广西猫儿山不同海拔常绿树种和落叶树种光合速率与氮的关系
2013-12-08白坤栋蒋得斌万贤崇
白坤栋, 蒋得斌, 万贤崇
(1. 广西壮族自治区中国科学院广西植物研究所,桂林 541006;2.中国林业科学研究院林业新技术研究所, 北京 100091;3. 广西猫儿山国家级自然保护区管理局, 桂林 541316)
广西猫儿山不同海拔常绿树种和落叶树种光合速率与氮的关系
白坤栋1,2, 蒋得斌3, 万贤崇2,*
(1. 广西壮族自治区中国科学院广西植物研究所,桂林 541006;2.中国林业科学研究院林业新技术研究所, 北京 100091;3. 广西猫儿山国家级自然保护区管理局, 桂林 541316)
分析广西猫儿山不同海拔常绿和落叶树种的光合作用-氮关系,探讨光合氮利用效率(PNUE)是否受到叶片习性和海拔的影响。落叶树种的PNUE都显著高于常绿树种,这与前者有较低的比叶重(LMA)和较高的单位叶重光合速率(Amass)、氮含量和气孔导度(gs)有密切关系。高海拔树种的PNUE显著低于中低海拔树种的PNUE,这与前者较高的LMA和较低的Amass和gs相关。PNUE和相关的叶片特征的主成分分析表明常绿-落叶树种和低海拔-中海拔-高海拔树种的分布是一个自然过渡的过程。此外,PNUE与土壤碳:氮比没有显著相关性,但与年均温正相关,这表明温度气候是调节PNUE沿海拔变化的主要环境因素。因此,这种叶片习性和温度气候调节的PNUE变化可能是调节猫儿山常绿树种沿海拔形成双峰分布的一种机制。
常绿树种;落叶树种;光合氮利用效率;海拔;双峰分布
植物叶片氮含量(N)是一个影响光合速率(A)的重要因素,这是因为叶片大部分的N分配到了光合机构当中[1]。在同一生境或不同生境中植物的A-N关系都普遍维持正相关关系[2- 3]。A和N的比值(即光合氮利用效率,PNUE)在植物之间可以达到100倍以上的差异[2],因此PNUE常被看作是一个描述植物叶片营养经济、生理和策略的重要的叶片特征[2,4- 6]。过去许多探讨同一生境内植物之间PNUE差异的研究表明,C4植物比C3植物有较高的PNUE,草本植物比木本植物有较高的PNUE,落叶植物比常绿植物有较高的PNUE[5,7- 10]。低PNUE的植物一般比高PNUE的植物有较高的比叶重(LMA)[6,11],这是因为较高的LMA可以通过增加二氧化碳扩散的阻力、减少叶片透射光的比率和增加分配到非光合组分的N比率来降低PNUE[2,6]。另一方面,树种之间PNUE的差异对树种在不同生境间的竞争分布有重要的影响。在全球范围内,进化上低PNUE的树种会在土壤贫瘠的生境或者低温的环境占优势而高PNUE会在干扰程度高或者土壤肥沃的生境中占优势,这种环境造成的PNUE分化是长期自然选择的结果[3,6,12]。
广西猫儿山(25°50′N,101°49′E)位于我国中亚热带,是桂林漓江水源的源头所在地。与东亚的热带地区常绿植物沿海拔的分布特征相似[13],猫儿山常绿植物沿海拔呈现明显的双峰分布[14- 16]。在海拔1300m 以下,常绿植物占优势。在海拔1300—1800m的中山地带分布着水青冈占优势的落叶林带。在海拔1800m 以上,常绿植物重新占据优势。根据猫儿山综合科学考察报告,气候类型随着海拔的增加依次为山地亚热带气候、山地暖温带气候和山地中温带气候,土壤表土有机质和氮含量随着海拔的增加而增多,这意味着常绿植物沿着海拔形成双峰分布可能与气候和土壤基质的变化有密切的关系[14,16]。尽管这些树种分布和环境之间有密切的关系,但至今尚未见是否树种间PNUE差异参与调节树种海拔分布的研究报道。本研究分析猫儿山不同海拔共生的常绿和落叶树种的A-N关系,本研究拟解决的关键科学问题是:(1)PNUE是否受到叶片习性和海拔的影响?(2)PNUE与其它叶片特征有何关联?这些问题的探讨可以揭示A-N关系在猫儿山常绿植物沿海拔形成双峰分布中的作用,为我国亚热带山地森林生物多样性的形成以及预测这类森林叶片营养经济、生理和生活史策略提供科学依据。
1 材料和方法
1.1 样地描述和树种选择
