典型森林和草地生态系统呼吸各组分间的相互关系
2013-12-09朱先进于贵瑞王秋凤高艳妮赵新全韩士杰闫俊华
朱先进,于贵瑞,王秋凤,高艳妮,赵新全,韩士杰,闫俊华
(1. 中国科学院地理科学与资源研究所生态网络观测与模拟重点实验室CERN综合研究中心, 北京 100101; 2. 中国科学院大学, 北京 100049; 3. 中国科学院西北高原生物研究所, 西宁 810001; 4. 中国科学院沈阳应用生态研究所, 沈阳 110016; 5. 中国科学院华南植物园, 广州 510650)
典型森林和草地生态系统呼吸各组分间的相互关系
朱先进1,2,于贵瑞1,*,王秋凤1,高艳妮1,2,赵新全3,韩士杰4,闫俊华5
(1. 中国科学院地理科学与资源研究所生态网络观测与模拟重点实验室CERN综合研究中心, 北京 100101; 2. 中国科学院大学, 北京 100049; 3. 中国科学院西北高原生物研究所, 西宁 810001; 4. 中国科学院沈阳应用生态研究所, 沈阳 110016; 5. 中国科学院华南植物园, 广州 510650)
生态系统呼吸是陆地生态系统碳收支的重要组成部分,分析其组分间的相互关系对理解生态系统呼吸过程和精确评价生态系统碳收支具有重要意义,也是当前碳循环研究工作的一大难点。利用ChinaFLUX的长白山温带针阔混交林(CBS),鼎湖山亚热带常绿阔叶林(DHS)和海北灌丛草甸(HBGC)3个典型生态系统的通量观测数据,分析了经验统计方法在中国典型生态系统中的适用性及敏感性,揭示了生态系统呼吸各组分的动态变化特征及相互关系。结果表明:采用呼吸组分拆分方法所获结果与理论推测及实测数据大致相同,拆分结果对净初级生产力与总初级生产力比值(NPP/GPP)的变化较为敏感,NPP/GPP变化0.1时,自养呼吸在生态系统呼吸中的比例(Ra/RE)改变0.05。各生态系统中,生态系统呼吸及其组分在年内均表现出明显的单峰型变化特征,在夏季生长旺盛的时节达到最大值。异养呼吸与生态系统呼吸的比值(Rh/RE)也具有明显的季节变化,但在生态系统间表现出明显差异,CBS和HBGC分别表现出先增大后减小和先减小后增大的变化趋势,DHS则相对稳定,在0.5附近波动, Ra/RE的季节动态与Rh/RE相反。在年总量上,HBGC主要通过异养呼吸向大气排放CO2,异养呼吸占生态系统呼吸的60%,而CBS和DHS的自养呼吸和异养呼吸所占比重大致相似,异养呼吸占生态系统呼吸的49%。这说明,经验统计学模型可以用来进行生态系统呼吸组分的拆分,进而可以为生态系统碳循环过程的精细研究提供参考数据,但今后应加强NPP/GPP的测定,以提高生态系统呼吸拆分的精度。
陆地生态系统;生态系统呼吸;涡度相关;碳通量;自养呼吸;异养呼吸
生态系统呼吸(RE)是碳循环的重要分量[1],与气候变化间存在着正反馈效应[2- 3],不仅在当前全球碳收支中占有较大的比重,而且在未来还会呈现不断增大的趋势[4]。已有研究表明,全球的RE约为103 PgC a-1[5],占全球总初级生产力(GPP)总量(123 PgC a-1[6])的83.74%。
按照呼吸底物的不同,RE可分为自养呼吸(Ra)和异养呼吸(Rh)。其中,Ra是植物维持自身生命活动所必需的新陈代谢过程,又分为维持呼吸和生长呼吸。维持呼吸与温度相关,随着温度的增加呈指数增加[7];生长呼吸与温度没有直接关系,但与GPP呈一定的比例[8]。Rh是残存有机质分解并向大气释放CO2的过程[9],主要受土壤温湿度和有机质含量的影响[10]。因而,影响RE组分的因素存在差异,拆分RE的各个组分、分析各组分间的相互关系有助于深入认识生态系统碳循环的过程机理,也可为陆地生态系统碳收支的精确评估提供理论依据。同时,Ra的变化对大气CO2浓度具有重要影响[11],科学界迫切需要拆分和评估RE的各组分,为评价Ra对大气CO2浓度变化的贡献提供数据。
目前,拆分RE组分的方法有许多种。第1种是将箱式法与涡度相关系统观测相结合,分别获取土壤呼吸(Rs)和RE,实现RE组分的拆分[12- 14]。然而,基于箱式法获取的Rs包含了Ra和Rh,对Rs进行拆分也是一项艰巨的工作。第2种是利用生物量调查法获取生态系统净初级生产力(NPP),结合涡度相关观测结果估算Ra和Rh[15- 17],但生物量调查和涡度相关观测的时间尺度不一致,并且生物量调查法耗费大量的人力物力,难以获得持续的、高质量观测数据。第3种是基于过程机理模型模拟Ra和Rh,但该方法所需参数较多,结果也受到模型设计者对RE过程理解程度的限制[10]。基于同位素分馏比也可以实现对RE组分的拆分[18- 19],但所需仪器设备非常昂贵。Schwalm等[20]提出了基于通量观测数据的统计学拆分方法,并对其不确定性进行了分析。结果表明,该方法具有很好的稳定性,不同参数方案间没有明显差异,这为利用通量观测数据分析RE组分提供了新的技术途径。
