海南铜鼓岭灌木林稀疏规律
2013-12-09杨小波龙文兴李东海卜广发
周 威,龙 成,杨小波,*,龙文兴,李东海,卜广发
(1. 海南大学热带作物种质资源保护与开发利用教育部重点实验室, 海口 570228;2. 海南大学园艺园林学院, 海口 570228)
海南铜鼓岭灌木林稀疏规律
周 威1,2,龙 成1,2,杨小波1,2,*,龙文兴1,2,李东海1,2,卜广发1,2
(1. 海南大学热带作物种质资源保护与开发利用教育部重点实验室, 海口 570228;2. 海南大学园艺园林学院, 海口 570228)
以海南省文昌市铜鼓岭国家级自然保护区160m×160m的山麓灌木林固定样地中胸径(DBH)≥1.5cm的所有木本植物为对象,并根据DBH划分为7个径级,研究其稀疏规律,结果表明:1)群落中Ⅰ级(DBHlt;4.5cm)个体数所占百分比最大,占64%,群落结构为“倒J型”,林分密度是幼树gt;小树gt;成年树,该群落正处于稳定状态,且个体间为争夺更多的生存空间和资源发生强烈的自疏和它疏作用,存在明显的稀疏现象。2)各径级地上生物量随着密度增加呈幂函数增加,幂函数方程为:AGB=6×107N0.4626或lnAGB=0.4626lnN+17.855,在较小的密度范围内,随林分密度的增加,群落地上总生物量增加较快,但当密度趋于0.6株/m2时,地上生物量变化缓慢,趋于恒定值。3)林分密度与各径级平均胸径呈负相关关系(密度越大,平均胸径越小),其幂函数关系式为:N=70.1d-3.5506,R2=0.8808。4)选择Yoda提出的幂函数方程对天然灌木林自然稀疏规律进行模拟,林分密度与平均生物量之间具有显著相关性,其关系式为:W=2219.1N-0.5374或lnW=7.7048-0.5374lnN;稀疏指数α值为0.5374,这与Yoda所提出的3/2指数相差甚远,并不满足-3/2自疏定律。5)此次调查的物种有常见种(0.2hm2样地中个体数≥5的种为常见种)41种,非常见种53种。天然灌木林在物种层面的稀疏也存在一定的规律。
灌木林群落;稀疏;林分密度;地上生物量
物种丰富、结构复杂的植物群落中种群数量变化受两个因素控制:一是种群密度的压力导致的种内竞争,二是不同种群争夺有限资源而导致的种间竞争。植物种内竞争导致自疏现象,种间竞争导致它疏现象[1]。对于植物种群而言,随初始种群密度的增加和个体的生长而密集,并因种群内个体之间为争夺各生态因子(如光、水、营养等)而发生竞争,导致部分个体死亡,即自疏[2]。它是群落具有的自我调节机能之一,通过密度调控把个体适应、种群数量动态、生物群落演替与生态系统稳定性联系起来,一直是生态学领域研究的热点问题[3- 4]。林分密度能引起植株个体间因生长资源的强制分配而产生相互作用,密度的增加导致植物种内竞争,使种群中单株生长量和生物量发生改变[5- 6],从而影响植物个体的异速生长模式。
生态学家和林业工作者对于植物种群所特有的自疏现象进行了大量的理论研究,尤以Yoda基于植物生长是几何相似的理论提出的反映同龄单一植物种群存活密度和平均个体重量之间关系的-3/2自疏法则被广泛接受,其表达式为:W=CN-3/2(其中W为平均个体质量,N为种群密度,C为常数)[7]。该理论被很多实验数据所支持[8- 12],到目前为止,已有100多种植物种群,包括人工种植种群和天然种群、较小的草本植物种群和高大乔木种群的自疏斜率均接近-3/2[13]。有一些研究表明在形态和生长型具有明显差异的两种植物混合种群中,个体平均生物量与密度间的异速关系斜率也很接近-3/2[14- 15];甚至有些生态学家认为-3/2幂指数法则适用于多物种的混合种群[16]。例如周永斌等以白石砬子自然保护区14个为多种类的混合种群,最优树种均未超过3成的天然林为研究对象,利用近30年间14次的连续定位观测数据进行自疏研究,发现其中9个天然林的自疏指数约为1.5,满足-3/2法则[17]。天然林植物群落存在种内竞争,同时也存在它疏现象,其植物的密度-生物量关系是否满足-3/2?目前没有研究结论。本文将自疏与它疏看成是一种综合性的“自疏”作用,这种“自疏”就是群落的稀疏,是二者共同作用的结果。本文将采用类似于自疏的研究方法来研究这种稀疏关系,因为无论是自疏还是它疏,都会反应在密度与生物量之间的关系上。本文选取海南沿海的文昌铜鼓岭国家级自然保护区的天然灌木林为研究对象,探讨它的年龄结构、林分密度与地上生物量、平均胸径及平均生物量的异速生长关系,推导稀疏指数,以期得出热带沿海灌木林的稀疏规律,为天然林经营管理提供理论依据。