研究样地在广西猫儿山3种典型的森林中,即低海拔的山地亚热带气候下的常绿阔叶林、中海拔的山地暖温带气候下的水青冈混交林和高海拔的山地中温带气候下铁杉混交林。根据猫儿山自然保护区九牛塘气象观测站(1200m)的观测记录,区域年均温12.8℃,月均最高最低温差为19.8℃,年降水量2510mm[16]。低海拔常绿阔叶林样地(海拔约为900m)的土壤基质为山地红壤,表层土(0—20cm)有机碳和全氮分别为4.33%和0.34%,年均温为14.1℃,选择的常绿树种是罗浮栲、细枝栲、木荷和华东润楠,选择的落叶树种是枫香、拟赤杨、君迁子和光皮桦。中海拔水青冈混交林样地(海拔约为1500m)的土壤基质为山地黄棕壤,表层土有机碳和全氮分别为9.74%和0.52%,年均温为11.5℃,选择的落叶树种是亮叶水青冈、青榨槭、缺萼枫香和钟萼木,选择的常绿树种是铁锥栲、曼青冈、桂南木莲和银木荷。高海拔铁杉混交林样地(海拔约为1900m)的土壤基质是泥炭土,表层土有机碳和全氮分别为16.23%和0.93%,年均温为8.1℃,选择的常绿树种是南方铁杉、褐叶青冈、厚叶杜鹃和包石栎,选择的落叶树种是白蜡树、中华槭和红叶木姜子。三类森林所选代表树种总共23种,来自于13个科,其中壳斗科的7个树种是这些森林的主要优势种(表1)。
1.2 实验方法
在2010年和2011年夏季(7—8月)用LI- 6400气体交换测定仪测定叶片光合特征。由于森林类型和物候是冠层树种决定的,选择冠层的样树进行采样,每个树种选择3—5株。所选样树平均株高超过12m,野外条件难以直接测定这些样树枝条叶片的光合作用,故采取离体测定[17- 18],利用高枝剪剪下枝条并迅速插入水中减少水分蒸腾的影响。单位叶面积的光合速率(Aarea)和气孔导度(gs)测定的时间是晴天9:30—11:30之间进行,测定过程中保持叶片的环境因子适宜且相对稳定。测定时叶室温度为20—25℃,二氧化碳浓度为380μmol m-2s-1,光强为1500μmol m-2s-1,预备实验表明此时光强对所有测定的植物而言都达到饱和光之上,且并未对植物的光合作用产生抑制作用。气体交换测定完毕后,叶片就收集下来测定LMA(叶片干重/叶片面积)和N含量[19]。随后,单位叶重的光合速率(Amass)由Aarea/LMA计算,而PNUE则由Amass/N计算。

表1 猫儿山不同海拔研究树种的生境特征、拉丁名、科名和叶片习性
1.3 数据处理
为了满足正态分布和方差齐性,所有变量都转化成对数形式(log10)。采用双因素方差分析(two-way ANOVA)叶片习性和海拔对叶片特征的影响。用主成分分析(PCA)叶片特征的多元关联,并用多元方差分析(MANOVA)检验最初两个主成分树种分数是否受到叶片习性或者海拔的影响。利用标准主轴(SMA)回归分析叶片特征之间的关系以及不同叶片习性和不同海拔之间回归关系斜率和截距的差异。此外,PNUE与年均温和土壤碳:氮比的关系也用SMA回归分析。所有数据的统计、分析和作图采用SPSS12.0、SMART 2.0和SigmaPlot10.0等软件完成。
2 结果和分析
6个叶片特征在所有树种之间的变化幅度为2—8倍,而且这些特征都不同程度受到叶片习性和海拔的影响(表2和表3)。LMA平均值113 g/m2,范围是52—227g/m2;常绿树种和高海拔树种有显著较高的LMA。N的平均值2.43%,范围是1.70%—3.48%;落叶树种有显著较高的N,但是不同海拔之间的N没有显著差异。Amass的平均值123 nmol g-1s-1,范围是29—234 nmol g-1s-1;落叶树种和中海拔树种有显著较高的Amass。Aarea的平均值11.8μmol m-2s-1, 范围是6.6—16.9μmol m-2s-1;落叶树种有显著较高的Aarea,但是不同海拔之间的Aarea没有显著差异。gs的平均值184mmol m-2s-1, 范围是66—292mmol m-2s-1;常绿树种和高海拔树种有显著较低的gs。PNUE平均值69μmol mol-1s-1, 范围是66—292μmol mol-1s-1;常绿树种和高海拔树种有显著较低的PNUE。
表2猫儿山不同海拔常绿和落叶树种叶片特征的平均值±标准差(n=3—5)和多元方差分析结果
Table2Mean±SD(n=3—5)ofleaftraitsandtheresultsofthemultivariateanalysisofvariance(MANOVA)forevergreenanddeciduoustreespeciesatdifferentaltitudesonMao′erMountain