中国陆地生态系统通量观测网络(ChinaFLUX)自2002年建立以来,已积累了大量的观测数据,构建了学术界普遍认可的通量数据处理流程[21]。然而,现有通量数据处理仅将生态系统碳通量分解为GPP和RE,尚没有拆分RE的组分。因此,本研究利用Schwalm等[20]分析方法,拆分并评价我国典型森林和草地生态系统的RE组分及其相互关系,进而阐明:(1)不同生态系统Ra和Rh的动态特征,(2)Rh/RE的年内动态变化特征,(3)不同生态系统间Ra和Rh年总量的差异。该研究可以为生态系统碳循环过程研究提供基础数据,也为通量数据处理过程中RE拆分方法的确定提供依据。
1 研究地点与研究方法
1.1 站点介绍
本研究选择ChinaFLUX中3个陆地生态系统为研究对象,分别为:长白山温带针阔混交林(CBS)、鼎湖山亚热带常绿阔叶林(DHS)和海北高寒灌丛草甸(HBGC),各生态系统的基本信息如表1所示。
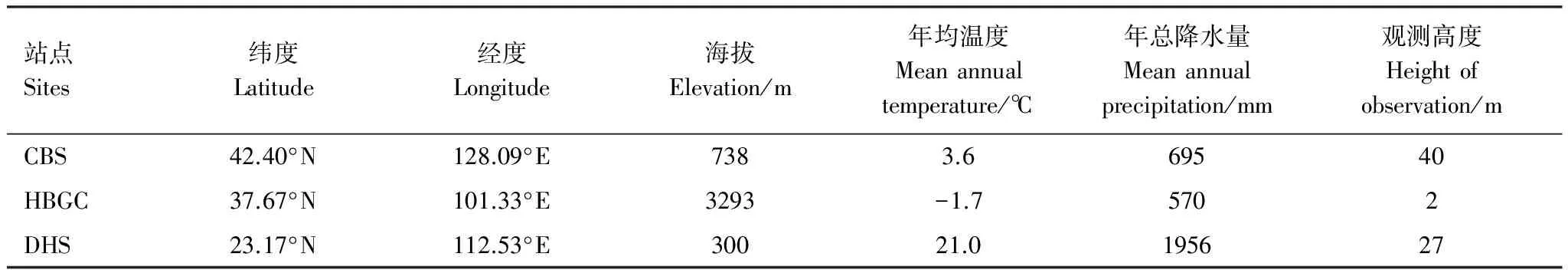
表1 研究站点基本信息
CBS:长白山温带针阔混交林Changbai Mountains temperate conifer and broadleaf mixed forest;HBGC:海北灌丛草甸Haibei alpine Shrub;DHS:鼎湖山亚热带常绿阔叶林Dinghu Mountains subtropical evergreen broadleaf forest
3个生态系统均利用开路式涡度相关系统(OPEC)进行生态系统碳水通量观测,原始采样频率为10 Hz,通过数据采集器计算并储存30 min的通量数据。同时观测温度、湿度、降水、辐射等气象要素并计算30 min平均值储存[21- 24]。此外,辅以静态箱-气相色谱法测定土壤呼吸,每隔7—10 d测定1次[25- 27]。
1.2 数据处理
1.2.1 通量数据处理
采用ChinaFLUX通用数据处理流程对30 min通量数据进行处理[21],包括三次坐标旋转、WPL校正、储存项计算及降水剔除、阈值剔除和方差剔除等,并根据Reichstein等[28]的方法确定夜间u*临界值、实现夜间数据剔除、完成数据质量控制。利用非线性回归方法对碳通量数据进行插补,并将碳通量拆分为GPP和RE。
1.2.2 自养呼吸和异养呼吸的拆分
本研究根据自养呼吸与生态系统呼吸的比值(Ra/RE)对RE进行拆分[20]。具体做法为:假设净生态系统生产力(NEP)与RE的比值为θ,Ra/RE为β,则:

(1)
因而,根据式(1)可以得到β,即:

(2)
已有研究表明,NPP/GPP介于0.47—0.60之间[20],因而,β的范围可以表示为:
(3)
在生态学研究中,Schwalm等[20]将Ra/RE或Rh/RE的最小值Ω设为0.10,并分析了Ω上下浮动0.05后的结果,他发现基于不同Ω得到的β没有明显差异,因而此处Ω设为0.10。
当β取最小值(0.10)时,θ=-0.75;当β取最大值(0.90)即Rh/RE取最小值(0.10)时,θ=0.70。因而,θlt;-0.75或者θgt;0.70时,公式(3)不再适用。当θ=-0.75时,βlower为0.10,βupper为0.13;当θ=0.70时,βlower为0.68,βupper为0.90。针对不同的θ值,设定不同的β范围,具体范围如表2所示。
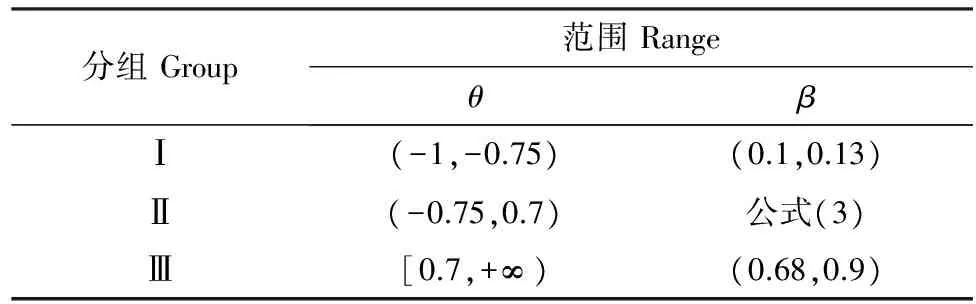
表2 不同θ值下的β范围
根据θ值,在β给定的范围内随机选取1000个数据,取其平均值为β。夜间时,θ=-1,利用公式(3)无法获取β值,因而将当日白天最后一个有效的β和次日最早一个有效的β进行线性内插的方法获取夜间β。
最后,根据β值和RE计算Ra和Rh,即:
Ra=β×RE
Rh=(1-β)×RE
(4)
1.3 生态系统呼吸拆分结果的验证
为了验证该方法的合理性,利用静态箱-气相色谱法观测的土壤呼吸(Rs)数据与本研究获取的Rh进行对比。由于静态箱-气相色谱法的观测时间是9:00—11:00,因此,Rh选用9:00—11:30的平均值。
1.4 数据统计
采用单因素方差分析法评价不同生态系统间RE组分的差异,采用线性回归分析拆分结果与静态箱-气相色谱法所获结果的差异。所有数据统计分析过程均在matlab7.7中进行。