1 材料和方法
1.1 研究地区概况
铜鼓岭自然保护区位于文昌市东部,东濒南海,西连内陆。地理位置110°58′30″—111°03′00″E,19°36′54″—19°41′21″N,属热带海洋性季风气候,雨水充沛、干湿季节分明、日照充足,平均气温23.9℃,年均降雨量1721.6 mm,年平均蒸发量1872.7mm,月平均风速多在8—10m/s之间。地质构造几乎全由海西——印支期似斑状黑云母花岗岩和燕山中粒黑云母花岗岩构成。山体沿海岸线自西北向东南延伸,构成海岸的天然屏障。土壤以砖红壤为主,土层浅薄。植被类型有山麓灌木林和热带常绿季雨矮林,灌木林从海拔15m到山顶(330m)均有分布,而季雨矮林则主要分布在山顶。本研究样地共记录了木本植物94种,隶属32科,74属,种类较多的有大戟科(9属10种)、茜草科(8属8种)、芸香科(6属8种)、无患子科(6属7种),其中乔木68种,灌木26种。主要以贡甲(Maclurodendronoligophlebium)、海南大风子(Hydnocarpushainanensis)、柄果木(Mischocarpussundaicus)、猪肚木(Canthiumhorridum)、多脉紫金牛(Ardisiacrassinervosa)、无患子(Sapindussaponaria)等为优势种。整个群落的落叶树种占到15%,如常见的有猪肚木(C.horridum)、无患子(S.saponaria)、黄牛木(Cratoxylumcochinchinense)、美叶菜豆树(Radermacherafrondosa)、算盘子(Glochidionpuberum)、菲岛算盘子(Glochidionphilippicum)、白背算盘子(Glochidionwrightii)、毛茶(Antirheachinensis)、山石榴(Catunaregamspinosa)等。尽管该群落出现了较多的乔木树种,有向该地区顶级群落季雨矮林演替的趋势,但目前还是属于灌木林,因为灌木树种个体数达到了70%,群落外观表现的也是灌木林。
1.2 调查方法
2011年在保护区内建成160×160m山麓灌木林固定样地,采用每木调查法调查胸径(DBH)≥1.5cm的所有木本植物的种名、胸径、高度、枝下高、冠幅、坐标。
1.3 径级划分
采用曲仲湘采用空间代替时间的方法[18],根据径级划分为不同林龄。将此次调查的胸径数据划分为7个径级,Ⅰ级:DBHlt;4.5cm;Ⅱ级:4.5cm≤DBHlt;7.5cm;Ⅲ级:7.5cm≤DBHlt;10.5cm;Ⅳ级:10.5cm≤DBHlt;13.5cm;Ⅴ级:13.5cm≤DBHlt;16.5cm;Ⅵ级:16.5cm≤DBHlt;19.5cm;Ⅶ:19.5cm≤DBH。
林分密度的计算公式为:N=n/SN为林分密度(株/m2)、n为个体数(株)、S为固定样地面积(m2)。
1.4 自疏模型
选择Yoda和Reineke等提出的自然稀疏规律进行模拟:W=CN-α,式中W为平均个体质量(g),N为种群密度(株/m2),C、α为常数。
1.5 生物量模型
采用J. Chave等[19]提出的干旱森林类型的地上生物量计算模型:
AGB = exp(-2.187+ 0.916×ln(ρD2H)) ×1000≡112×(ρD2H)0.916
J. Chave等在文中提到的森林类型是基于蒸发量、降水量和海拔来划分的,铜鼓岭地区这些指标均与干旱森林类型比较符合,且该地区旱季长达半年之久,常常出现春旱,另外土层浅薄,发育程度低,以重石质砂壤、轻壤-石质土为主,土壤的持水能力弱,再加上风速大造成水分大量蒸发,而且从植物组成来看,存在一定比例的落叶树种,更好的说明该森林类型为干旱森林,因而认为采用该模型比较好。其中AGB为植物地上生物量(g),ρ为木材密度(g/cm3),D为胸径(cm),H为树高(m)。木材密度主要从网站(http://www.worldagroforestrycentre.org/sea/products/AF-Dbases/wd/Index.htm)及《海南木材》等查阅,对于少数木材密度未能在以上数据库中找到的,将采用同属比较接近的种的木材密度。
