冬期间4树种叶片生理指标的相关性分析
越冬过程中4种观赏针叶树叶片9项生理指标的相关系数见表4。由表4可以看出:MDA含量与膜电解质外渗率、O-·2产生速率、可溶性蛋白质含量、CAT活性和POD活性均有显著或极显著的相关性;APX活性与O-·2产生速率、可溶性蛋白质含量、SOD活性和POD活性均有显著或极显著的相关性。O-·2产生速率与膜电解质外渗率、可溶性蛋白质含量呈显著负相关;SOD活性与膜电解质外渗率和CAT活性分别呈极显著正相关和显著负相关;POD活性与可溶性蛋白质含量和CAT活性均呈极显著正相关。由于这些指标间具有较为显著的相关性,使其所提供的信息重叠,直接利用这些指标难以准确评价4树种越冬适应性的强弱。
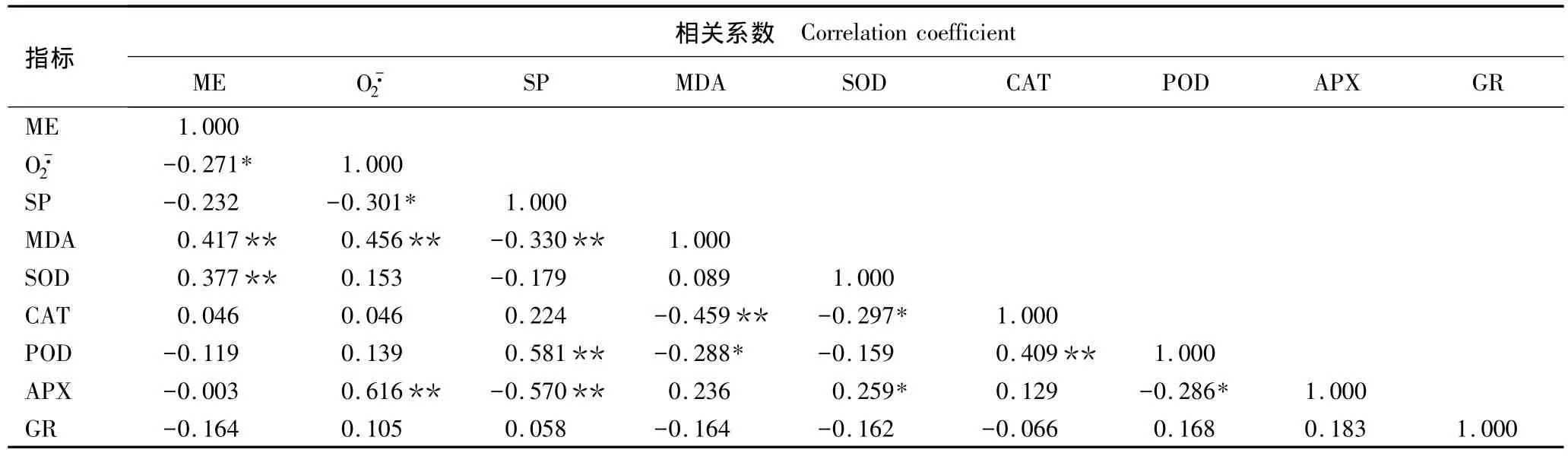
表4 越冬过程中刺柏属4种观赏针叶树叶片生理指标的相关性分析1)Table 4 Correlation analysis of physiological indexes of leaves of four ornamental conifers of Juniperus Linn.during the overwintering period1)
2.3 越冬期间4树种叶片生理指标的主成分分析
越冬过程中供试4种观赏针叶树叶片9项生理指标的主成分分析结果见表5。由表5可见:9项生理指标可转化为4项综合指标,即第1、第2、第3和第4 主成分,贡献率分别为 29.66%、19.93%、16.61%和11.62%,累计贡献率达到77.82%。根据9项指标的标准化值和综合指标的标准化特征向量分别得到第1、第2、第3和第4主成分的回归方程:CI(1)=-0.010x1+0.559x2- 0.797x3+0.672x4+0.425x5-0.448x6- 0.624x7+0.704x8- 0.068x9;CI(2)=-0.748x1+0.633x2+0.191x3+0.308x4- 0.424x5+0.278x6+0.454x7+0.282x8+0.399x9;CI(3)=0.515x1+0.280x2- 0.202x3- 0.524x4+0.226x5+0.637x6+0.229x7+0.546x8+0.174x9;CI(4)=0.003x1+0.280x2+0.218x3+0.241x4+0.372x5+0.187x6+0.321x7-0.100x8-0.759x9。
主成分特征向量代表了各变量对相应主成分的作用权数,即各单项指标对综合指标的贡献大小[17]。由表5还可以看出:在第1主成分中特征向量较大的指标是可溶性蛋白质含量,其次是APX活性和MDA含量。植物在低温和干旱等逆境胁迫下会诱导产生新的蛋白质以维持植物细胞较低的渗透势;而MDA是膜脂过氧化的最终产物,其含量常作为细胞膜损伤程度的判定指标,反映细胞膜脂过氧化程度和植物对逆境条件反应的强弱[18],因而,第1主成分可以定义为生物膜功能因子。第2主成分中特征向量较大的是膜电解质外渗率,其次是产生速率。由于膜电解质外渗率可以反映膜系统的完整性,而作为活性氧自由基之一对细胞膜有明显的毒害作用[19],因此第2主成分可以定义为膜结构稳定因子。在第3主成分中特征向量较大的是CAT活性,其次是APX活性;在第4主成分中特征向量较大的是GR活性,其次是SOD和POD活性。在逆境胁迫中,植物主要通过抗氧化酶系统和抗氧化剂来清除活性氧,因此,第3和第4主成分可定义为清除活性氧因子。
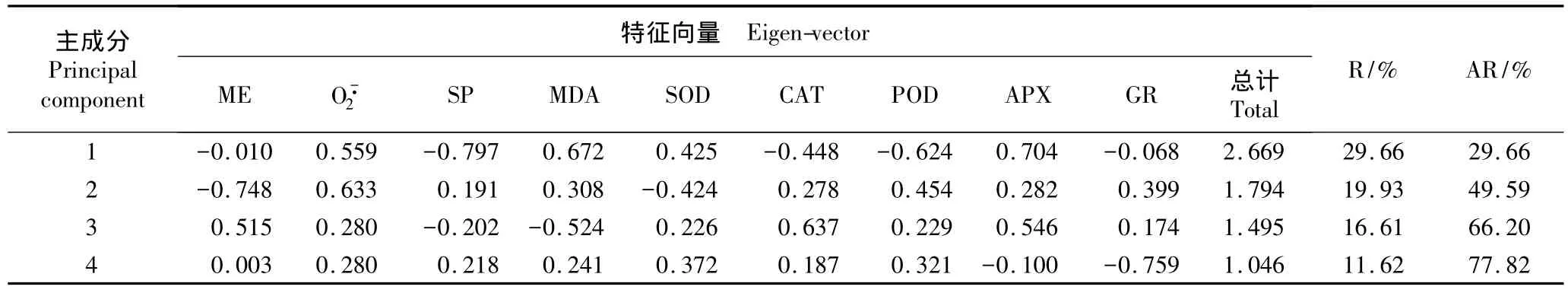
表5 越冬过程中刺柏属4种观赏针叶树叶片生理指标的主成分分析结果1)Table 5 Result of principal component analysis of physiological indexes of leaves of four ornamental conifers of Juniperus Linn.during the overwintering period1)