2 结果与分析
2.1 生态系统呼吸及其组分的季节动态
生态系统碳通量具有明显的季节变化特征[29- 31]。由于年际间生态系统的季节动态特征相似[32- 34],本文以2003年数据为例分析中国典型森林和草地生态系统碳通量的季节动态(图1)。

图1 2003年典型生态系统气温及碳通量的季节变化特征Fig.1 Seasonal dynamics of air temperature and carbon fluxes in typical ecosystems in 2003
气温(Ta)在3个典型生态系统均表现出单峰型变化(图1),最大值出现在7月份前后,但不同生态系统,其年内气温的变化幅度存在明显差异,DHS常年保持较高的气温,北方两个生态系统(CBS和HBGC)年内气温变幅较大。不同生态系统,GPP的季节动态有所差异(图1),北方生态系统(CBS和HBGC)表现出明显的单峰型特点,但DHS的GPP在秋冬季节较高,在6—8月因温度过高而略低,这与已有结果[23]相一致。NEP的季节动态与GPP相似(图1),但峰值出现时间较GPP略有提前。
3个生态系统的RE均表现出明显的单峰型变化,峰值均出现在7—8月份(图1),与气温的季节动态(图1)相似,这是因为,生态系统尺度上,气温的波动决定了RE的季节动态[35]。生态系统间,最大呼吸速率存在差异,CBS可以达到9.20 gC m-2d-1,DHS和HBGC明显小于CBS,均不足4 gC m-2d-1。这主要是生产力的差异引起的,生产力与RE存在着密切的耦合关系[35- 37]。
与RE的动态特征相似,Ra和Rh也表现出单峰型的季节动态(图1),峰值出现的时间也与RE相似,峰值大小约为RE峰值的一半。
2.2 生态系统呼吸组分与生态系统呼吸比值(Rh/RE和Ra/RE)的季节动态
异养呼吸与生态系统呼吸的比值(Rh/RE)表现出明显的季节波动,但在生态系统间表现出不同的变化特征(图2)。
Rh/RE在CBS表现为年初和年末略低、生长季开始和结束时略有增加的趋势,在生长旺盛时节大致维持在0.5左右,DHS全年均在0.5附近波动。与两个森林生态系统的变化趋势不同,Rh/RE在HBGC表现出明显的单峰型动态,非生长季可以达到0.9,随着生长季的开始,Rh/RE逐渐降低,并在生长旺季达到0.5左右的最小值,随着生长季的结束,异养呼吸占据主导,Rh/RE又开始增大。
自养呼吸与生态系统呼吸的比值(Ra/RE)表现出与Rh/RE相反的季节变化趋势。

图2 2003年三个典型生态系统的生态系统呼吸组分在生态系统呼吸中所占比例(Rh/RE, Ra/RE)的季节动态特征Fig.2 Seasonal dynamics of the ratio of autotrophic respiration and heterotrophic respiration to ecosystem respiration (Ra/RE, Rh/RE) in typical ecosystems in 2003
2.3 生态系统呼吸及其组分的年总量
生态系统呼吸的组分反映了生态系统消耗碳量的不同途径。中国3个典型生态系统的RE及其组分的年总量如表3所示。
生态系统间,RE、Ra和Rh均存在显著差异,均表现为CBS最高,DHS居中,HBGC最低。生态系统间Rh/RE的差异表现为:两个森林生态系统(CBS和DHS)间没有明显差别,约为0.49,但显著小于HBGC的0.60。Ra/RE表现出相反的特征,即HBGC的数值为0.40,明显小于CBS和DHS的0.51。
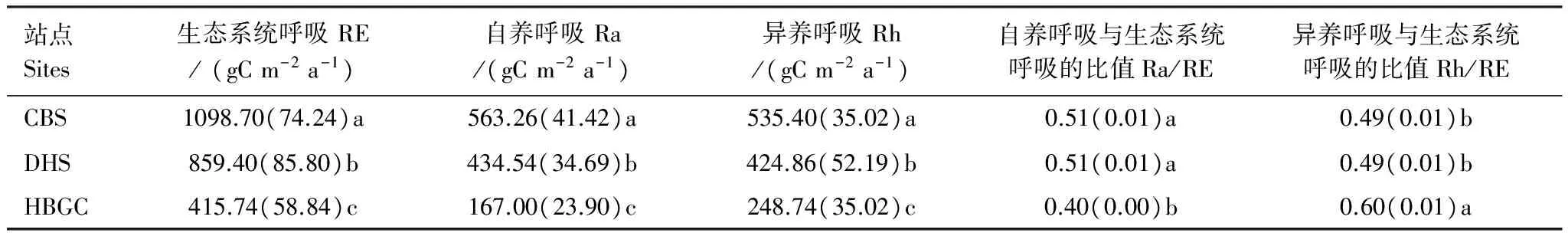
表3 典型生态系统的生态系统呼吸及其组分的年总量
表中数据为2003—2005年的平均值,括号内数值为2003—2005年的标准差
3 讨论
3.1 生态系统呼吸拆分结果的不确定性
本研究采用的方法是Schwalm等[20]对加拿大森林RE进行研究时提出的,目前尚没有在其他生态系统中应用。但相对于其他研究方法,本方法结构简单、可操作性强并具有一定的生态学理论基础。
理论上来说,当NPP/GPP保持恒定的比值时,Ra/RE随着NEP/RE的增加而增大,即随着NEP/RE的增加,RE中的Rh组分降低,从图2可知,Ra/RE的变化与NEP的变化有相似的规律,这证实了该方法在理论上是可行的。
在理论分析的基础上,用静态箱-气相色谱法观测的Rs与本研究中拆分得到的Rh作对比(图3),以验证该方法的拆分效果。