1.6 数据分析方法
采用SAS软件和Excel软件进行统计分析
2 研究结果
2.1 群落各径级的林分密度和种群数的变化趋势
胸径小于4.5cm的幼年个体占有最大百分数(64%)(表1),而且,从表1和图1可以看出,研究样地内不同胸径级植株在群落中呈“倒J型”分布,个体平均密度都是幼树gt;小树gt;成年树,该群落正处于稳定状态。且Ⅰ级到Ⅱ、Ⅲ、Ⅳ、Ⅴ、Ⅵ、Ⅶ级的个体数、密度及种群数均迅速减少,存在明显的稀疏现象。
2.2 群落各径级地上生物量与林分密度关系
各径级植株地上生物量随着径级方向呈现减少,基本与林分密度随径级方向减少的规律一致,但是从Ⅰ级到Ⅱ级的植株地上生物量与总体趋势不一致(表1)。
各径级生物量与林分密度具有极显著正相关关系(图2)。群落各径级生物量随着密度增加呈幂函数增加,幂函数方程为:AGB=6×107N0.4626或lnAGB=0.4626lnN+17.855(R2=0.9068,Plt;0.0001)
该模型能较好说明群落各径级生物量与林分密度的关系;且当密度趋于0.6株/m2时,地上生物量变化缓慢,趋于恒定值。

表1 各径级个体数、地上生物量及种群数统计表
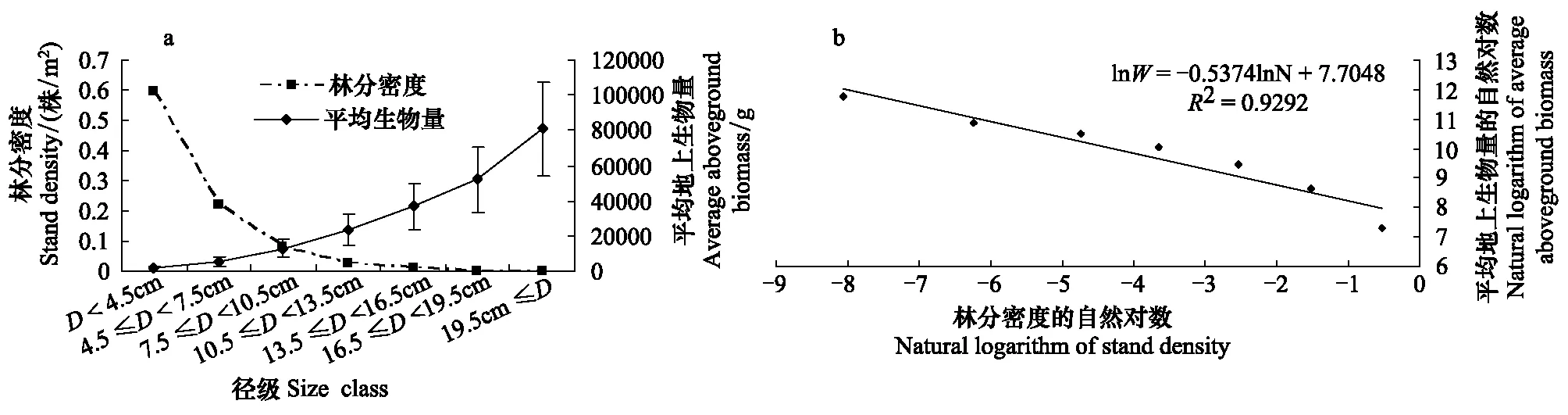
图1 灌木林自疏,各径级林分密度与平均地上生物量的回归分析Fig.1 Self-thinning of the shrubbery. The regression between the stand density and average aboveground biomass for the seven size class
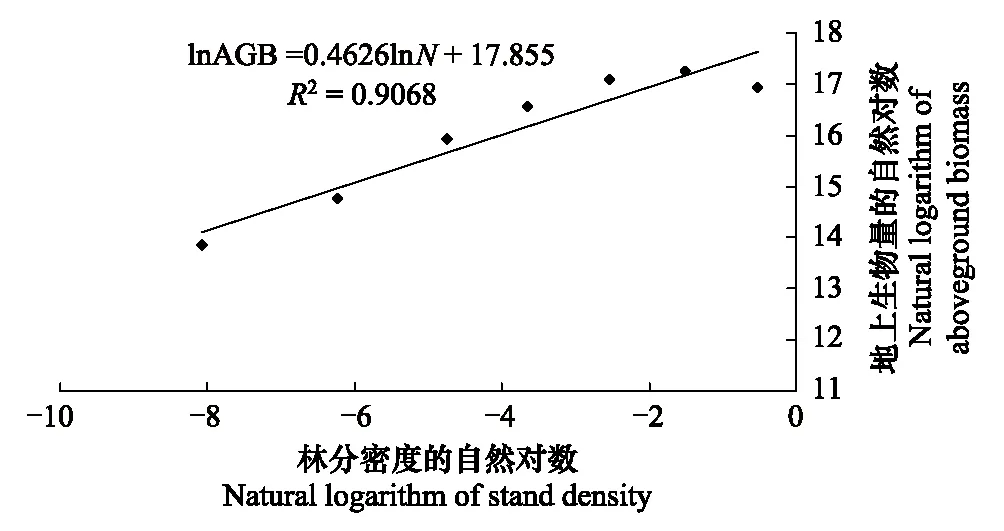
图2 各径级地上生物量与林分密度的回归分析 Fig.2 Regression between the stand density and aboveground biomass (AGB) for the seven size class
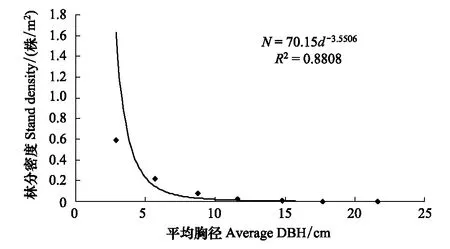
图3 平均胸径与林分密度的相关关系Fig.3 Correlativity of stand density and average DBH
2.3 平均胸径与林分密度的动态关系
群落林分密度与群落各径级平均胸径间呈显著负相关关系(图3)。林分密度与平均胸径呈幂函数关系,关系式为N=70.1d-3.5506(R2=0.8808,P=0.0017)。
2.4 各径级植株平均地上生物量与密度关系
通过图1可以看出,铜鼓岭灌木林自然稀疏现象明显,且具有连续性。随着林分密度的降低,平均胸径提高。林分密度与平均生物量之间具有显著相关性(图1)。用回归分析得到平均生物量与林分密度的关系式为:W=2219.1N-0.5374或 lnW=7.7048-0.5374lnN(R2=0.9292,r=-0.9640,P=0.0005),稀疏指数α值为0.5374。
2.5 群落各径级种群变化规律
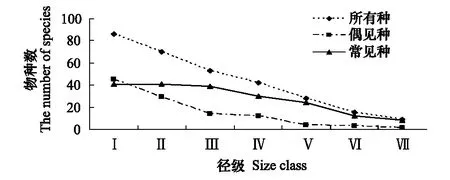
图4 各径级的物种组成图Fig.4 The chart of the species composition
由表1可以看出,种群数随径级增大而减少,但是具体相关情况需要进一步探究。为了便于分析,本研究参照Milton对物种的划分方法[20],将0.2hm2样地中个体数≥5的种作为常见种,这样就将所有植物种群分为常见种和偶见种两种类型。此次调查的物种有常见种41种,非常见种53种。
