土壤Cu胁迫对观赏牡丹生长、生理及Cu富集能力的影响
2013-12-14董春兰徐迎春陈亚华程家高
董春兰,徐迎春,①,陈亚华,程家高
(1.南京农业大学:a.园艺学院,b.生命科学学院,江苏南京210095;2.安徽铜陵高科技农业示范园,安徽铜陵244131)
牡丹(Paeonia suffruticosa Andr.)具有很高的观赏价值和深厚的文化内涵,深受国人喜爱,在国际上也享有盛誉。牡丹育苗一般采用嫁接法,近年来在牡丹种苗产区广泛用凤丹(Paeonia suffruticosa‘Feng Dan’)根作为牡丹嫁接的砧木,采用凤丹实生苗作砧木,比芍药(Paeonia lactiflora Pall.)砧木育砧期短;与牡丹自砧苗和以芍药为砧木嫁接的牡丹苗相比,以凤丹为砧木嫁接的牡丹苗成活率高、抗逆性强、苗木质量高[1-2]。据报道:凤丹可耐受一定浓度的 Cu胁迫[3],并在安徽凤凰山铜尾矿区生长良好[4];凤丹对600 mg·kg-1以下的Cu胁迫具有较强耐性,对于900 mg·kg-1Cu胁迫也具有一定的耐性[5];并且,凤丹对Cu污染土壤的修复效果明显[6]。
有研究者认为:嫁接可以缓解Cu胁迫对黄瓜(Cucumis sativus Linn.)的毒害作用[7-9]。据此推测,将牡丹嫁接在凤丹砧木上,也有可能提高牡丹的耐Cu能力。为此,作者采用以凤丹为砧木的牡丹品种‘肉芙蓉’(‘Roufurong’)5年生嫁接苗为实验材料,研究了不同质量浓度土壤Cu胁迫对其生长和生理指标的影响,并分析了其根、茎和叶中的 P、K、Ca、Mg、Cu、Fe、Mn和Zn含量以及Cu富集和转运系数,以期揭示牡丹嫁接苗的耐Cu性及其耐性机制,并探明牡丹修复Cu污染土壤的潜力。
1 材料和方法
1.1 材料
实验在南京农业大学园艺学院观赏植物生理生态研究室的塑料大棚内进行。以凤丹实生苗为砧木嫁接牡丹品种‘肉芙蓉’,选用植株大小及长势均较一致的5年生嫁接苗为实验材料。
1.2 方法
1.2.1 土壤Cu胁迫处理 于2009年10月15日进行盆栽实验。盆栽土为采集于原南京农业大学芳华园艺中心的表层壤土与黄沙按体积比2∶1拌匀的混合土壤。土壤中,有机质含量21.34 g·kg-1、全氮含量0.75 g·kg-1、速效 P 含量 11.67 mg·kg-1、速效 K 含量115.73 mg·kg-1、有效态Mg含量 64.39 mg·kg-1、有 效 态 Ca 含 量1 218.06 mg·kg-1、有效态Fe含量9.77 mg·kg-1、有效态 Mn含量128.64 mg·kg-1、有效态 Zn 含量18.65 mg·kg-1以及 Cu 含量 12.8 mg·kg-1,土壤 pH 6.62。
将混合土壤装入内径39.0 cm、高度36.5 cm的栽培盆内,每盆24 kg,按实验设定的Cu处理浓度将土壤与铜盐(用硫酸铜与碱式碳酸铜按Cu2+摩尔比1∶1的比例配制)混合均匀,设置低(300 mg·kg-1)、中(600 mg·kg-1)、高(900 mg·kg-1)3个Cu处理水平,以不添加铜盐的混合土壤为对照(CK)。每盆栽植1株,每处理3盆,每盆视为1个重复。实验期间,采用干透浇透的方式进行浇水,夏季覆盖遮阳网并注意通风,各处理组的管理方式均保持一致。
1.2.2 指标测定 采用单株取样法于2011年6月中旬叶片生长的旺盛期采集嫁接苗的叶片用于各项生理指标测定。其中,丙二醛(MDA)含量测定参照李合生[10]的方法;超氧化物歧化酶(SOD)活性测定采用NBT法[11-12];过氧化物酶(POD)活性测定采用愈创木酚法[13-14];叶绿素含量测定采用乙醇提取法[15]。
于2011年9月中旬测定各处理植株的各项生长指标。直接用直尺测量新枝长度、主根长度和一级侧根长度;用游标卡尺测量根茎直径;采用剪纸法测定叶面积;采用打孔法测定比叶质量;并统计一级侧根的数量。
于2011年9月中旬采集各处理植株,将植株分为根、茎、叶3部分,用自来水将植株各部分反复冲洗干净,再用去离子水清洗3遍,吸干表面水分后置于105℃杀青30 min,然后于80℃干燥至恒质量,研磨后过60目筛;用V(浓硝酸)∶V(高氯酸)=3∶1的混合酸对样品进行消煮,然后采用Optima 5300 DV等离子光谱质谱仪(美国PE公司)测定样品中P、K、Ca、Mg、Cu、Fe、Mn 和 Zn的含量。
1.3 数据统计分析