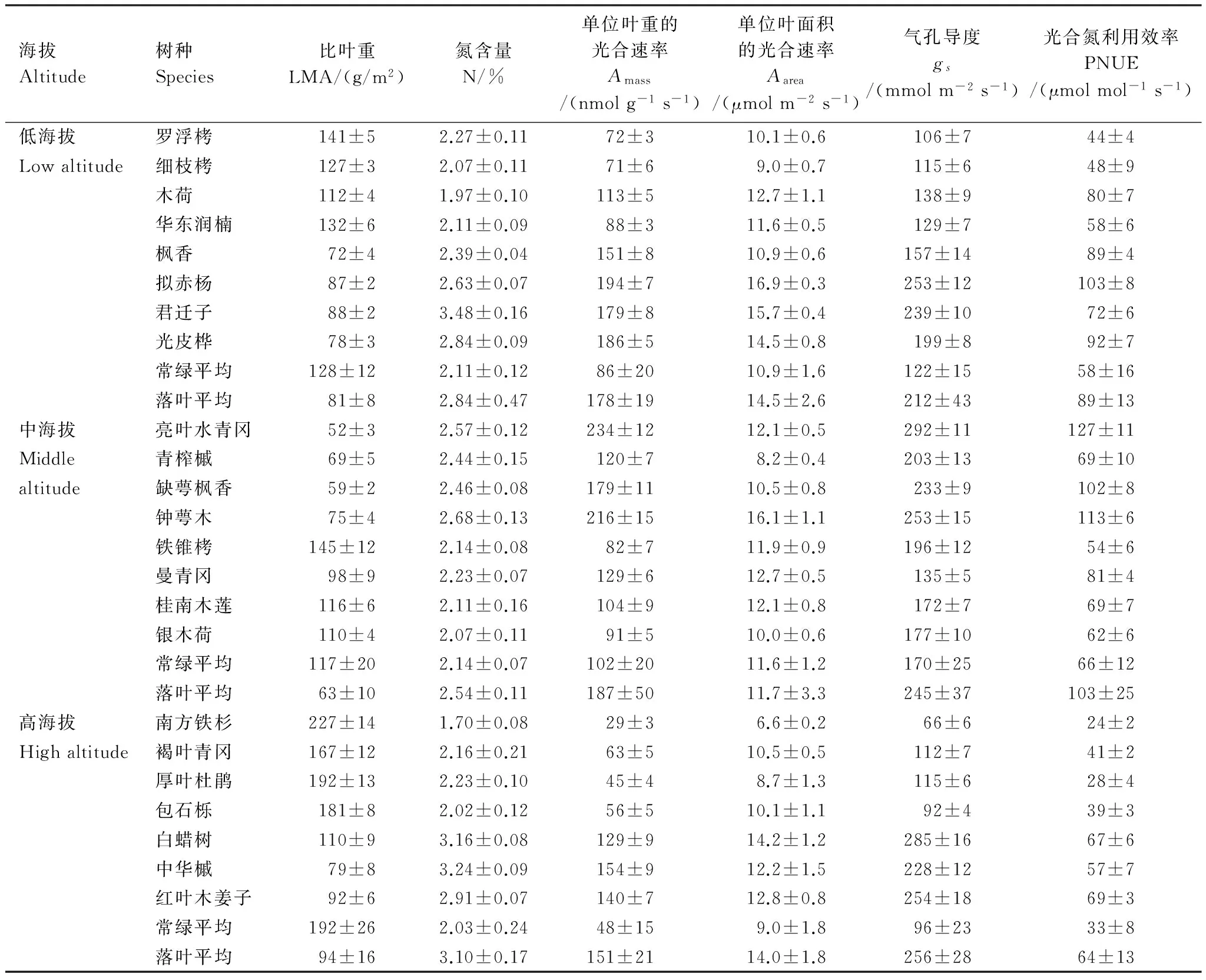
海拔Altitude树种Species比叶重LMA/(g/m2)氮含量N/%单位叶重的光合速率Amass/(nmolg-1s-1)单位叶面积的光合速率Aarea/(μmolm-2s-1)气孔导度gs/(mmolm-2s-1)光合氮利用效率PNUE/(μmolmol-1s-1)低海拔罗浮栲141±52.27±0.1172±310.1±0.6106±744±4Lowaltitude细枝栲127±32.07±0.1171±69.0±0.7115±648±9木荷112±41.97±0.10113±512.7±1.1138±980±7华东润楠132±62.11±0.0988±311.6±0.5129±758±6枫香72±42.39±0.04151±810.9±0.6157±1489±4拟赤杨87±22.63±0.07194±716.9±0.3253±12103±8君迁子88±23.48±0.16179±815.7±0.4239±1072±6光皮桦78±32.84±0.09186±514.5±0.8199±892±7常绿平均128±122.11±0.1286±2010.9±1.6122±1558±16落叶平均81±82.84±0.47178±1914.5±2.6212±4389±13中海拔亮叶水青冈52±32.57±0.12234±1212.1±0.5292±11127±11Middle青榨槭69±52.44±0.15120±78.2±0.4203±1369±10altitude缺萼枫香59±22.46±0.08179±1110.5±0.8233±9102±8钟萼木75±42.68±0.13216±1516.1±1.1253±15113±6铁锥栲145±122.14±0.0882±711.9±0.9196±1254±6曼青冈98±92.23±0.07129±612.7±0.5135±581±4桂南木莲116±62.11±0.16104±912.1±0.8172±769±7银木荷110±42.07±0.1191±510.0±0.6177±1062±6常绿平均117±202.14±0.07102±2011.6±1.2170±2566±12落叶平均63±102.54±0.11187±5011.7±3.3245±37103±25高海拔南方铁杉227±141.70±0.0829±36.6±0.266±624±2Highaltitude褐叶青冈167±122.16±0.2163±510.5±0.5112±741±2厚叶杜鹃192±132.23±0.1045±48.7±1.3115±628±4包石栎181±82.02±0.1256±510.1±1.192±439±3白蜡树110±93.16±0.08129±914.2±1.2285±1667±6中华槭79±83.24±0.09154±912.2±1.5228±1257±7红叶木姜子92±62.91±0.07140±712.8±0.8254±1869±3常绿平均192±262.03±0.2448±159.0±1.896±2333±8落叶平均94±163.10±0.17151±2114.0±1.8256±2864±13

表3 猫儿山不同海拔常绿和落叶树种叶片特征的双因素方差分析结果
PCA表明最初的两个主成分解释了这6个叶片特征90.1%的总变异,其中主成分1(PC1)解释了79.3%而主成分2(PC2)解释了10.8%(表4)。MANOVA表明叶片习性和海拔对PC1和PC2有显著影响(表4),这意味着常绿和落叶树种之间或者不同海拔树种之间的差别可以通过树种分数在PC1和PC2的二维图上区分出来。这种二维图表明常绿-落叶树种或者不同海拔树种在两个主成分上有一定的重叠,这反映了常绿-落叶树种或者低海拔-中海拔-高海拔树种的分布是一种自然的过渡(图1)。
表4因子负荷、特征值、解释的方差百分比和最初两个主成分的多元方差分析(MANOVA)检验