从图4中可以看出,常见种和偶见种沿随着径级增大均减少,但是常见种在Ⅰ、Ⅱ、Ⅲ级的数量基本没变化,到了Ⅲ级以后,物种数呈直线下降,减少的速度较快;而偶见种在前3个径级内物种减少的非常快,后4个径级也一直在减少,但减少幅度相比前3个径级内的减少幅度要小。因此,前3个径级所有物种数的减少主要是偶见种的减少,而后4个径级的所有种的减少是二者共同减少的结果。将偶见种和常见种的数据进行相关分析发现,二者呈显著正相关关系(R2=0.6461,P=0.0294)。进一步分析各径级的物种组成发现,Ⅰ级以后非常见种有8种新出现记录的种,分别是大青(Clerodendrumcyrtophyllum)、白桐(Claoxylonindicum)、盾柱卫矛(Pleurostyliaopposita)、海南胶核木(Myxopyrumpierrei)、鸭脚木(Scheffleraheptaphylla)、鱼尾葵(Caryotamitis)、褐叶柄果木(Mischocarpuspentapetalus)、肖蒲桃(Syzygiumacuminatissimum)。这些种绝大多数都是林下较耐阴植物。从图4也可以看出Ⅰ级径级内非常见种要比常见种多,偶见种是45种,常见种是41种。
3 讨论
3.1 天然灌木林群落结构特征
森林群落中不同径级树种的数量分布是反映群落结构稳定状态的重要指标[21]。从表1和图1可知:群落中Ⅰ级(Dlt;4.5cm)个体数所占百分比最大(64%);不同胸径级植株在群落中呈“倒J型”分布,林分密度是幼树gt;小树gt;成年树。“倒J型”径级数量分布说明森林群落正处于稳定状态。这与霸王岭、尖峰岭、五指山等地的山地雨林群落径级结构特征一致[21- 23]。不同径级植株的数量和密度差异必然与植物竞争作用有关[24]。同种个体具有同样的生长习性和资源需求, 由于资源有限, 同种邻体之间发生资源(光照、营养、空间等)竞争, 这样就可导致同种个体生长量下降、死亡率升高[25]。随着幼苗的继续生长和需光量的增加,个体间为争夺更多的生存空间和资源而必然会发生强烈的自疏和它疏作用,从而出现表1中密度迅速下降,导致年龄结构上出现I级以后个体数量急骤减少的现象。
3.2 林分密度与植株地上生物量关系
图2表明,该研究样地各径级植株地上总生物量随着密度增加呈幂函数增加,幂函数方程为:AGB=6×107N0.4626或lnAGB=0.4626lnN+17.855(R2=0.9068,Plt;0.0001);但是表1中从Ⅰ级到Ⅱ级的地上生物量是增加的,这是因为虽然Ⅱ级个体数并不比Ⅰ级个体数多,但是Ⅰ级都是小树,因而生物量会小于Ⅱ级,出现与整体趋势不一致的点。
从图2可知,各径级地上生物量随林分密度的增加而增加,在较小的密度范围内,随林分密度的增加,群落地上总生物量增加较快,但当密度趋于0.6株/m2时,地上生物量变化缓慢,当密度大于临界点密度时单位面积地上生物量呈缓慢增加直至维持在一定水平,这与单位面积蓄积量的变化规律一致,也是产量密度效应的一种体现[26- 27]。
3.3 林分密度与平均胸径关系
很多研究表明,胸径生长与密度密切相关,多呈负相关关系[28- 30],本次研究也得出了类似的结论。图1资料显示本研究的林分密度与平均胸径的幂函数关系式为:N=70.1d-3.5506(R2=0.8808,P=0.0017)。从图上和幂函数关系式可知,林分密度与群落平均胸径呈负相关关系,密度越大,平均胸径越小。这是因为高密度林分条件下,林木间竞争加剧,林木生长营养空间减小,导致林木的胸径减小;而低密度林分条件下,林木间竞争减小,林木个体拥有足够的营养空间,则林木的胸径较大。
3.4 天然林的稀疏
本文按Yoda提出的自疏模型得出的稀疏指数为-0.5376,与-3/2相去甚远,与周永斌等以白石砬子自然保护区14个天然林为研究对象所得出的结果也不一样[17]。这主要是因为天然植物群落自疏与它疏作用并存,共同作用于整个天然林群落,造成群落的稀疏。但是也并不是说天然林群落的稀疏指数不可能出现-3/2,因为群落不同时期两种作用强弱会有不同,如果当某一时期天然林自疏作用强于其他疏作用,且足够强时,也是有可能会出现满足指数接近-3/2的。至于自疏作用强弱与群落演替的时期之间的关系以及是否物种越单一,越接近纯林的天然林的稀疏指数越接近-3/2等问题可以在以后的研究中进一步去探究。另外,-3/2自疏指数本身的普适性就一直存在争议,本文得出的稀疏指数也有其存在的地位,也许是对该指数又一新证与补充。因为越来越多的研究者们发现α值并不是恒定的,它是可变的,而且其可变性比以前预测的大得多[3,31- 33]。事实上,密度-生物量异速指数并不恒定在-3/2或-4/3,而是受到诸多环境因子(如土壤营养水平、冠层光资源)的约束,这一观点己获得越来越多生态学家的认可。很多研究发现,种群个体平均生物量与种群密度的异速指数会随种群的物种基因型而变化,也会随着水分梯度、盐分浓度、海拔以及光照条件等环境变化而有相应的响应[34- 38]。