按下列公式分别计算不同部位Cu的富集系数和转运系数:Cu富集系数=植物组织中Cu含量/土壤中Cu含量;Cu转运系数=库器官Cu含量/源器官Cu含量。
采用SPSS 20.0统计分析软件对实验数据进行方差分析,并采用LSD检验进行多重比较。
2 结果和分析
2.1 土壤Cu胁迫对牡丹嫁接苗生长的影响
在土壤Cu胁迫条件下牡丹品种‘肉芙蓉’5年生嫁接苗的生长状况见表1。由表1可以看出:经不同质量浓度Cu胁迫处理后嫁接苗当年生新枝长度和一级侧根长度或高于对照或低于对照但均无显著差异(P>0.05)。经300 mg·kg-1Cu处理后嫁接苗叶面积大于对照但差异不显著;而经600和900 mg·kg-1Cu处理后叶面积均小于对照,其中仅后者与对照有显著差异(P<0.05),表明900 mg·kg-1Cu处理显著抑制嫁接苗叶面积的扩展。经不同质量浓度Cu胁迫处理后嫁接苗比叶质量和主根长度均低于对照,其中,仅900 mg·kg-1Cu处理组的比叶质量和主根长度与对照有显著差异。经不同质量浓度Cu胁迫处理后嫁接苗的根茎直径均高于对照,其中600 mg·kg-1Cu处理组根茎直径与对照有显著差异。经不同质量浓度Cu胁迫处理后嫁接苗一级侧根数量均少于对照且差异达显著水平,但各处理组间差异不显著。
表1 土壤Cu胁迫对牡丹品种‘肉芙蓉’5年生嫁接苗生长指标的影响(±SE)1)Table 1 Effect of Cu stress in soil on growth indexes of 5-year-old grafted seedlings of Paeonia suffruticosa‘Roufurong’(±SE)1)

表1 土壤Cu胁迫对牡丹品种‘肉芙蓉’5年生嫁接苗生长指标的影响(±SE)1)Table 1 Effect of Cu stress in soil on growth indexes of 5-year-old grafted seedlings of Paeonia suffruticosa‘Roufurong’(±SE)1)
1)同列中不同的小写字母表示经LSD检验差异显著(P<0.05)Different small letters in the same column indicate the significant difference by LSD test(P<0.05).
?
2.2 土壤Cu胁迫对牡丹嫁接苗生理指标的影响
在土壤Cu胁迫条件下牡丹品种‘肉芙蓉’5年生嫁接苗叶片中叶绿素含量的变化见表2;叶片中SOD和POD活性以及MDA含量的变化见表3。
2.2.1 对叶绿素含量的影响 经过2年的土壤Cu胁迫处理,所有嫁接苗的叶片均未表现出明显的黄化现象。由叶绿素含量的测定结果(表2)可见:经不同质量浓度Cu胁迫处理后嫁接苗叶片中的叶绿素a、叶绿素b和总叶绿素含量或高于对照或低于对照但与对照均无显著差异(P>0.05),说明Cu胁迫对牡丹品种‘肉芙蓉’嫁接苗叶绿素的合成和积累没有明显影响。但各处理组叶绿素a/b比值均低于对照,且随Cu质量浓度的提高呈现出逐渐减小的趋势,其中,600和900 mg·kg-1Cu处理组的叶绿素a/b比值与对照有显著差异(P<0.05),表明中、高质量浓度Cu胁迫处理对牡丹品种‘肉芙蓉’5年生嫁接苗叶绿素组成有较大影响,其中,对叶绿素a合成和积累的抑制作用较大。
表2 土壤Cu胁迫对牡丹品种‘肉芙蓉’5年生嫁接苗叶片中叶绿素含量的影响(±SE)1)Table 2 Effect of Cu stress in soil on chlorophyll content in leaf of 5-year-old grafted seedlings of Paeonia suffruticosa‘Roufurong’(±SE)1)

表2 土壤Cu胁迫对牡丹品种‘肉芙蓉’5年生嫁接苗叶片中叶绿素含量的影响(±SE)1)Table 2 Effect of Cu stress in soil on chlorophyll content in leaf of 5-year-old grafted seedlings of Paeonia suffruticosa‘Roufurong’(±SE)1)
1)同列中不同的小写字母表示经LSD检验差异显著(P<0.05)Different small letters in the same column indicate the significant difference by LSD test(P<0.05).