从图3可以看出,在CBS,Rh与Rs的比值为0.34(图3),这在之前研究结果[38- 39]的范围内,并与东北林区其他研究[40]相似。在DHS和HBGC,Rh与Rs间没有明显的关系。这是因为,在CBS,利用箱式法观测的Rs是生态系统呼吸的一部分(图3),可以反映RE的动态,但在DHS和HBGC,箱式法观测到的Rs几乎与该时段的RE相一致,这可能是由箱式法本身观测的误差引起的。
进一步整理了已发表文献的NPP数据,结合涡度相关观测的碳通量结果,估算RE及其组分年总量间的相互关系。在CBS,张娜等[41]模拟发现,1995年该生态系统Ra/RE高达0.82,高于本研究结果(表3),这可能是他们估算的NEP过高及NPP偏小导致的,NEP值高达392 gC m-2a-1[41],比涡度相关观测结果[34]高54%。在DHS,2003—2004年的NPP为970 gC m-2a-1[42],结合Yu等[34]的结果可以看到,这两年该生态系统的Ra/RE为52.33%,与本研究结果(表3)相似。
虽然Piao等[11]总结已有研究结果[1, 43- 44]发现,Ra与GPP的比值虽然表现出随着温度的增加先降低后增大,但具有较大的变异性,基于本研究的方法,CBS、DHS、HBGC的Ra/GPP分别为0.40、0.31、0.35,也在Piao等[11]的范围之内。
因而,本研究所用方法具有简便易行、无需额外测定即可准确拆分RE,这为分析碳收支组分及其影响因素提供了可能,可以在生态系统碳循环研究中应用。
3.2 生态系统呼吸拆分结果对NPP/GPP的敏感性
本研究将NPP/GPP的值设为0.47—0.60,但Litton等[43]整理文献数据发现,NPP/GPP变异很大,最小甚至可能低于0.20。因而有必要进一步分析NPP/GPP的变化对RE组分拆分的影响。
此处设定两个方案,即将NPP/GPP(θ)的上下限(0.47—0.60)分别下调0.10(方案I)和上调0.10(方案II),相应的RE/NEP(β)的临界比值发生改变(表4)。
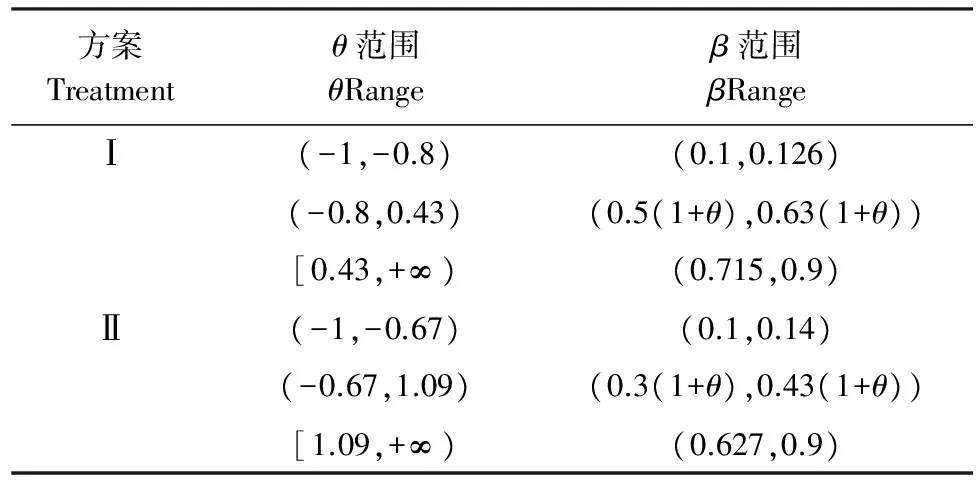
表4 不同处理方案下θ值下的β范围
基于不同数据处理方案获得了生态系统呼吸组分相互关系的季节动态(图4),Ra/RE及Rh/RE的季节动态与图2没有明显差异,说明不同NPP/GPP的变化不会对Ra/RE及Rh/RE的季节动态产生明显影响。
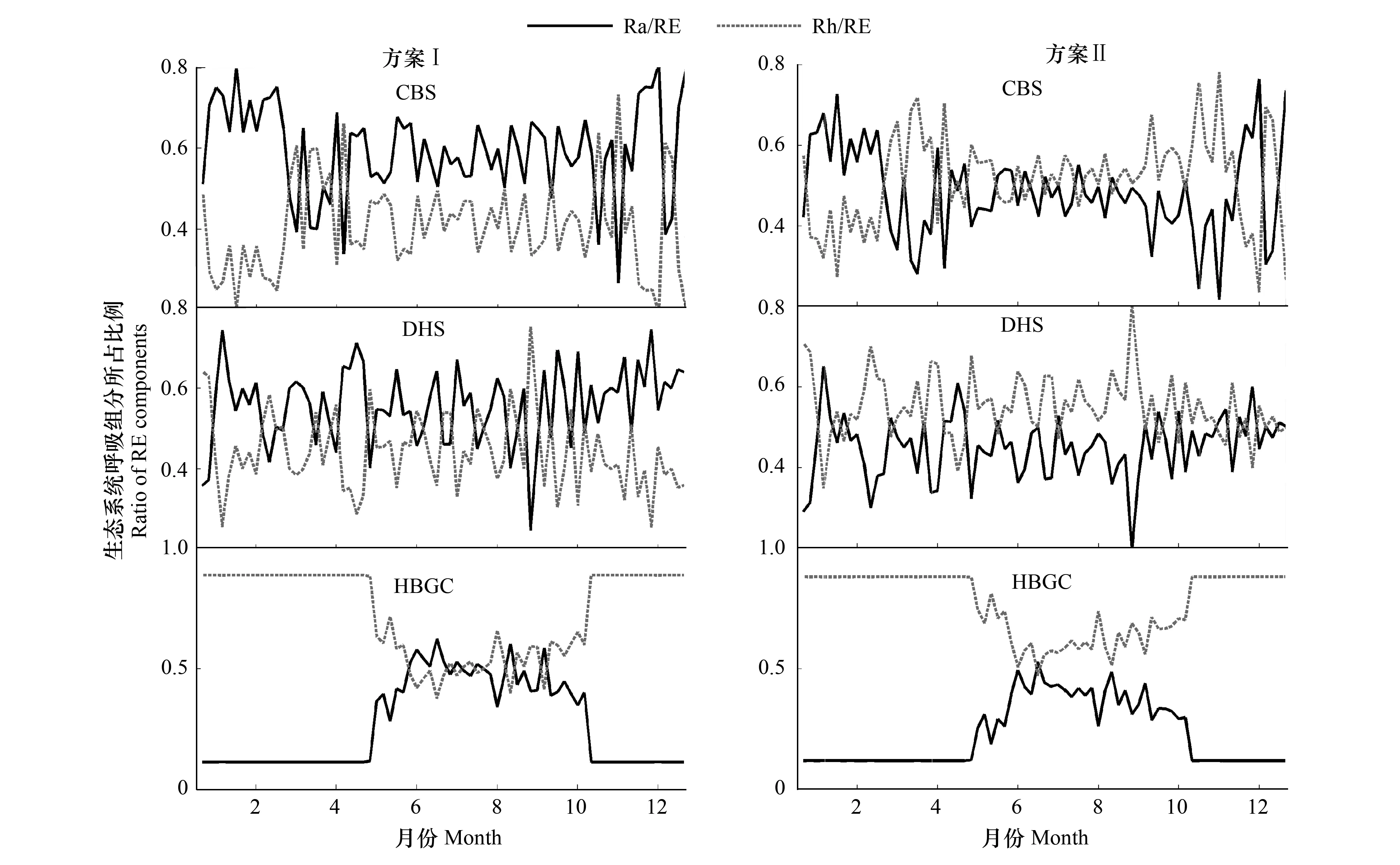
图4 2003年不同NPP/GPP的典型生态系统的生态系统呼吸组分在生态系统呼吸中所占比例(Rh/RE, Ra/RE)的季节动态特征Fig.4 Seasonal dynamics of the ratio of autotrophic respiration and heterotrophic respiration to ecosystem respiration (Ra/RE, Rh/RE) in typical ecosystems in 2003 with different NPP/GPP
不同数据处理方案下,生态系统呼吸组分在生态系统呼吸中所占的比例发生一定的波动(表5),NPP/GPP每改变0.1,Ra/RE变化0.05,Ra/RE随着NPP/GPP的增加而降低。
表5不同数据处理方案生态系统呼吸组分在生态系统呼吸中所占比例
Table5Theratioofautotrophicrespirationandheterotrophicrespirationtoecosystemrespiration(Ra/RE, Rh/RE)underdifferentdatatreatments
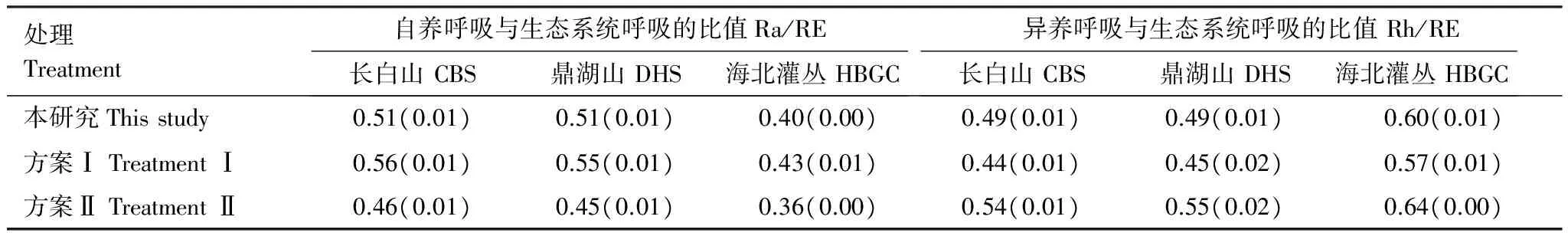
处理Treatment自养呼吸与生态系统呼吸的比值Ra/RE长白山CBS鼎湖山DHS海北灌丛HBGC异养呼吸与生态系统呼吸的比值Rh/RE长白山CBS鼎湖山DHS海北灌丛HBGC本研究Thisstudy0.51(0.01)0.51(0.01)0.40(0.00)0.49(0.01)0.49(0.01)0.60(0.01)方案ⅠTreatmentⅠ0.56(0.01)0.55(0.01)0.43(0.01)0.44(0.01)0.45(0.02)0.57(0.01)方案ⅡTreatmentⅡ0.46(0.01)0.45(0.01)0.36(0.00)0.54(0.01)0.55(0.02)0.64(0.00)
表中数据为2003—2005年的平均值,括号内数值为观测期间的标准差