天然林的稀疏不仅仅是密度层面的变化,在物种的构成上也会出现一定的规律,本研究发现,种群数沿着径级方向也呈现减少,但是常见种在Ⅰ、Ⅱ、Ⅲ级的数量基本没变化,只是偶见种减少迅速。这主要是因为偶见种往往是从群落外大量补充到群落中的(Ⅰ级内非常见种数比常见种多),刚开始幼苗或幼树还是能存活,但随着个体的长大,以及小气候环境与原生地的不同,种内与种间的竞争加剧,必定会淘汰一大部分外来者,即偶见种,少部分会进入后面几个径级的生长与适应,所以在前3个径级的偶见种出现减少,且减少的速度相比后4个径级也明显要快。而常见种就刚好相反,它往往就是本群落中已经占据并适应该群落小气候的物种,所以常见种在Ⅰ、Ⅱ、Ⅲ级的物种数量基本没变化。由于到了Ⅲ级以后,径级变大,个体已经得到壮大,种内与种间的竞争由于发生资源(光照、营养、空间等)竞争,所有种、常见种、偶见种都会继续减少,三者减少速率基本一致。偶见种和常见种的数据的个体变化呈显著正相关,这就意味着常见种树木对非常见种的生长会有一定的促进作用,这与黄世能等对海南岛尖峰岭两类热带山地雨林次生群落在15a演替过程中的林木消长的研究得出的结果不一致[39],可能是本研究采用的“空间代替时间”的方法,未能彻底而全面的反应二者之间的关系,但是常见种相比偶见种适应能力强,生长快,会为偶见种种子的萌发和幼树的生长提供一定有利条件,这也是一种促进作用。Finegan认为,理论上新热带次生林的演替将出现原始林树种或耐阴树种不断侵入的结果[40]。研究发现Ⅰ级以后补充到群落的8个树种绝大多数都是林下耐阴植物,与Finegan结论类似。
4 主要结论
(1)海南文昌铜鼓林国家级自然保护区的天然灌木林不同胸径级植株在群落中呈“倒J型”分布,林分密度是幼树gt;小树gt;成年树。胸径小于4.5cm的幼年个体占有最大百分数(64%),该群落正处于稳定状态。由于个体间为争夺更多的生存空间和资源发生强烈的自疏和它疏作用,所以Ⅰ级到Ⅱ、Ⅲ、Ⅳ、Ⅴ、Ⅵ、Ⅶ级的个体数、密度及种群数均迅速减少,存在明显的稀疏现象。
(2)群落各径级植物地上生物量随着密度增加呈幂函数增加,幂函数方程为:
AGB=6×107N0.4626或 lnAGB=0.4626lnN+17.855
在较小的密度范围内,随林分密度的增加,群落地上总生物量增加较快,但当密度趋于0.6株/m2时,地上生物量变化缓慢,趋于恒定值。
(3)林分密度与群落平均胸径呈负相关关系,密度越大,平均胸径越小。林分密度与平均胸径的幂函数关系式为:N=70.1d-3.5506,R2=0.8808。
(4)选择Yoda提出的幂函数方程对天然灌木林自然稀疏规律进行模拟,林分密度与平均生物量之间具有显著相关性,自疏方程能很好的拟合各数值,具有可靠性。其关系式为:W=2219.1N-0.5374或者为lnW=7.7048-0.5374lnN(R2=0.9292,P=0.0005),稀疏指数α值为0.5374,这与Yoda所提出的3/2指数相差甚远,并不满足-3/2自疏定律。
(5)此次调查的物种有常见种(0.2hm2样地中个体数≥5的种为常见种)41种,非常见种53种。常见种和非常见种沿径级方向均减少,但常见种在Ⅰ、Ⅱ、Ⅲ级内的数量基本没变化,到了Ⅲ级以后,物种数呈直线下降,减少的速度较快;而非常见种在前3个径级内物种减少的非常快,后4个径级也一直在减少,但减少幅度相比在前3个径级内的减少幅度要小。前3个径级所有物种数的减少主要是非常见种的减少,而后4个径级的所有种的减少是二者共同减少的结果。非常见种和常见种呈显著正相关关系(R2=0.6461,P=0.0294)。
[1] Cai X Y, Hong W, Wu C Z, Zhang Y Y, Zhang Y Y, Guo Y, Zhang C Y, Ling C Y, Deng H H, Zhang S G, Qiang Y P, Zhang W J. The self-thinning law ofPinustaiwanensisnatural forest in Wuyishan national nature reserve. Journal of Tropical and Subtropical Botany, 2008, 16(5): 414- 418.