?
2.2.2 对SOD和POD活性及MDA含量的影响 由表3可以看出:经300和600 mg·kg-1Cu胁迫处理后牡丹嫁接苗叶片的SOD活性显著高于对照;经900 mg·kg-1Cu胁迫处理后叶片SOD活性虽然较前2个处理有所降低且也高于对照,但与对照的差异未达到显著水平。经300 mg·kg-1Cu胁迫处理后叶片的POD活性显著高于对照,而经600和900 mg·kg-1Cu处理后嫁接苗叶片的POD活性均低于对照,其中600 mg·kg-1Cu处理组叶片POD活性与对照有显著差异。说明在一定质量浓度土壤Cu胁迫条件下,牡丹嫁接苗体内的SOD和POD活性受到激发,起到保护酶的作用;但Cu质量浓度高于一定范围时,SOD和POD活性受到明显抑制,且POD活性较SOD活性更易受Cu胁迫的抑制。
表3 土壤Cu胁迫对牡丹品种‘肉芙蓉’5年生嫁接苗叶片中SOD和POD活性及MDA含量的影响(±SE)1)Table 3 Effect of Cu stress in soil on activities of SOD and POD and MDA content in leaf of 5-year-old grafted seedlings of Paeonia suffruticosa‘Roufurong’(±SE)1)

表3 土壤Cu胁迫对牡丹品种‘肉芙蓉’5年生嫁接苗叶片中SOD和POD活性及MDA含量的影响(±SE)1)Table 3 Effect of Cu stress in soil on activities of SOD and POD and MDA content in leaf of 5-year-old grafted seedlings of Paeonia suffruticosa‘Roufurong’(±SE)1)
1)同列中不同的小写字母表示经LSD检验差异显著(P<0.05)Different small letters in the same column indicate the significant difference by LSD test(P<0.05).
?
由表3还可见:经300 mg·kg-1Cu胁迫处理后叶片中 MDA含量低于对照,而经600和900 mg·kg-1Cu处理后MDA含量均高于对照,其中600 mg·kg-1Cu处理组MDA含量与对照有显著差异,说明在中、高质量浓度Cu胁迫条件下嫁接苗的膜脂过氧化加剧,Cu胁迫对细胞膜造成一定的损伤。
2.3 土壤Cu胁迫对牡丹嫁接苗各部位矿质元素含量的影响
经不同质量浓度Cu胁迫处理后牡丹品种‘肉芙蓉’5年生嫁接苗不同部位大量元素P、K、Ca和Mg含量的测定结果见表4;不同部位微量元素Cu、Fe、Mn和Zn含量的测定结果见表5。
2.3.1 对大量元素含量的影响 由表4可知:经不同质量浓度Cu处理后叶片中的P含量均高于对照、茎中的P含量均低于对照、根中的P含量或高于对照或低于对照,其中,经600和900 mg·kg-1Cu处理后叶片中的P含量与对照差异显著,经300 mg·kg-1Cu处理后茎中P含量与对照差异显著,而各处理组根中的P含量与对照均无显著差异。