Table4Factorloading,eigenvalues,andthepercentageofvarianceexplained,andthemultivariateanalysisofvariance(MANOVA)testsofthefirsttwoprincipalcomponents
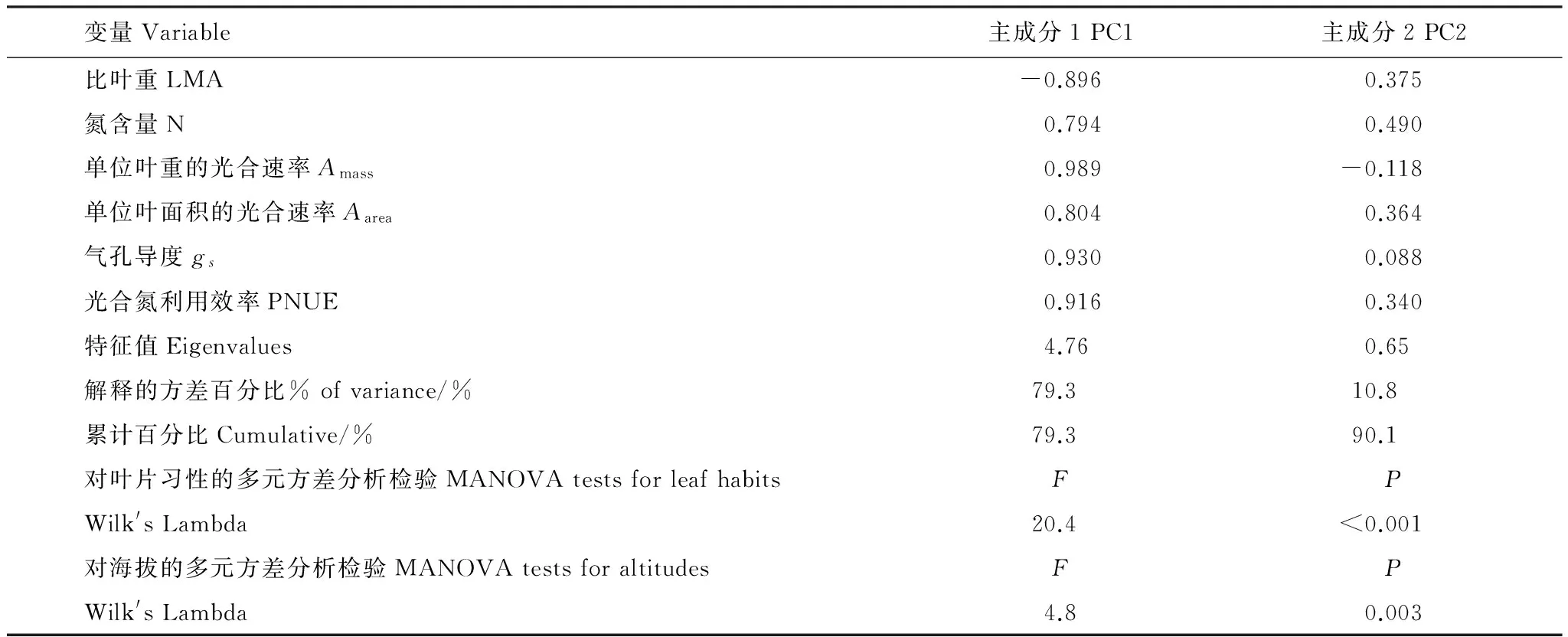
变量Variable主成分1PC1主成分2PC2比叶重LMA-0.8960.375氮含量N0.7940.490单位叶重的光合速率Amass0.989-0.118单位叶面积的光合速率Aarea0.8040.364气孔导度gs0.9300.088光合氮利用效率PNUE0.9160.340特征值Eigenvalues4.760.65解释的方差百分比%ofvariance/%79.310.8累计百分比Cumulative/%79.390.1对叶片习性的多元方差分析检验MANOVAtestsforleafhabitsFPWilk'sLambda20.4lt;0.001对海拔的多元方差分析检验MANOVAtestsforaltitudesFPWilk'sLambda4.80.003

图1 对常绿和落叶树种和3个海拔的最初两个主成分的主成分分析Fig.1 Principal component analysis for evergreen species (closed circles) and deciduous species (open circles) and the three altitudes for the first two components
SMA回归分析表明,6个叶片特征之间是高度相关联的(表5)。所有树种的Amass与LMA负相关而与N、Aarea、gs和PNUE正相关。由于Amass=Aarea/LMA,但是Aarea不受海拔的影响而且受到叶片习性的影响弱于Amass,因此Amass同时受到叶片习性和海拔的显著影响主要是受到LMA的调节,即常绿树种和高海拔树种较低的Amass是与其较高的LMA有密切关系(表3和表5)。Amass-N关系斜率和截距分别是3.01和0.91。常绿树种比落叶树种有较高的Amass-N关系斜率(P=0.012),这意味着相同Amass的情况下常绿树种需要投入更多的N,从而导致常绿树种有较低的PNUE(图2)。中海拔树种有显著较大的Amass-N关系截距(Plt;0.001),这意味着在相同N的情况下中海拔树种有较高的Amass,从而导致中海拔树种有较高的PNUE(图2)。PNUE-LMA的负相关和PNUE-gs的正相关关系进一步说明PNUE受到了叶片结构和气孔的调节(表5和图3)。此外,PNUE与环境因子有密切的关系。PNUE与年均温显著正相关,但与土壤碳:氮比无显著相关性(图4)。
表5猫儿山不同海拔所有树种叶片比叶重(LMA)、单位叶重的光合速率(Amass)、氮含量(N)、单位叶面积的光合速率(Aarea)、气孔导度(gs)和光合氮利用效率(PNUE)之间的标准主轴回归分析
Table5Standardizedmajoraxisregressionanalysisamongleafmassperarea(LMA),nitrogencontent(N),photosyntheticratepermass(Amass),photosyntheticrateperarea(Aarea),stomatalconductance(gs)andphotosyntheticnitrogenuseefficiency(PNUE)acrosstreespeciesatdifferentaltitudesonMao′erMountain

log(LMA)log(Amass)log(N)log(Aarea)log(gs)log(PNUE)log(LMA)-0.72;3.49-2.17;2.85-1.70;3.83-0.96;4.16-0.91;3.66log(Amass)0.86***3.01;0.912.36;3.141.33;-0.921.26;-0.24log(N)0.38***0.52***0.79;-0.450.44;-0.610.42;-0.38log(Aarea)0.24*0.60***0.44**0.56;-0.190.53;0.10log(gs)0.65***0.77***0.62***0.49***0.95;0.52log(PNUE)0.81***0.91***0.24*0.50***0.62***