[2] Wang G, Zhang D Y. Theories of Biological Competition. Xian: Shanxi Science and Technology Press, 1996: 28- 371.
[3] Silvertown J W, Doust J L. Introduction to Plant Population Ecology. 3rd ed. New York: Longman Scientific and Technical, 1993: 51- 72.
[4] Begon M, Harper J L, Townsend C R. Ecology: Individuals, Populations and Communities. Oxford: Blackwell Science, 1996: 369- 400.
[5] Li B, Chen J K, Watkinson A R. A literature review on plant competition. Chinese Bulletin of Botany, 1998, 15(4): 18- 29.
[6] Li B, Yang C, Lin P. Ecology. Beijing: Higher Education Press, 2000.
[7] Yoda K, Kira T, Ogawa H, Hozumi K. Self thinning in overcrowded pure stands under cultivated and natural conditions. Journal of Biology of Osaka City University, 1963, 14: 107- 129.
[8] Inoue A, Miyake M, Nishizono T. Allometric model of the Reineke equation for Japanese cypress (Chamaecyparisobtuse) and red pine (Pinusdensiflora) stands. Journal of Forest Research, 2004, 9(4): 319- 324.
[9] Bi H Q, Wan G H, Turvey N D. Estimating the self-thinning boundary line as a density-dependent stochastic biomass frontier. Ecology, 2000, 81(6): 1477- 1483.
[10] Ogawa K. Relationship between mean shoot and root masses and density in an overcrowded population of hinoki (Chamaecyparisobtusa(Sieb. et Zucc.) Endl.) seedlings. Forest Ecology and Management, 2005, 213(1/3): 391- 398.
[11] Ogawa K. Time-trajectory of mean phytomass and density during a course of self-thinning in a sugi (CryptomeriajaponicaD. Don) plantation. Forest Ecology and Management, 2005, 214(1/3): 104- 110.
[12] Pretzsch H. Species-specific allometric scaling under self-thinning: evidence from long- term plots in forest stands. Oecologia, 2006, 146(4): 572-583.
[13] Li T. Response Mechanism of Desert Plant Allometric Exponents and Spatial Patterns to a Precipitation Gradient [D]. Lanzhou: Lanzhou University, 2010.
[14] Gorham E. Shoot height, weight and standing crop in relation to density of monospecific plant stands. Nature, 1979, 279(5709): 148- 150.
[15] White J. Demographic factors in populations of plants // Solbrig O T, ed. Demography and Evolution in Plant Population. Berkeley, California: University of California Press, 1980: 21- 48.
[16] White J. The allometric interpretation of the self-thinning rule. Journal of Theoretical Biology, 1981, 89(3): 475- 500.
[17] Zhou Y B, Yin Y, Yin M F, Zhang F. Self-thinning of natural broadleaved forests in Baishilazi Nature Reserve. Acta Ecologica Sinica, 2011, 31(21): 6469-6480.
[18] Qu Z X, Wang H X, Wu Y S, Jiang H Q, Tang Y G. Plant Ecology. 2nd ed. Beijing: Higher Education Press, 1983: 180- 181.
[19] Chave J, Andalo C, Brown S, Cairns M A, Chambers J Q, Eamus D, Fölster H, Fromard F, Higuchi N, Kira T, Lescure J P, Nelson B W, Ogawa H, Puig H, Riéra B, Yamakura T. Tree allometry and improved estimation of carbon stocks and balance in tropical forests. Ecosystem Ecology, 2005, 145(1): 87- 99.
[20] Milton K, Laca E A, Demment M W. Successional patterns of mortality and growth of large trees in a Panamanian lowland forest. The Journal of Ecology, 1994, 82(1): 79- 88.
[21] Fang J Y, Li Y D, Zhu B, Liu G H, Zhou G Y. Community structures and species richness in the montane rain forest of Jianfengling, Hainan Island, China. Biodiversity Science, 2004, 12(1): 29- 43.