经不同质量浓度Cu处理后叶片和根中的K含量均高于对照,茎中的K含量或高于对照或低于对照,其中,叶片K含量在300和900 mg·kg-1Cu胁迫条件下与对照有显著差异,茎中K含量在300 mg·kg-1Cu胁迫条件下与对照有显著差异,而根中的K含量则在600和900 mg·kg-1Cu胁迫条件下与对照差异显著。
表4 土壤Cu胁迫对牡丹品种‘肉芙蓉’5年生嫁接苗不同部位大量元素含量的影响(±SE)1)Table 4 Effect of Cu stress in soil on macro element content in different parts of 5-year-old grafted seedlings of Paeonia suffruticosa‘Roufurong’(±SE)1)
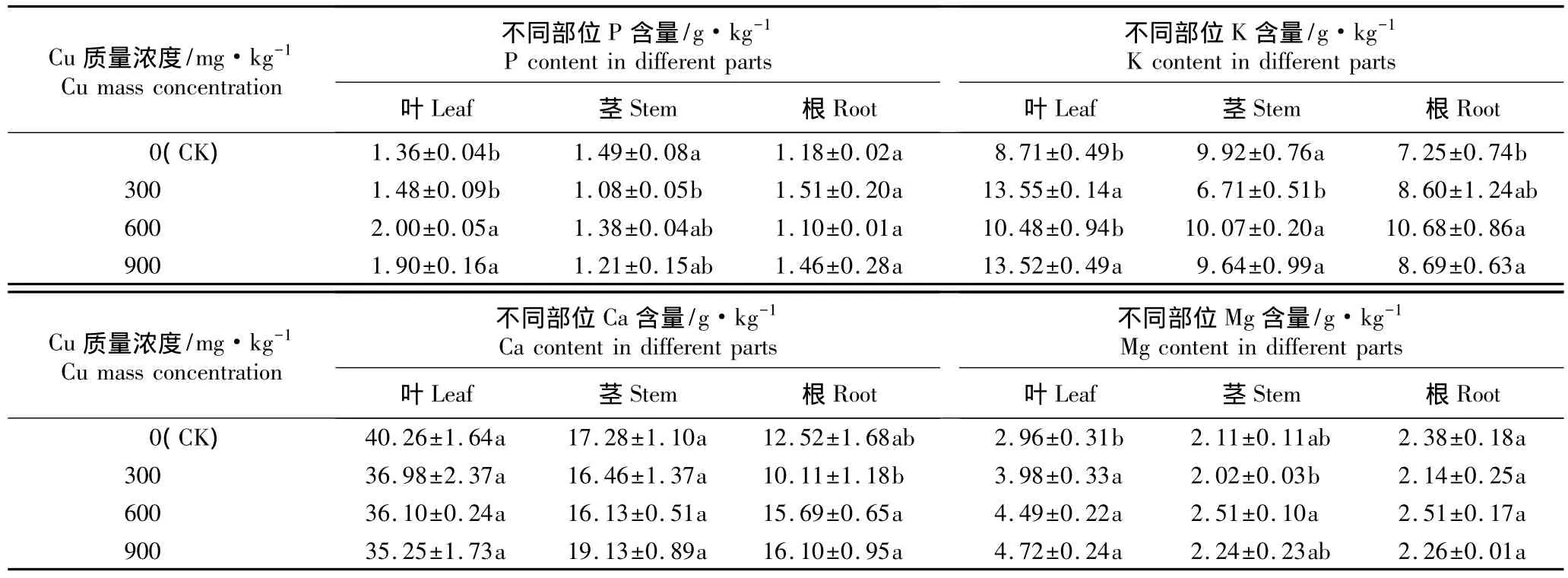
表4 土壤Cu胁迫对牡丹品种‘肉芙蓉’5年生嫁接苗不同部位大量元素含量的影响(±SE)1)Table 4 Effect of Cu stress in soil on macro element content in different parts of 5-year-old grafted seedlings of Paeonia suffruticosa‘Roufurong’(±SE)1)
1)同列中不同的小写字母表示经LSD检验差异显著(P<0.05)Different small letters in the same column indicate the significant difference by LSD test(P<0.05).
?
表5 土壤Cu胁迫对牡丹品种‘肉芙蓉’5年生嫁接苗不同部位微量元素含量的影响(±SE)1)Table 5 Effect of Cu stress in soil on micro element content in different parts of 5-year-old grafted seedlings of Paeonia suffruticosa‘Roufurong’(±SE)1)

表5 土壤Cu胁迫对牡丹品种‘肉芙蓉’5年生嫁接苗不同部位微量元素含量的影响(±SE)1)Table 5 Effect of Cu stress in soil on micro element content in different parts of 5-year-old grafted seedlings of Paeonia suffruticosa‘Roufurong’(±SE)1)
1)同列中不同的小写字母表示经LSD检验差异显著(P<0.05)Different small letters in the same column indicate the significant difference by LSD test(P<0.05).
?