标准主轴回归斜率和截距(斜率;截距)列在矩阵的右边部分(应变量是列1,自变量是行1),决定系数(r2)和显著性水平(***Plt;0.001, **Plt;0.01,*Plt;0.05)列在矩阵的左边部分Standardized major axis regression slopes and intercepts (slope; intercept) are given in the upper right section of the matrix (y variable is column 1, x variable in row 1). Coefficients of determination (r2) and significant level (***Plt;0.001, **Plt;0.01,*Plt;0.05) are given in the lower leaf section of the matrix

图2 猫儿山不同海拔所有树种光合氮利用效率(PNUE)与单位叶重的光合速率(Amass)和氮含量(N)的三维关系图Fig.2 Three-dimensional relationships between photosynthetic nitrogen use efficiency (PNUE) and photosynthetic rate per mass (Amass) and nitrogen content (N) across all species at different altitudes on Mao′er Mountain
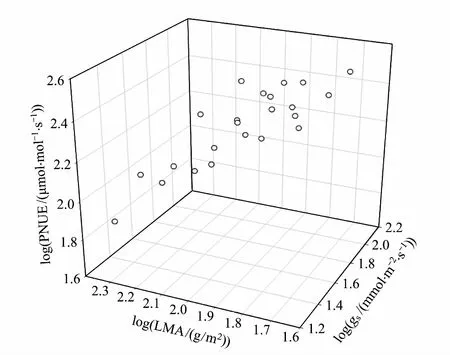
图3 猫儿山不同海拔所有树种光合氮利用效率(PNUE)与比叶重(LMA)和气孔导度(gs)的三维关系图Fig.3 Three-dimensional relationships between photosynthetic nitrogen use efficiency (PNUE) and leaf mass per area (LMA) and stomatal conductance (gs) across all species at different altitudes on Mao′er Mountain
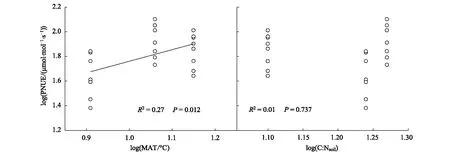
图4 猫儿山不同海拔所有树种光合氮利用效率(PNUE)与年均温(MAT)和土壤碳∶氮比(C∶Nsoil)的标准主轴回归关系Fig.4 Standardized major axis regression relationship between photosynthetic nitrogen use efficiency (PNUE) and mean annual temperature (MAT) and soil carbon∶nitrogen ratio (C∶Nsoil) across all species at different altitudes on Mao′er Mountain
3 讨论
PNUE是一个与植物叶片经济、生理和策略密切相关的叶片特征[6]。本研究表明, PNUE受到叶片习性的显著影响,落叶树种比常绿树种有较高的PNUE,这与过去的研究一致[20- 21]。Amass-N的关系表明,落叶树种较高的PNUE是因其在相同N增量的情况下有较高的Amass增量。落叶树种较高的PNUE与其较低的LMA也有密切的关系,因为较低的LMA的树种可以分配更多的氮比率到光合组分并最终提高PNUE[2,20]。落叶树种较高的PNUE还与其较高的gs密切相关,因为较高的gs可以提高胞间二氧化碳浓度从而促进PNUE[6]。在进化上,叶片寿命短的树种有较低的LMA、较高的N、较高的Amass和PNUE,这些特征代表树种可以快速生长,有利于其在土壤肥沃的环境获得竞争优势[2,21- 22]。与落叶树种相反,常绿树种有较高的LMA、较低的N、较长的叶片寿命和较低的PNUE,这些特征可以使常绿树种有一个更长的叶片氮平均滞留时间和光合季节、以较低的叶片营养成本替换衰老的叶片、有更多能量和物质投入在结构性物质上(如纤维素等)从而降低被虫食可能性和风吹冰害等机械性损伤以及降低木质部空穴化的可能性促进水分传输的安全性等得到提升,因此这些特征有利于常绿植物在土壤贫瘠或者低温的环境占据优势[12,23]。