[22] Yang X B, Lin Y, Liang S Q. The forest vegetation of Wuzhishan Mountain in Hainan Island. II. The analysis of forest plant population and forest structure of Wuzhishan Mountain. Natural Science Journal of Hainan University, 1996, 12(4): 311- 323.
[23] Zang R G, Yang Y C, Jiang Y X. Community structure and tree species diversity characteristics in a tropical montane rain forest in Bawangling Natural Reserve, Hainan Island. Acta Phytoecologica Sinica, 2001, 25(3): 270- 275.
[24] Detzin S, Dean C, He F L, Trofymow J A, Wiegand K, Wiegand T. Spatial patterns and competition of tree species in a Douglas-fir chronosequence on Vancouver Island. Ecography, 2006, 29(5): 671- 682.
[25] Comita L S, Hubbell S P. Local neighborhood and species′ shade tolerance influence survival in a diverse seedling bank. Ecology, 2009, 90(2): 328- 334.
[26] Bai J, Tian Y L, H Z R G T, Guo L S, Guo L S. The research on the relationship between the ground biomass, the leaf area index and the stand density inpinustabulaeformisartificial forest. Journal of Arid Land Resources and Environment, 2008, 22(3): 183- 187.
[27] Pan H, Lin D G, Wang G H, Allay Y B, Li B F. Studies on the adaptability ofpinustaedain Fujian-Ⅲ. The density effect laws ofPinustaeda. Acta Agriculture Universitatis Jiangxiensis, 2001, 23(2): 224- 227.
[28] Tong S Z, Sheng W T, Zhang J G. Studies on the density effects of Chinese fir stands. Forest Research, 2002, 15(1): 66- 75.
[29] Duan A G, Zhang J G, Tong S Z, Bao E J, He C Y. Studies on dynamics of diameter structure of Chinese fir plantations and affection of density on it. Forest Research, 2004, 17(2): 178- 184.
[30] Zheng H S, Li M, Wang B G, Feng Y Q. Effect of initial planting spacing on the growth ofBetulaalnoides. Forest Research, 2003, 16(1): 81- 86.
[31] Bi H Q. Stochastic frontier analysis of a classic self- thinning experiment. Austral Ecology, 2004, 29(4): 408- 417.
[32] Fang J Y. Self-thinning rule in plant population. Journal of Ecology and Rural Environment, 1992, (2): 7- 12.
[33] Wang G, Yuan J L. Competitive regulation of plant allometry and a generalized model for the plant self-thinning process. Bulletin of Mathematical Biology, 2004, 66(6): 1875- 1885.
[34] Deng J M, Wang G X, Morris E C, Wei X P, Li D X, Chen B M, Zhao C M, Liu J, Wang Y. Plant mass-density relationship along a moisture gradient in north-west China. Journal of Ecology, 2006, 94(5): 953- 958.
[35] Dai X F, Jia X, Zhang W P, Bai Y, Zhang J, Wang Y, Wang G. Plant height-crown radius and canopy coverage-density relationships determine aboveground biomass-density relationship in stressful environments. Biology Letters, 2009, 5(4): 571- 573.
[36] Chu C J, Maestre F T, Xiao S, Weiner J, Wang Y S, Duan Z H, Wang G. Balance between facilitation and resource competition determines biomass-density relationships in plant populations. Ecology Letters, 2008, 11(11): 1891- 1197.
[37] Chu C J, Weiner J, Maestre F T, Wang Y S, Morris C, Xiao S, Yuan J L, Du G Z, Wang G. Effects of positive interactions, size symmetry of competition and abiotic stress on self-thinning in simulated plant populations. Annals of Botany, 2010, 106(4): 647- 652.
[38] Charru M, Seynave I, Morneau F, Rivoire M, Bontemps J D. Significant differences and curvilinearity in the self-thinning relationships of 11 temperate tree species assessed from forest inventory data. Annals of Forest Science, 2012, 69(2): 195- 205.
[39] Huang S N, Li Y D, Wang B S. Tree population mortality, recruitment and growth during a 15-year period of secondary succession in tropical montane rainforests at Jianfengling, Hainan Island, China. Acta Phytoecologica Sinica, 2000, 24(6): 710- 717.
[40] Finegan B. Pattern and process in neotropical secondary rain forest: the first 100 years of succession. Tends in Ecology and Evolution, 1996, 11(3): 191- 124.
参考文献:
[1] 蔡小英, 洪伟, 吴承祯, 张艳艳, 郭研, 夏鑫, 张彩云, 林存炎, 邓慧华, 张尚炬, 钱永平, 张文娟. 武夷山国家级自然保护区黄山松天然林自然稀疏规律研究. 热带亚热带植物学报, 2008, 16(5): 414- 418.
[2] 王刚, 张大勇. 生物竞争理论. 西安: 陕西科学技术出版社, 1996: 28- 371.
[5] 李博, 陈家宽, Watkinson A R. 植物竞争研究进展. 植物学通报, 1998, 15(4): 18- 29.
[6] 李博, 杨持, 林鹏. 生态学. 北京: 高等教育出版社, 2000.
[13] 李涛. 荒漠植物异速生长指数及其空间格局适应降雨梯度变化的规律与机制研究 [D]. 兰州: 兰州大学, 2010.