经不同质量浓度Cu处理后叶片中Ca含量均低于对照但差异均不显著;经300和600 mg·kg-1Cu处理后茎中的Ca含量低于对照,而经900 mg·kg-1Cu处理后茎中的Ca含量则高于对照,但差异均不显著;经300 mg·kg-1Cu处理后根中的Ca含量低于对照,而经600和900 mg·kg-1Cu处理后根中的Ca含量则均高于对照,但差异均不显著。
经不同质量浓度Cu胁迫处理后牡丹嫁接苗叶片中的Mg含量均高于对照且与对照差异显著;茎和根中的Mg含量或高于对照或低于对照,但与对照均无显著差异。
2.3.2 对微量元素含量的影响 由表5可知:经不同质量浓度Cu处理后叶片中的Cu含量均低于对照但无显著差异,茎和根中的Cu含量均高于对照,其中,600和900 mg·kg-1Cu处理组茎中的Cu含量与对照差异显著;而各处理组根中的Cu含量均大幅度高于叶和茎,且与对照均有显著差异,并随Cu质量浓度的提高而逐渐增加。由此可见,牡丹品种‘肉芙蓉’嫁接苗根部是吸收Cu的主要器官,能使其地上部分的Cu含量维持在较低水平,从而减轻Cu对植株地上部分的毒害作用。
由表5还可见:经不同质量浓度Cu处理后嫁接苗叶、茎和根中的Fe含量或高于对照或低于对照,但与对照均无显著差异,说明在土壤Cu胁迫条件下嫁接苗各部位对Fe的吸收均未受到明显影响。经不同质量浓度Cu处理后嫁接苗叶和根中的Mn含量或高于对照或低于对照,但差异均不显著;而茎中Mn含量均高于对照,其中仅300 mg·kg-1Cu处理组茎中的Mn含量与对照有显著差异。经不同质量浓度Cu处理后嫁接苗叶、茎和根中的Zn含量均低于对照,但叶片中Zn含量与对照差异显著,茎中Zn含量与对照无显著差异,仅900 mg·kg-1Cu处理组根中Zn含量显著低于对照。
2.4 土壤Cu胁迫条件下牡丹嫁接苗的Cu富集及转运能力分析
经不同质量浓度Cu胁迫处理后牡丹品种‘肉芙蓉’5年生嫁接苗不同部位的Cu富集系数以及不同部位Cu转运系数见表6。
表6 土壤Cu胁迫条件下牡丹品种‘肉芙蓉’5年生嫁接苗不同部位的Cu富集及转运系数(±SE)1)Table 6 Cu enrichment and translocation coefficients in different parts of 5-year-old grafted seedlings of Paeonia suffruticosa‘Roufurong’under Cu stress in soil(±SE)1)
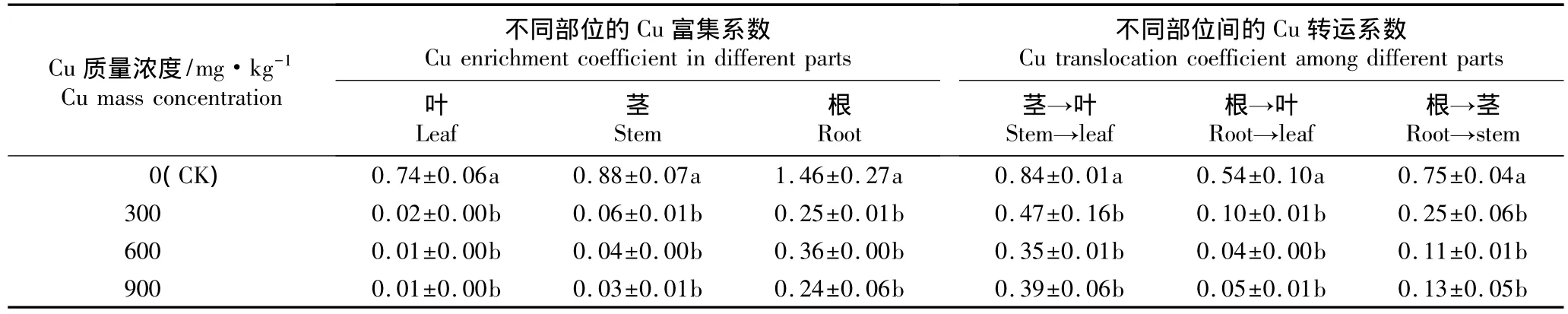
表6 土壤Cu胁迫条件下牡丹品种‘肉芙蓉’5年生嫁接苗不同部位的Cu富集及转运系数(±SE)1)Table 6 Cu enrichment and translocation coefficients in different parts of 5-year-old grafted seedlings of Paeonia suffruticosa‘Roufurong’under Cu stress in soil(±SE)1)
1)同列中不同的小写字母表示经LSD检验差异显著(P<0.05)Different small letters in the same column indicate the significant difference by LSD test(P<0.05).