Amass-N的关系分析表明,PNUE也受到海拔的显著调节。高海拔树种比中低海拔树种有显著较低的PNUE,这与前者LMA较高有关系,也与Amass和gs较低是一致的。这种高海拔PNUE受到抑制的现象与过去的研究相似[24]。既然PNUE都受到叶片习性和海拔的调节,那么这对常绿树种沿海拔形成双峰分布有何重要的意义呢?常绿和落叶树种的竞争分布是对土壤资源和气候的长期适应而形成的[12]。在本研究的森林中,土壤水资源并不是一个限制常绿和落叶树种生长的因子,因为猫儿山年均降水量超过2500mm[16]。猫儿山低海拔常绿阔叶林、中海拔水青冈混交林和高海拔铁杉混交林的土壤碳:氮比分别为12.7、18.7和17.5,这些比值都低于全球热带亚热带山地(15.4)、暖温带山地(20.6)和冷温带山地(20.2)的平均值[25]。低海拔常绿阔叶林的碳:氮比低于全球土壤碳:氮比的临界点14.3[26],可以认为低海拔的土壤受到氮限制。猫儿山低海拔树种叶片N平均值(2.47%)比全球的平均值(2.01%)[27]高出23%,这表明叶片水平的氮含量可能不受限制,但是低海拔树种的PNUE平均值74μmol mol-1s-1明显低于全球树种的PNUE平均值130μmol mol-1s-1[5],这种反差可能是猫儿山低海拔树种叶片的氮分配到光合机构的比例相对较低从而造成PNUE大幅度低于全球平均水平[6,10]。由于土壤碳:氮比是衡量土壤有机质降解程度和质量的指标,低的土壤碳:氮比意味着土壤相对较贫瘠[28]。PNUE与土壤碳:氮比没有显著的相关性(图4),这意味着土壤贫瘠程度可能不是调节猫儿山PNUE沿海拔变化的主要环境因素。另一方面,随着海拔的升高,年均温也随之降低,PNUE与年均温正相关(图2),这表明温度气候是调节猫儿山PNUE沿海拔变化的主要环境因素。在低海拔山地亚热带气候下,一年中不利于植物生长的季节很短,有利于常绿树种在一年中积累更多的碳收益,促进常绿树种占据优势。在中海拔山地暖温带气候下,一年中不适宜的生长季节变长,冬季的冰冻灾害容易造成阔叶树种枝条导管栓塞和断裂,极易造成树种枝条掉落甚至整株死亡,这很容易形成林窗的干扰[14]。在干扰容易发生的生境,高PNUE的落叶树种就容易形成优势[6],猫儿山中海拔的落叶树种就有最高的PNUE。在高海拔的山地中温带气候下,一年中不适宜的生长季节进一步延长,树种叶片形成高水平的LMA来抵御高海拔带来的低温和强风影响, 这与过去的研究一致[24,29]。由于LMA和PNUE的负相关关系(图3),常绿树种低水平的PNUE就更能够适应高海拔的环境,这与过去认为低水平的PNUE是对低温环境的一种适应的研究结论是一致的[6]。值得注意的是,PNUE和相关叶片特征的主成分分析表明常绿-落叶树种或者不同海拔树种在两个主成分上有一定的重叠,这反映了常绿-落叶树种或者低海拔-中海拔-高海拔树种的分布是一种自然的过渡,这与我国热带喀斯特森林常绿和落叶树种的研究结果类似[30]。
4 结论
PNUE受到叶片习性和海拔的影响。常绿树种和高海拔树种有显著较低的PNUE,这与前者有较高的LMA和较低的Amass和gs有密切关系。PNUE和相关的叶片特征的PCA表明常绿-落叶树种和低海拔-中海拔-高海拔树种的分布是一个自然过渡的过程。PNUE与不同海拔的土壤碳:氮比没有显著相关性而与年均温正相关,这意味着温度气候是调节猫儿山PNUE沿海拔变化的主要环境因素。因此,这种叶片习性和温度气候调节的PNUE变化可能是调节猫儿山常绿树种沿海拔形成双峰分布的一种机制。
[1] Evans J R. Photosynthesis and nitrogen relationships in leaves of C3plants. Oecologia, 1989, 78: 9- 19.