[17] 周永斌, 殷有, 殷鸣放, 张飞. 白石砬子国家级自然保护区天然林的自然稀疏. 生态学报, 2011, 31(21): 6469- 6480.
[18] 曲仲湘, 王焕校, 吴玉树, 姜汉桥, 唐廷贵. 植物生态学 (第二版). 北京: 高等教育出版社, 1983: 180- 181.
[21] 方精云, 李意徳, 朱彪, 刘国华, 周光益. 海南岛尖峰岭山地雨林的群落结构、物种多样性以及在世界雨林中的地位. 生物多样性, 2004, 12(1): 29- 43.
[22] 杨小波, 林英, 梁淑群. 海南岛五指山的森林植被Ⅱ. 五指山森林植被的植物种群分析与森林结构分析. 海南大学学报: 自然科学版, 1996, 12(4): 311- 323.
[23] 臧润国, 杨彦承, 蒋有绪. 海南岛霸王岭热带山地雨林群落结构及树种多样性特征的研究. 植物生态学报, 2001, 25(3): 270- 275.
[26] 白静, 田有亮, 韩照日格图, 郭连生. 油松人工林地上生物量、叶面积指数与林分密度关系的研究. 干旱区资源与环境, 2008, 22(3): 183- 187.
[27] 潘辉, 林德根, 王高红, 赖彦斌, 李宝福. 福建省火炬松适应性的研究——Ⅲ密度效应规律. 江西农业大学学报, 2001, 23(2): 224- 227.
[28] 童书振, 盛炜彤, 张建国. 杉木林分密度效应研究. 林业科学研究, 2002, 15(1): 66- 75.
[29] 段爱国, 张建国, 童书振, 保尔江, 何彩云. 杉木人工林林分直径结构动态变化及其密度效应的研究. 林业科学研究, 2004, 17(2): 178- 184.
[30] 郑海水, 黎明, 汪炳根, 冯益谦. 西南桦造林密度与林木生长的关系. 林业科学研究, 2003, 16(1): 81- 86.
[32] 方精云. 植物种群的自然稀疏法则. 农村生态环境, 1992, (2): 7- 12.
ThethinningregularofthetheshrubberyatTonggulingNationalNatureReserveonHainanIsland,China
ZHOU Wei1,2,LONG Cheng1,2,YANG Xiaobo1,2,*,LONG Wenxing1,2,LI Donghai1,2,BU Guangfa1,2
1KeyLaboratoryofProtectionandDevelopmentUtilizationofTropicalCropGermplasmResources,HainanUniversity,MinistryofEducation,Haikou,Hainan570228,China2CollegeofHorticultureandLandscapes,HainanUniversity,Haikou,Hainan570228,China
Woody plants with its DBH≥1.5cm at a 160×160m shrubbery permanent forest plot at Tongguling National Nature Reserve in Wenchang, Hainan, have been investigated., and those plants have been classified into 7 size classes based on DBH to determine the regulation of Thinning. Results showed that: 1) Class Ⅰ (DBHlt;4.5cm) occupied a maximum percentage (64%). The community was “Inverse- J type”, the order of stands density was young trees gt; small treesgt; grow-up trees,the community was in a stable state. It has a typical phenomenon of thinning caused by the self-thinning and alien-thinning which caused by the individuals to compete for more living space and resources. 2) Above ground biomass of each classes increased with the growth of density, in a small density range, above ground biomass increased swiftly with the growth of density, but changed slowly and tended to a constant value when the density tend to be 0.6 plant per square meter, and the power function equation was AGB=6×107N0.4626or lnAGB=0.4626lnN+17.855. 3) It had a negative correlation between the density and the average DBH of each class (larger the density, smaller the average DBH) and its power function equation was:N=70.1d-3.5506,R2=0.8808. 4) We simulated the sel-thinning law put forward by Yoda, and found that there was a significant coherence between stand density and average biomass, the correlation was:W=2219.1N-0.5374or lnW=7.7048-0.5374lnN, thinning indexαvalue was 0.5374, it was largely differ from the 3/2 index put forward by Yoda,did not satisfy the -3/2 law of self-thinning. 5) 41 common species (individuals more than or equals 5 per 0.2hm2) and 51 rare species have been investigated. It has a certain discipline in terms of thinning of species of natural shrubbery.
shrubbery community; thinning; stand density; above ground biomass
国家自然科学基金项目(31060073);中医药行业科研专项海南省代表性区域中药资源保护利用项目(201207002- 03);科技部973计划前期项目(2010CBI34512)
2012- 12- 03;
2013- 04- 18
*通讯作者Corresponding author.E-mail: yangxb62@yahoo.com.cn
10.5846/stxb201212031730
周威,龙成,杨小波,龙文兴,李东海,卜广发.海南铜鼓岭灌木林稀疏规律.生态学报,2013,33(20):6569- 6576.
Zhou W,Long C,Yang X B,Long W X,Li D H,Bu G F.The thinning regular of the the shrubbery at Tongguling National Nature Reserve on Hainan Island,China.Acta Ecologica Sinica,2013,33(20):6569- 6576.