生态系统间,不管NPP/GPP发生何种变化,HBGC的Ra/RE均小于CBS和DHS,并且该值始终小于0.5,因而HBGC的生态系统呼吸以异养呼吸为主。随着NPP/GPP的变化,CBS和DHS中Ra/RE在0.5上下浮动,表明森林生态系统中自养和异养呼吸所占的比例大致相似。
可见,NPP/GPP影响着RE组分拆分,生物量调查获取的NPP将为生态系统碳通量组分的拆分提供重要基础数据。
3.3 生态系统呼吸各组分间的相互关系
在季节动态上,Ra/RE随着NEP的增加有增大的趋势[20],表明随着植物生长的开始,生态系统在不断固碳的基础上通过自养呼吸向外排放的碳量也在增加。这是因为Ra中的维持呼吸与温度相关,随着温度的增加呈指数增加[7];生长呼吸与温度没有直接关系,但与GPP呈一定的比例关系[8];而Rh受土壤温湿度和有机质含量的影响。因此,在生长季开始后,随着GPP和气温的增大,Ra增加速度快于Rh,导致Ra/RE开始增加。
生态系统之间,Ra年总量与RE年总量的比值也明显不同,森林生态系统明显高于草地生态系统,表明森林生态系统Ra所占比例高于草地生态系统,与Schwalm等[20]的结果相同。这是因为,森林生态系统具有较高的NEP,NEP/RE也较高,导致Ra/RE高。比较中国与加拿大森林生态系统间[20]的Ra/RE的差异可以发现,中国森林生态系统的Ra/RE明显高于加拿大森林生态系统,这也是由中国森林生态系统中NEP/RE高于加拿大所决定的。
4 结论
基于ChinaFLUX3个陆地生态系统(CBS:长白山温带混交林,DHS:鼎湖山亚热带常绿阔叶林,HBGC:海北高寒灌丛草甸)的涡度相关观测数据,利用统计分析方法拆分了RE组分,分析了该方法在中国陆地生态系统的适用性及对NPP/GPP的敏感性,阐明了RE组分的动态特征及其相互关系。结果表明:基于涡度相关观测数据及NPP/GPP的比值,可以准确拆分中国典型生态系统的RE,但该方法对NPP/GPP较为敏感。3个生态系统中,RE及其组分年内均表现出单峰型的变化,在生长旺盛时节(7月份)达到最大值。异养呼吸在生态系统呼吸中所占的比例(Rh/RE)表现出明显的季节变化,CBS表现出先增大后减小的季节特征,DHS则常年维持为0.5左右,HBGC表现出先降低后增大的特点。年总量上,Rh/RE因生态系统的不同而不同,CBS和DHS两个森林生态系统的自养和异养呼吸大致相当,HBGC的碳排放则主要通过异养呼吸来实现。该方法可以拆分RE组分,进而为生态系统碳循环的过程研究提供参考数据,但今后应加强NPP/GPP的测定,以提高生态系统呼吸拆分的精度。
Referrences:
[1] Luyssaert S, Inglima I, Jung M, Richardson A D, Reichsteins M, Papale D, Piao S L, Schulzes E D, Wingate L, Matteucci G, Aragao L, Aubinet M, Beers C, Bernhoffer C, Black K G, Bonal D, Bonnefond J M, Chambers J, Ciais P, Cook B, Davis K J, Dolman A J, Gielen B, Goulden M, Grace J, Granier A, Grelle A, Griffis T, Grunwald T, Guidolotti G, Hanson P J, Harding R, Hollinger D Y, Hutyra L R, Kolar P, Kruijt B, Kutsch W, Lagergren F, Laurila T, Law B E, Le Maire G, Lindroth A, Loustau D, Malhi Y, Mateus J, Migliavacca M, Misson L, Montagnani L, Moncrieff J, Moors E, Munger J W, Nikinmaa E, Ollinger S V, Pita G, Rebmann C, Roupsard O, Saigusa N, Sanz M J, Seufert G, Sierra C, Smith M L, Tang J, Valentini R, Vesala T, Janssens I A. CO2balance of boreal, temperate, and tropical forests derived from a global database. Global Change Biology, 2007, 13(12): 2509- 2537.