[2] Wright I J, Reich P B, Cornelissen J H C, Falster D S, Garnier E, Hikosaka K, Lamont B B, Lee W, Oleksyn J, Osada N, Poorter H, Villar R, Warton D I, Westoby M. Assessing the generality of global leaf trait relationships. New Phytologist, 2005, 166: 485- 496.
[3] Reich P B, Oleksyn J, Wright I J. Leaf phosphorus influences the photosynthesis-nitrogen relation: a cross-biome analysis of 314 species. Oecologia, 2009,160: 207- 212.
[4] Field C B, Mooney H A. The photosynthesis-nitrogen relationship in wild plants//Givnish T J, ed. On the economy of plant form and function. Cambridge University Press, Cambridge, 1986: 22- 25.
[5] Hikosaka K, Hanba Y T, Hirose T, Terashima I. Photosynthetic nitrogen-use efficiency in leaves of woody and herbaceous species. Functional Ecology, 1998, 12: 896- 905.
[6] Hikosaka K. Interspecific difference in photosynthesis-nitrogen relationship: patterns, physiological causes, and ecological importance. Journal of Plant Research, 2004, 117: 481- 494.
[7] Anten N P R, Schieving F, Werger M J A. Patterns of light and nitrogen distribution in relation to whole canopy gain in C3and C4mono- and dicotyledonous species. Oecologia, 1995, 101: 504- 513.
[8] Hikosaka K, Hirose T. Photosynthetic nitrogen-use efficiency in evergreen broad-leaved woody species coexisting in a warm-temperate forest. Tree Physiology, 2000, 20: 1249- 1254.
[10] Hikosaka K, Shigeno A. The role of Rubisco and cell walls in the interspecific variation in photosynthetic capacity. Oecologia, 2009, 160: 443- 451.
[11] Poorter H, Evans J R. Photosynthetic nitrogen use efficiency of species that differed inherently in specific leaf area. Oecologia, 1998, 116: 26- 37.
[12] Givnish T J. Adaptive significance of evergreen vs. deciduous leaves: solving the triple paradox. Silver Fennica, 2002, 36: 703- 743.
[13] Kikuzawa K. Geographical distribution of leaf life span and species diversity of trees simulated by a leaf-longevity model. Vegetatio, 1996, 122: 61- 67.
[14] Cao K F.Fagusdominance in Chinese montane forests: natural regeneration ofFaguslucidaandFagushayataevar.pashannca. Ph.D. Thesis, Wageningen, 1995.
[17] Koch G W, Sillett S C, Jennings G M, Davis S D. The limits to tree height. Nature, 2004, 428: 851- 854.
[19] Bai K D, Jiang D B, Cao K F, Liao D B, Wan X C. The physiological advantage of an ecological filter species,Indocalamuslongiauritus, over co-occurringFaguslucidaandCastanopsislamontiiseedlings. Ecological Research, 2011, 26: 15- 25.
[20] Reich P B, Kloeppel B D, Ellsworth D S, Walters M B. Different photosynthesis-nitrogen relations in deciduous hardwood and evergreen coniferous tree species. Oecologia, 1995, 104: 24- 30.
[21] Takashima T, Hikosaka K, Hirose T. Photosynthesis or persistence: nitrogen allocation in leaves of evergreen and deciduousQuercusspecies. Plant, Cell and Environment, 2004, 27: 1047- 1054.
[22] Wright I J, Reich P B, Westoby M, Ackerly D D, Baruch Z, Bongers F, Cavender-Bares J, Chapin F S, Cornelissen J H C, Diemer M, Flexas J, Garnier E, Groom P K, Gulias J, Hikosaka K, Lamont B B, Lee T, Lee W, Lusk C, Midgley J J, Navas M L, Niinemets Ü, Oleksyn J, Osada N, Poorter H, Poot P, Prior L, Pyankov V I, Roumet C, Thomas S C, Tjoelker M G, Veneklaas E J, Villar R. The world-wide leaf economics spectrum. Nature, 2004, 428: 821- 827.