2.4 越冬期间4树种越冬适应性的综合评价
根据公式[15]计算供试4种观赏针叶树越冬适应性的各项评价指标,结果见表6。
2.4.1 隶属函数分析 由表6可见:对于同一综合指标的隶属函数值,如CI(1)而言,高山桧‘蓝星’在11月份(2006年11月)的 U(1)最大(为1.000),表明高山桧‘蓝星’在CI(1)这一综合指标上表现为越冬适应性最强;欧洲桧‘金锥’在次年1月份(2007年1月)的U(1)最小(为0.000),表明欧洲桧‘金锥’在CI(1)这一综合指标上越冬适应能力最差。
2.4.2 权重的确定 根据4项综合指标的贡献率(表5)确定4项综合指标的权重分别为0.381、0.256、0.213 和 0.149。
2.4.3 越冬适应性综合评价 根据4项综合指标的U(x)与权重,获得4种观赏针叶树在越冬过程中的不同月份的综合评价值(D),D值大小反映了其越冬适应性的强弱。由表6可以看出:同一树种综合评价值在不同月份变化规律不一致。随着逆境胁迫时间的延长,高山桧‘蓝星’和桧‘金羽’分别在次年1月份和2月份(2007年2月)D值较高,分别为0.768和0.520;杂种桧‘黄鹿角’和欧洲桧‘金锥’在10月份(2006年10月)D值最高,分别为0.639和0.483。
在10月份至次年2月份,同一月份4树种的D值由大到小均依次排序为:高山桧‘蓝星’、杂种桧‘黄鹿角’、桧‘金羽’、欧洲桧‘金锥’。因此,在越冬过程中4树种的越冬适应性由高到低依次为:高山桧‘蓝星’、杂种桧‘黄鹿角’、桧‘金羽’、欧洲桧‘金锥’。
3 讨 论
植物的越冬适应性是一个复杂的综合性状,受长期低温、干旱、多风等多种自然因素的交叉影响;且植物的抗逆性是受多基因控制的数量性状,用单一指标难以真实、准确地反映植物越冬适应性的强弱,而采用多种指标进行综合评价则较为科学和准确。本研究结果表明:在越冬过程中供试4树种多项生理指标(除GR活性外)存在一定的相关性,造成信息重叠及交叉等现象,因此不能直接用多项生理指标进行越冬适应性的综合评价。
利用主成分分析法将原来数量较多且彼此相关的指标转化成数量较少且相互独立的综合指标,克服了信息的重叠性和指标之间的相关性,从而使评价结果更为科学合理。主成分分析结果表明:在越冬过程中,供试4树种9项生理指标的测定结果可以转化成4项综合指标,累计贡献率达到77.82%。根据生物学功能定位可分为生物膜功能因子(可溶性蛋白质含量和MDA含量)、膜结构稳定因子(膜电解质外渗率和产生速率)和清除活性氧因子(CAT、APX、GR、SOD和POD活性),可作为供试4树种越冬适应性的鉴定指标,这与陈银萍等[20]和江萍等[21]的观点基本一致。
由于单一综合指标的大小不能完全决定某一植物越冬适应性的强弱,因此,根据4项综合指标的贡献率获得相应的隶属函数值,并根据各综合指标的相对重要性(权重)进行加权,得到4树种越冬适应性的综合评价值(D),从而使各树种间越冬适应性的差异具有可比性。根据D值确定供试4树种在北京地区的适应性:高山桧‘蓝星’在北方地区越冬适应性最强,杂种桧‘黄鹿角’和桧‘金羽’次之,欧洲桧‘金锥’的越冬适应性最差,这一结果与4树种在北京地区田间的越冬表现[22]存在一定差异,推测主要与冬季复杂的自然因素有关。针叶树在越冬过程中除受到低温和干旱等因素影响外,光抑制和光氧化等现象也非常严重[23]。因此,在树种的越冬适应性研究中,应采用更多的生物学指标才能更为客观、准确地评价针叶树的越冬适应性,使其抗逆性鉴定更有预见性。
[1]陆庆轩,宁祝华.生理干旱对北方地区城市森林中针叶树生长的影响[J].中国城市林业,2010,8(1):52-54.