?
富集系数(enrichment coefficient)能够反映植物对重金属的富集能力,富集系数越高,表明植物对该重金属元素的吸收能力越强[16]。由表6可以看出:在对照和不同质量浓度Cu胁迫条件下,牡丹嫁接苗根的Cu富集系数均最大、叶的Cu富集系数均最小,说明在土壤Cu胁迫条件下根系对Cu的吸收能力最强。在不同质量浓度Cu胁迫条件下嫁接苗各部位的Cu富集系数均显著低于对照,但各处理组间无显著差异,这一现象可能与各处理组盆栽基质中Cu质量浓度过高有关。
转运系数(translocation coefficient)能够反映出植物从下部组织向上部组织运输重金属的能力,也能体现重金属在植物组织中的分布状况。由表6还可见:在对照和不同质量浓度Cu胁迫条件下,牡丹嫁接苗由茎至叶的Cu转运系数均最大,由根至叶的Cu转运系数均最小;且在不同质量浓度Cu胁迫条件下,嫁接苗体内由茎至叶、由根至叶和由根至茎的Cu转运系数均显著低于对照,且各处理组间无显著差异。说明在土壤Cu胁迫条件下,牡丹品种‘肉芙蓉’嫁接苗能够将体内的Cu积累在下部组织中,并能有效阻止Cu向上部组织转运,从而减轻Cu对植株地上部分的毒害作用。
3 讨论和结论
在重金属胁迫条件下,一些植物能够将大部分重金属离子积累在根部,以减轻重金属对植株地上部分的毒害作用[17-18]。本研究中,经不同质量浓度Cu胁迫处理后牡丹品种‘肉芙蓉’5年生嫁接苗体内Cu主要积累在根部,且根中Cu含量随土壤中Cu质量浓度的提高显著增加,据此可认为牡丹品种‘肉芙蓉’嫁接苗属于根部囤积型植物。
田胜尼等[19]的研究结果表明:铜尾矿对豆科(Fabaceae)植物主根的伸长及侧根的发生具有抑制作用,本研究也得出类似的结果,即土壤中高质量浓度Cu胁迫能够抑制牡丹品种‘肉芙蓉’嫁接苗主根的伸长生长,在土壤Cu胁迫条件下一级侧根数量也显著减少。由于根系直接与Cu胁迫环境接触,且积累在根部的Cu离子对细胞分裂有一定的负作用,尤其在高质量浓度Cu胁迫条件下根系中过多的Cu导致毒害作用进一步加深,对主根的生长和侧根的形成产生明显的抑制作用。
在本研究中,不同质量浓度Cu胁迫处理对牡丹品种‘肉芙蓉’嫁接苗的新枝长度无显著影响,仅高质量浓度(900 mg·kg-1)Cu处理对叶面积扩展及比叶质量增加有显著的抑制作用;不同质量浓度Cu胁迫对叶片叶绿素a、叶绿素b及总叶绿素含量均无显著影响,但对叶绿素a/b比值有较大影响,其中经中、高质量浓度(600和900 mg·kg-1)Cu胁迫处理后叶绿素a/b比值均较对照显著下降。根据“叶片衰老时叶绿素a/b比值降低[20]”这一规律,可见经中、高质量浓度Cu胁迫处理后牡丹品种‘肉芙蓉’嫁接苗叶片提前进入衰老进程,且高质量浓度Cu胁迫处理对牡丹嫁接苗植株的光合作用及营养物质的积累有明显的抑制作用。
重金属胁迫可导致植物体内大量活性氧自由基的产生,从而对细胞膜造成伤害;而植物自身能够启动保护系统来清除体内的活性氧自由基,以减轻胁迫环境对机体的伤害[21]。在中、低质量浓度(600和300 mg·kg-1)Cu胁迫条件下,牡丹品种‘肉芙蓉’嫁接苗的SOD活性均较对照显著增加,而在高质量浓度Cu胁迫条件下SOD活性并未表现出较中、低质量浓度Cu胁迫条件下显著降低的现象,表明在土壤Cu胁迫条件下牡丹品种‘肉芙蓉’嫁接苗启动体内的SOD系统以清除体内产生的活性氧自由基,且其SOD系统对Cu胁迫具有很强的耐受能力。