[23] Kikuzawa K. A cost-benefit analysis of leaf habit and leaf longevity of trees and their geographical pattern. American Naturalist, 1991, 138:1250- 1263.
[24] Hikosaka K, Nagamatsu D, Ishii H S, Hirose T. Photosynthesis-nitrogen relationships in species at different altitudes on Mount Kinabalu, Malaysis. Ecological Research, 2002, 17: 305- 313.
[25] Post W M, Pastor J, Zinke P J, Stangenberger G. 1985. Global patterns of soil nitrogen storage. Nature, 1985, 317: 613- 616.
[26] Cleveland C C, Liptzin D. C: N:P stoichiomentry in soil: is there a “Redfield ratio” for the microbial biomass?. Biogeochemistry, 2007, 85: 235- 252.
[27] Reich P B, Oleksyn J. Global patterns of plant leaf N and P in relation to temperature and latitude. Proceedings of the National Academy of Sciences of the United States of America, 2004, 101: 11001- 11006.
[28] Batjes N H. Total carbon and nitrogen in soils of the world. European Journal of Soil Science, 47: 151- 163.
[29] Tang C Q, Oshawa M. Altitudinal distribution of evergreen broad-leaved trees and their leaf-size pattern on a humid subtropical mountain, Mt. Emei, Sichuan, China. Plant Ecology, 1999, 145: 221- 233.
[30] Fu P L, Jiang Y J, Wang A Y, Brodribb TJ, Zhang J L, Zhu S D, Cao K F. Stem hydraulic traits and leaf-water stress tolerance are co-ordinated with the leaf phenology of angiosperm trees in an Asian tropical dry karst forest. Annals of Botany, 2012,doi: 10.1093/aob/mcs092
参考文献:
[9] 郑淑霞,上官周平. 不同功能型植物光合特性及其与叶氮、比叶重的关系. 生态学报,2007, 27(1):171- 181.
[15] 朱彪,陈安平,刘增力,李光照,方精云. 广西猫儿山植物群落物种组成、群落结构及树种多样性的垂直分布格局. 生物多样性,2004,12(1):44- 52.
[16] 黄金玲,蒋得斌. 广西猫儿山自然保护区综合科学考察. 湖南:湖南科学技术出版社,2002:165- 181.
[18] 屈小云,于贵瑞,盛文萍,方华军.中国东部亚热带样带森林优势植物叶片的水分利用效率和氮素利用效率.应用生态学报,2012,23(3):587- 594.
Photosynthesis-nitrogenrelationshipinevergreenanddeciduoustreespeciesatdifferentaltitudesonMao′erMountain,Guangxi
BAI Kundong1,2, JIANG Debing3, WAN Xianchong2,*
1GuangxiInstituteofBotany,GuangxiZhuangAutonomousRegionandChineseAcedemyofSciences,Guilin541006,China2InstituteofNewForestTechnology,ChineseAcademyofForestry,Beijing100091,China3BureauofGuangxiMao′erMountainNatureReserve,Guilin541316,China
The present study analyzed the photosynthesis-nitrogen relationship in evergreen and deciduous tree species at different altitudes on Mao′er Mountain, Guangxi, and explored whether photosynthetic nitrogen use efficiency (PNUE) was affected by leaf habit and altitude. Deciduous tree species had significanlty higher PNUE than evegreen tree species because of lower leaf mass per area (LMA) and higher photosynthetic rate per mass (Amass), nitrogen content and stomatal conductance (gs) in the former. Tree species at high altitude had significantly lower PNUE than those at middle and low altitudes due to higher LMA and lowerAmassandgsin the former. Principal component analysis for PNUE and its related leaf traits suggested that the tree species distribution of evergreen-deciduous and low-middle-high altitude were in the process of natural transition. In addition, PNUE was not related to soil carbon: nitrogen ratio, but significantly positive related to mean annual temperature, suggesting that temperature climate was the main environmental factor that regulated the altitudianl changes in PNUE. Therefore, this differential PNUE that was affected by leaf habit and temperature climate was a mechanism for the bimodal distribution of evergreen tree species along the altitudes on Mao′er Montain.
evergreen tree species; deciduous tree species; photosynthetic nitrogen use efficiency; altitude; bimodal distribution
国家自然科学基金资助项目(31100285); 中央级公益性科研院所基本科研业务费专项资金(CAFINT2009C08); 国家科技支撑计划课题(2012BAC16B01)
2012- 05- 20;
2012- 10- 23
*通讯作者Corresponding author.E-mail: wxc@caf.ac.cn
10.5846/stxb201205200750
白坤栋, 蒋得斌,万贤崇.广西猫儿山不同海拔常绿树种和落叶树种光合速率与氮的关系.生态学报,2013,33(16):4930- 4938.
Bai K D, Jiang D B, Wan X C.Photosynthesis-nitrogen relationship in evergreen and deciduous tree species at different altitudes on Mao′er Mountain, Guangxi.Acta Ecologica Sinica,2013,33(16):4930- 4938.