[2] Davidson E A, Janssens I A, Luo Y Q. On the variability of respiration in terrestrial ecosystems: moving beyond Q10. Global Change Biology, 2006, 12(2): 154- 164.
[3] Houghton R A, Davidson E A, Woodwell G M. Missing sinks, feedbacks, and understanding the role of terrestrial ecosystems in the global carbon balance. Global Biogeochemical Cycles, 1998, 12(1): 25- 34.
[4] Cox P M, Betts R A, Jones C D, Spall S A, Totterdell I J. Acceleration of global warming due to carbon-cycle feedbacks in a coupled climate model. Nature, 2000, 408(6809): 184- 187.
[5] Yuan W P, Luo Y Q, Li X L, Liu S G, Yu G R, Zhou T, Bahn M, Black A, Desai A R, Cescatti A, Marcolla B, Jacobs C, Chen J Q, Aurela M, Bernhofer C, Gielen B, Bohrer G, Cook D R, Dragoni D, Dunn A L, Gianelle D, Grünwald T, Ibrom A, Leclerc M Y, Lindroth A, Liu H P, Marchesini L B, Montagnani L, Pita G, Rodeghiero M, Rodrigues A, Starr G, Stoy P C. Redefinition and global estimation of basal ecosystem respiration rate. Global Biogeochemical Cycles, 2011, 25: GB4002, doi: 10.1029/2011gb004150.
[6] Beer C, Reichstein M, Tomelleri E, Ciais P, Jung M, Carvalhais N, Rodenbeck C, Arain M A, Baldocchi D, Bonan G B, Bondeau A, Cescatti A, Lasslop G, Lindroth A, Lomas M, Luyssaert S, Margolis H, Oleson K W, Roupsard O, Veenendaal E, Viovy N, Williams C, Woodward F I, Papale D. Terrestrial gross carbon dioxide uptake: global distribution and covariation with climate. Science, 2010, 329(5993): 834- 838.
[7] Ryan M G. A simple method for estimating gross carbon budgets for vegetation in forest ecosystems. Tree Physiology, 1991, 9(1/2): 255- 266.
[8] Chen J M, Liu J, Cihlar J, Goulden M L. Daily canopy photosynthesis model through temporal and spatial scaling for remote sensing applications. Ecological Modelling, 1999, 124(2/3): 99- 119.
[9] Trumbore S. Carbon respired by terrestrial ecosystems-recent progress and challenges. Global Change Biology, 2006, 12(2): 141- 153.
[10] Yu G R. Scientific Frontier on Human Activities and Ecosystem Changes. Beijing: Higher Education Press, 2009.
[11] Piao S L, Luyssaert S, Ciais P, Janssens I A, Chen A P, Cao C, Fang J Y, Friedlingstein P, Luo Y Q, Wang S P. Forest annual carbon cost: a global-scale analysis of autotrophic respiration. Ecology, 2010, 91(3): 652- 661.
[12] Irvine J, Law B E, Martin J G, Vickers D. Interannual variation in soil CO2efflux and the response of root respiration to climate and canopy gas exchange in mature ponderosa pine. Global Change Biology, 2008, 14(12): 2848- 2859.
[13] Nagy Z, Pintér K, Pavelka M, Darenová E, Balogh J. Carbon fluxes of surfaces vs. ecosystems: advantages of measuring eddy covariance and soil respiration simultaneously in dry grassland ecosystems. Biogeosciences, 2011, 8(9): 2523- 2534.
[14] Griffis T J, Black T A, Gaumont-Guay D, Drewitt G B, Nesic Z, Barr A G, Morgenstern K, Kljun N. Seasonal variation and partitioning of ecosystem respiration in a southern boreal aspen forest. Agricultural and Forest Meteorology, 2004, 125(3/4): 207- 223.
[15] Law B E, Thornton P E, Irvine J, Anthoni P M, Van Tuyl S. Carbon storage and fluxes in ponderosa pine forests at different developmental stages. Global Change Biology, 2001, 7(7): 755- 777.
[16] Luyssaert S, Reichstein M, Schulze E D, Janssens I A, Law B E, Papale D, Dragoni D, Goulden M L, Granier A, Kutsch W L, Linder S, Matteucci G, Moors E, Munger J W, Pilegaard K, Saunders M, Falge E M. Toward a consistency cross-check of eddy covariance flux-based and biometric estimates of ecosystem carbon balance. Global Biogeochemical Cycles, 2009, 23: GB3009, doi: 10.1029/2008gb003377.
[17] Tan Z H, Zhang Y P, Yu G R, Sha L Q, Tang J W, Deng X B, Song Q H. Carbon balance of a primary tropical seasonal rain forest. Journal of Geophysical Research, 2010, 115: D00H26, doi: 10.1029/2009jd012913.
[18] Riveros-Iregui D A, Hu J, Burns S P, Bowling D R, Monson R K. An interannual assessment of the relationship between the stable carbon isotopic composition of ecosystem respiration and climate in a high-elevation subalpine forest. Journal of Geophysical Research, 2011, 116: G02005, doi: 10.1029/2010jg001556.