在本研究中,牡丹品种‘肉芙蓉’嫁接苗叶片中的POD活性在低质量浓度Cu胁迫条件下显著升高,但在中、高质量浓度Cu胁迫条件下POD活性却明显降低,表明其POD系统对Cu胁迫较为敏感。在低质量浓度Cu胁迫条件下,嫁接苗体内的MDA含量低于对照且无显著差异,但在中、高质量浓度Cu胁迫条件下MDA含量均较对照增加,说明在低质量浓度Cu胁迫条件下,牡丹品种‘肉芙蓉’嫁接苗体内的抗氧化酶系统能有效清除体内产生的活性氧,而中、高质量浓度Cu胁迫对嫁接苗的伤害程度则超出了抗氧化酶系统的防御限度,致使细胞膜结构遭受一定程度的破坏,膜脂过氧化程度增加,表现为体内的MDA含量增加。
Graham等[22]的研究结果显示:随Cu胁迫浓度的提高,植物对其他矿质元素的吸收能力降低。但在本实验中,土壤中添加不同质量浓度的Cu对根系中的P、Ca、Mg、Fe和 Mn 含量均无显著影响,可见牡丹品种‘肉芙蓉’嫁接苗植株能够有效阻止这些矿质元素的流失。赵艳等[23]认为55~80 mg·L-1Cu处理可以促进根系对K的吸收。本研究也得出类似结果,即在低质量浓度Cu胁迫条件下,牡丹品种‘肉芙蓉’嫁接苗根系中的K含量无显著变化,且一定质量浓度的Cu处理还能促进嫁接苗根系对K的吸收,推测这可能是植物的一种自我保护反应,通过吸收K+来提高细胞内阳离子的浓度,从而缓解土壤Cu胁迫对植物细胞的毒害作用。在 Cu胁迫条件下,紫鸭跖草(Setcreasea purpurea Boom)根系中 Zn 含量下降[24]。本研究也得出类似的结果,即在不同质量浓度Cu胁迫条件下牡丹品种‘肉芙蓉’嫁接苗叶、茎和根中的Zn含量均低于对照。
本研究结果表明:在不同质量浓度Cu胁迫条件下,牡丹品种‘肉芙蓉’嫁接苗叶片中除Zn含量外,其他矿质元素的含量总体上未显著降低,其中,叶片中的P含量在中、高质量浓度Cu胁迫条件下显著增加,K含量在一定质量浓度Cu胁迫条件下显著增加,且Mg含量也均显著增加。说明在土壤Cu胁迫条件下茎作为矿质元素的运输通道发挥了极为重要的作用,保证了叶片对矿质元素的需求,使土壤Cu胁迫条件下牡丹品种‘肉芙蓉’嫁接苗体内大部分矿质元素的代谢仍能正常进行。
孙雨亮等[25]的研究结果表明:在不同浓度Cu胁迫条件下溪荪(Iris sanguinea Donn ex Horn.)对Cu的富集系数和转运系数均显著低于对照且各处理组间差异显著。本研究也得出类似的研究结果,即在不同质量浓度Cu胁迫条件下牡丹品种‘肉芙蓉’嫁接苗根、茎和叶的Cu富集系数以及由茎到叶、由根到茎、由根到叶的Cu转运系数均显著低于对照,但各处理组间差异不显著。推测Cu富集系数的显著降低可能与盆栽土中Cu添加量过高、远超出牡丹嫁接苗的吸收能力有关;而Cu转运系数的显著下降则说明牡丹嫁接苗能将Cu积累在下部组织中,有效阻止Cu向上部组织的转运,从而减轻土壤Cu胁迫对植株上部组织的伤害。
综合分析结果表明:以凤丹实生苗为砧木获得的牡丹品种‘肉芙蓉’嫁接苗对土壤Cu胁迫具备一定的耐性,在低质量浓度Cu胁迫条件下植株生长良好,可用于轻度Cu污染土壤的植物修复。但是,关于Cu胁迫对其开花品质的影响则仍有待进一步研究。
[1]郭 霞,张桂荣,赵孝庆,等.凤丹实生苗嫁接观赏牡丹技术[J].山东林业科技,1999(3):47-48.
[2]薛 杰,侯西明,申玉美,等.实用牡丹繁殖技术[J].林业实用技术,2003(1):44-45.