[2]袁志发,周静芋.多元统计分析[M].北京:科学出版社,2002:90.
[3]周永学,龚月桦,樊军锋.4种松树的抗寒性的比较[J].西北林学院学报,2011,26(5):69-71.
[4]仲 强,康 蒙,郭 明,等.浙江天童常绿木本植物的叶片相对电导率及抗寒性[J].华东师范大学学报:自然科学版,2011(4):45-52.
[5]倪福太,李长有,王占武,等.四种松属植物叶的抗寒抗旱特点研究[J].吉林师范大学学报:自然科学版,2012,32(2):110-112.
[6]李合生.植物生理生化实验原理和技术[M].北京:高等教育出版社,2000.
[7]王爱国,罗广华.植物的超氧物自由基与羟胺反应的定量关系[J].植物生理学通讯,1990(6):55-57.
[8]中国科学院上海植物生理研究所,上海市植物生理学会.现代植物生理学实验指南[M].北京:科学出版社,1999:308-309.
[9]HEALTH R L,PARKER L.Photoperoxidation in isolated chloroplasts:Ⅰ.Kinetics and stoichiometry of fatty acid peroxidation[J].Archives of Biochemistry and Biophysics,1968,125:189-198.
[10]张志良.植物生理学实验指导[M].3版.北京:高等教育出版社,2003.
[11]ARRIGONI O,De GARA L,TOMMASI F,et al.Changes in the ascorbate system during seed development of Vicia faba L.[J].Plant Physiology,1992,99:235-238.
[12]KNORZER O C,DURNER J,BOGER P.Alterations in the antioxidative system of suspension-cultured soybean cells(Glycine max)induced by oxidative stress[J].Physiologia Plantarum,1996,97:388-396.
[13]张 云,林 凡.基于主成分分析的木麻黄抗旱性评价[J].江苏农业科学,2008(5):159-162.
[14]林海明.如何用SPSS软件进一步算出主成分得分值[J].统计与信息论坛,2007,22(5):15-17.
[15]曾小玲,方淑桂,陈文辉,等.不同大白菜品种苗期耐热性的综合评价[J].福建农业学报,2010,25(2):183-186.
[16]王 萍,张成军,陈国祥,等.低温对水稻剑叶膜脂过氧化和脂肪酸组分的影响[J].作物学报,2006,32(6):568-572.
[17]王凯红,刘向平,张乐华,等.5种杜鹃幼苗对高温胁迫的生理生化响应及耐热性综合评价[J].植物资源与环境学报,2011,20(3):29-35.
[18]CHENG F Y,HSU S Y,KAO C H.Nitrc oxide counteracts the senescence of detached rice leaves induced by dehydration and polyethylene glycol but not by sorbitol[J].Plant Growth Regulation,2002,38:265-272.
[19]奉 斌,代其林,王 劲.非生物胁迫下植物体内活性氧清除酶系统的研究进展[J].绵阳师范学院学报,2009,28(11):50-53,77.
[20]陈银萍,张满效,陈 拓,等.圆柏属常绿木本植物叶片抗氧化系统季节变化与抗冷冻的关系[J].生态学杂志,2006,25(11):1318-1322.
[21]江 萍,孙向宁.华北落叶松酶活性及抗逆性机理研究[J].山西林业科技,2004(1):11-13.
[22]孙敬爽.引进矮生型针叶树繁殖技术及适应性研究[D].北京:北京林业大学园林学院,2007:61-83.
[23]PERKS M P,OSBORNE B A,MITCHELL D T.Rapid predictions ofcold tolerance in Douglas-firseedlingsusing chlorophyll fluorescence after freezing[J].New Forests,2004,28:49-62.