[19] Schuur E A G, Trumbore S E. Partitioning sources of soil respiration in boreal black spruce forest using radiocarbon. Global Change Biology, 2006, 12(2): 165- 176.
[20] Schwalm C R, Black T A, Morgenstern K, Humphreys E R. A method for deriving net primary productivity and component respiratory fluxes from tower-based eddy covariance data: a case study using a 17-year data record from a Douglas-fir chronosequence. Global Change Biology, 2007, 13(2): 370- 385.
[21] Yu G R, Fu Y L, Sun X M, Wen X F, Zhang L M. Recent progress and future directions of ChinaFLUX. Science in China Series D: Earth Sciences, 2006, 49(Suppl II): 1- 23.
[22] Zhang J H, Yu G R, Han S J, Guan D X, Sun X M. Seasonal and annual variation of CO2flux above a broad-leaved Korean pine mixed forest. Science in China Series D: Earth Sciences, 2006, 49(Suppl II): 63- 73.
[23] Wang C L, Yu G R, Zhou G Y, Yan J H, Zhang L M, Wang X, Tang X L, Sun X M. CO2flux evaluation over the evergreen coniferous and broad-leaved mixed forest in Dinghushan, China. Science in China Series D: Earth Sciences, 2006, 49(Suppl II): 127- 138.
[24] Li Y N, Sun X M, Zhao X Q, Zhao L, Xu S X, Gu S, Zhang G F, Yu G R. Seasonal variations and mechanism for environmental control of NEE of CO2concerning thePotentillafruticosain alpine shrub meadow of Qinghai-Tibet Plateau. Science in China Series D: Earth Sciences, 2006, 49(Suppl II): 174- 185.
[25] Zheng Z M, Yu G R, Sun X M, Cao G M, Wang Y S, Du M Y, Li J, Li Y N. Comparison of eddy covariance and static chamber/gas chromatogram methods in measuring ecosystem respiration. Chinese Journal of Applied Ecology, 2008, 19(2): 290- 298.
[26] Lin L S, Han S J, Wang Y S, Gu Z J. Soil CO2flux in several typical forests of Mt. Changbai. Chinese Journal of Ecology, 2004, 23(5): 42- 45.
[27] Zhang D Q, Sun X M, Zhou G Y, Yan J H, Wang Y S, Liu S Z, Zhou C Y, Liu J X, Tang X L, Li J, Zhang Q M. Seasonal dynamics of soil CO2effluxes with responses to environmental factors in lower subtropical forests of China. Science in China Series D: Earth Sciences, 2006, 49(Suppl II): 139- 149.
[28] Reichstein M, Falge E, Baldocchi D, Papale D, Aubinet M, Berbigier P, Bernhofer C, Buchmann N, Gilmanov T, Granier A, Grunwald T, Havrankova K, Ilvesniemi H, Janous D, Knohl A, Laurila T, Lohila A, Loustau D, Matteucci G, Meyers T, Miglietta F, Ourcival J M, Pumpanen J, Rambal S, Rotenberg E, Sanz M, Tenhunen J, Seufert G, Vaccari F, Vesala T, Yakir D, Valentini R. On the separation of net ecosystem exchange into assimilation and ecosystem respiration: review and improved algorithm. Global Change Biology, 2005, 11(9): 1424- 1439.
[29] Law B E, Williams M, Anthoni P M, Baldocchi D D, Unsworth M H. Measuring and modelling seasonal variation of carbon dioxide and water vapour exchange of aPinusponderosaforest subject to soil water deficit. Global Change Biology, 2000, 6(6): 613- 630.
[30] Valentini R, Gamon J A, Field C B. Ecosystem ags-exchange in a California grassland-seasonal patterns and implications for scaling. Ecology, 1995, 76(6): 1940- 1952.
[31] Guan D X, Wu J B, Zhao X S, Han S J, Yu G R, Sun X M, Jin C J. CO2fluxes over an old, temperate mixed forest in northeastern China. Agricultural and Forest Meteorology, 2006, 137(3/4): 138- 149.
[32] Barr A G, Black T A, Hogg E H, Griffis T J, Morgenstern K, Kljun N, Theede A, Nesic Z. Climatic controls on the carbon and water balances of a boreal aspen forest, 1994—2003. Global Change Biology, 2007, 13(3): 561- 576.
[33] Polley H W, Frank A B, Sanabria J, Phillips R L. Interannual variability in carbon dioxide fluxes and flux-climate relationships on grazed and ungrazed northern mixed-grass prairie. Global Change Biology, 2008, 14(7): 1620- 1632.
[34] Yu G R, Zhang L M, Sun X M, Fu Y L, Wen X F, Wang Q F, Li S G, Ren C Y, Song X, Liu Y F, Han S J, Yan J H. Environmental controls over carbon exchange of three forest ecosystems in eastern China. Global Change Biology, 2008, 14(11): 2555- 2571.
[35] Wen X F, Wang H M, Wang J L, Yu G R, Sun X M. Ecosystem carbon exchanges of a subtropical evergreen coniferous plantation subjected to seasonal drought, 2003—2007. Biogeosciences, 2010, 7(1): 357- 369.
[36] Wang X C, Wang C K, Yu G R. Spatio-temporal patterns of forest carbon dioxide exchange based on global eddy covariance measurements. Science in China Series D: Earth Sciences, 2008, 51(8): 1129- 1143.
[37] Lasslop G, Reichstein M, Detto M, Richardson A D, Baldocchi D D. Comment on Vickers et al.: self-correlation between assimilation and respiration resulting from flux partitioning of eddy-covariance CO2fluxes. Agricultural and Forest Meteorology, 2010, 150(2): 312- 314.
[38] Subke J A, Inglima I, Francesca Cotrufo M. Trends and methodological impacts in soil CO2efflux partitioning: a metaanalytical review. Global Change Biology, 2006, 12(6): 921- 943.
[39] Zhan X Y, Yu G R, Zheng Z M, Wang Q F. Carbon emission and spatial pattern of soil respiration of terrestrial ecosystems in China: based on geostatistic estimation of flux measurement. Progress in Geography, 2012, 31(1): 97- 108.
[40] Wang C K, Yang J Y. Rhizospheric and heterotrophic components of soil respiration in six Chinese temperate forests. Global Change Biology, 2007, 13(1): 123- 131.