[3]刘雁丽,史 萍,夏 妍,等.Cu和丹皮酚磺酸钠处理对凤丹根系生长、丹皮酚含量及H+-ATPase活性的影响[J].植物资源与环境学报,2012,21(1):20-27.
[4]张 敏,江建华,徐 洁.铜矿尾矿库复垦种植牡丹可行性研究[J].上海环境科学,2000,19(12):585-587.
[5]周 燕,张桂花,徐迎春,等.铜对凤丹生长、生理及体内矿质元素吸收分配的影响[J].生态学杂志,2011,30(3):415-423.
[6]沈章军,王友保,王广林,等.铜陵铜尾矿凤丹种植基地重金属污染初探[J].应用生态学报,2005,16(4):673-677.
[7]ROUPHAEL Y,CARDARELLI M,REA E,et al.Grafting of cucumber as a means to minimize copper toxicity[J].Environmental and Experimental Botany,2008,63(1/3):49-58.
[8]张自坤,刘世琦,刘素慧,等.嫁接对铜胁迫下黄瓜幼苗根系多胺代谢的影响[J].应用生态学报,2010,21(8):2051-2056.
[9]张自坤,刘作新,张 颖,等.铜胁迫对嫁接和自根黄瓜幼苗光合作用及营养元素吸收的影响[J].中国生态农业学报,2009,17(1):135-139.
[10]李合生.植物生理生化实验原理和技术[M].北京:高等教育出版社,2000:260-263.
[11]BEAUCHAMP C,FRIDOVICH I.Superoxide dismutase:improved assays and an assay applicable to acrylamide gels[J].Analytical Biochemistry,1971,44(1):276-287.
[12]ZHOU W,ZHAO D,LIN X.Effects of waterlogging on nitrogen accumulation and alleviation of waterlogging damage by application of nitrogen fertilizer and mixtalol in winter rape (Brassica napus L.)[J].Journal of Plant Growth Regulation,1997,16(1):47-53.
[13]MUÑOZ-MUÑOZ J L,GARCÍA-MOLINA F,GARCÍA-RUIZ P A,et al.Enzymatic and chemical oxidation of trihydroxylated phenols[J].Food Chemistry,2009,113(2):435-444.
[14]QUINTANILLA-GUERRERO F, DUARTE-VÁZQUEZ M A,GARCÍA-ALMENDAREZ B E,et al.Polyethylene glycol improves phenol removal by immobilized turnip peroxidase[J].Bioresource Technology,2008,99(18):8605-8611.
[15]ARNON D L.Copper enzymes in isolated chloroplasts.Polyphenoloxidase in Beta vulgaris[J].Plant Physiology,1949,24(1):1-15.
[16]夏汉平,束文圣.香根草和百喜草对铅锌尾矿重金属的抗性与吸收差异研究[J].生态学报,2001,21(7):1122-1129.
[17]施 翔,陈益泰,王树凤,等.3种木本植物在铅锌和铜矿砂中的生长及对重金属的吸收[J].生态学报,2011,31(7):1818-1826.
[18]李 影,王友保.4种蕨类草本植物对Cu的吸收和耐性研究[J].草业学报,2010,19(3):191-197.
[19]田胜尼,刘登义,王峥峰,等.铜尾矿对5种豆科植物根系生长的影响[J].生态环境,2005,14(2):199-203.
[20]BISWAS A K,CHOUDHURI M A.Mechanism of monocarpic senescence in rice[J].Plant Physiology,1980,65(2):340-345.
[21]周希琴,莫灿坤.植物重金属胁迫及其抗氧化系统[J].新疆教育学院学报,2003,19(2):103-108.
[22]GRAHAM J H,TIMMER L W,FARDELMANN D.Toxicity of fungicidal copper in soil to citrus seedlings and vesicular-arbuscular mycorrhizal fungi[J].Phytopathology,1986,76(1):66-70.
[23]赵 艳,徐迎春,吴晓丽,等.Cu胁迫对狭叶香蒲体内元素吸收分配的影响[J].生态学杂志,2009,28(4):665-670.
[24]黄长干,陈 赟,邱业先.铜盐毒害对紫鸭跖草养分吸收和生长的影响[J].中国生态农业学报,2008,16(1):168-172.
[25]孙雨亮,黄苏珍,原海燕.Cu污染土壤中溪荪和花菖蒲的生长状况及对Cu的积累及转运能力[J].植物资源与环境学报,2011,20(2):49-55.