[41] Zhang N, Yu G R, Zhao S D, Yu Z L. Carbon budget of ecosystem in Changbai Mountain natural reserve. Environmental Science, 2003, 24(1): 24- 32.
[42] Yan J H, Wang Y P, Zhou G Y, Zhang D Q. Estimates of soil respiration and net primary production of three forests at different succession stages in South China. Global Change Biology, 2006, 12(5): 810- 821.
[43] Litton C M, Raich J W, Ryan M G. Carbon allocation in forest ecosystems. Global Change Biology, 2007, 13(10): 2089- 2109.
[44] DeLucia E H, Drake J E, Thomas R B, Gonzalez-Meler M. Forest carbon use efficiency: is respiration a constant fraction of gross primary production?. Global Change Biology, 2007, 13(6): 1157- 1167.
[10] 于贵瑞. 人类活动与生态系统变化的前沿科学问题. 北京: 高等教育出版社, 2009.
[25] 郑泽梅, 于贵瑞, 孙晓敏, 曹广民, 王跃思, 杜明远, 李俊, 李英年. 涡度相关法和静态箱/气相色谱法在生态系统呼吸观测中的比较. 应用生态学报, 2008, 19(2): 290- 298.
[26] 林丽莎, 韩士杰, 王跃思, 谷志静. 长白山四种林分土壤CO2释放通量的研究. 生态学杂志, 2004, 23(5): 42- 45.
[39] 展小云, 于贵瑞, 郑泽梅, 王秋凤. 中国区域陆地生态系统土壤呼吸的碳排放及空间格局——基于通量观测的地学统计评估. 地理科学进展, 2012, 31(1): 97- 108.
[41] 张娜, 于贵瑞, 赵士洞, 于振良. 长白山自然保护区生态系统碳平衡研究. 环境科学, 2003, 24(1): 24- 32.
Theinteractionbetweencomponentsofecosystemrespirationintypicalforestandgrasslandecosystems
ZHU Xianjin1,2, YU Guirui1, *, WANG Qiufeng1, GAO Yanni1,2, ZHAO Xinquan3, HAN Shijie4, YAN Junhua5
1SynthesisResearchCenterofCERN,KeyLaboratoryofEcosystemNetworkObservationandModeling,InstituteofGeographicSciencesandNaturalResourcesResearch,ChineseAcademyofSciences,Beijing100101,China2UniversityofChineseAcademyofSciences,Beijing100049,China3NorthwestPlateauInstituteofBiology,ChineseAcademyofSciences,Xining810001,China4InstituteofAppliedEcology,ChineseAcademyofSciences,Shenyang110016,China5SouthChinaBotanicalGarden,ChineseAcademyofSciences,Guangzhou510650,China
Ecosystem respiration (RE), the release of carbon from an ecosystem into the atmosphere, is a key component of the terrestrial ecosystem carbon budget and plays an important role in the global carbon balance. Analyzing the interaction between RE components is essential to understand RE and to accurately evaluate the ecosystem carbon budget. There are many methods for separating RE into autotrophic respiration (Ra) and heterotrophic respiration (Rh), but each approach has disadvantages. Large RE data have obtained through long-term eddy covariance measurements, while the interaction between Ra and Rh is poorly documented, which inhibits the accurate assessment of global carbon budget. In this study, we used an empirical statistical method to separate RE into its two components and to examine component relationships and seasonal dynamics at three ChinaFLUX sites: 1) Changbaishan temperate mixed forest (CBS); 2) Dinghushan subtropical evergreen broad-leaf forest (DHS); and 3) Haibei shrub meadow (HBGC). The applicability and sensitivity of this method in typical ecosystems of China were also evaluated. The method used in this study was based on the ratio of Ra to RE (Ra/RE). The range of Ra/RE was obtained by calculating two ratios: the ratio of RE to net ecosystem productivity (NEP) (RE/NEP) and that of net primary productivity (NPP) to gross primary productivity (GPP) (NPP/GPP). Within the range of Ra/RE, 1000 Ra/REs were randomly selected and the value of Ra/RE used in this study was set as the mean of the 1000 random Ra/REs. Ra and Rh were then calculated using Ra/RE and RE.
Our study showed that the RE separating method produced consistent results with those obtained through static chamber/gas chromatographic techniques at the same sites, as well as with biomass surveys and theoretical speculation. The interaction of RE components was sensitive to the variation of NPP/GPP: a ten-percent increase of NPP/GPP led to a five-percent decrease of Ra/RE. In all three ecosystems, RE and its components showed similar seasonal dynamics, with a single-peak pattern achieving its maximum midway through the growing season. The ratio of Rh to RE (Rh/RE) also showed different seasonal dynamic among the three ecosystems. In CBS, Rh/RE increased during the first half of the year, reached its peak during the growing season then decreased. However, Rh/RE in HBGC decreased during the first half of the year and increased again later in growing-season. In DHS, Rh/RE was relatively stable at approximately 0.5. Seasonal dynamics of Ra/RE were opposite to those in Rh/RE. The annual total Rh accounted for 60% of the RE in HBGC, suggesting that a large proportion of emitted carbon was released by Rh in this ecosystem. In CBS and DHS, Rh was only 49% of RE, indicating that the release of carbon through Ra and Rh was nearly the same in these two forest ecosystems. Results indicate that this statistical method, which requires detailed observations of NPP/GPP, can successfully separate RE into Rh and Ra and can provide necessary data for the detailed analysis of the ecosystem carbon cycle.
terrestrial ecosystem; ecosystem respiration; eddy covariance; carbon fluxes; autotrophic respiration; heterotrophic respiration
国家重点基础研究发展规划资助项目(2010CB833504); 国家自然科学基金资助项目(31061140359, 30590380); 中国科学院战略性先导科技专项资助项目(XDA05050601)
2012- 07- 13;
2013- 06- 21
*通讯作者Corresponding author.E-mail: yugr@igsnrr.ac.cn
10.5846/stxb201207130988
朱先进,于贵瑞,王秋凤,高艳妮,赵新全,韩士杰,闫俊华.典型森林和草地生态系统呼吸各组分间的相互关系.生态学报,2013,33(21):6925- 6934.
Zhu X J, Yu G R, Wang Q F, Gao Y N, Zhao X Q, Han S J, Yan J H.The interaction between components of ecosystem respiration in typical forest and grassland ecosystems.Acta Ecologica Sinica,2013,33(21):6925- 6934.
