大花萱草‘金娃娃’的花器结构和繁育系统观察
2013-12-06赵天荣蔡建岗施永泰倪建刚
赵天荣,蔡建岗,施永泰,倪建刚,沈 岚
(宁波市农业科学研究院,浙江 宁波 315040)
大花萱草‘金娃娃’的花器结构和繁育系统观察
赵天荣,蔡建岗,施永泰,倪建刚,沈 岚
(宁波市农业科学研究院,浙江 宁波 315040)
通过观察,运用杂交指数、花粉/胚珠、花粉及柱头活力、坐果率等指标对大花萱草‘金娃娃’(Hemerocallishybriduscv.‘Stella de oro’)的花部特征、开花动态及授粉特性等进行研究。在人工栽培条件下,金娃娃萱草5-9月开花,花苔分3~4次抽生,首次开花群体花期32 d,单苔花期9~16 d,单花花期1 d。雄蕊短于花柱,花药与柱头相差2.71 cm,在花朵开放的整个过程中雌、雄蕊的相对位置始终不变。开花前1 d的花粉活力最强,开花前1 d和开花当天的柱头具有较强的可授性。杂交指数为4,花粉/胚珠值为1 164,结合坐果率,可以判断其繁育系统属于兼性异交,部分自交亲和,需要传粉者。特殊的花部结构、缺少传粉者和花期短可能是金娃娃萱草结实率较低的原因。
大花萱草‘金娃娃’;花部特征;繁育系统;杂交指数;花粉/胚珠
萱草属(Hemerocallis)是百合科(Liliaceae)多年生草本,全属植物约14种,主要分布于亚洲温带至亚热带地区,我国有11种原产种及部分自然杂交变种;少数分布在日本、朝鲜和前苏联[1-2]。19世纪末,欧美兴起了群众性培育萱草新品种的活动。1893年英国人Georgeyeld 登记注册了第1个萱草栽培品种Aprioot[3]。20世纪以来,培育出了大量优质杂交萱草。目前美国萱草协会每年都有近万个萱草新品种登记注册,萱草已经成为品种最丰富的宿根花卉之一[4]。
系统地了解植物花的结构特征和繁育特性,是认识植物生活史的前提,也是开展相关研究必备的知识背景[5]。但是迄今为止,国内对萱草的研究主要集中在细胞学、孢粉学、植物分类和倍性育种等方面[6-12]。国外对萱草的研究主要集中在衰老基因、抗病基因、倍性育种和遗传特性等方面[13-16]。虽然萱草育种历史悠久,在美国萱草协会注册的园艺品种已经有6万多个,但较系统地对萱草花器结构、花期物候和开花授粉等繁育特性方面的研究少见报道。然而花粉有无活力或活力的高低对雌蕊授粉成功率有直接的影响[17]。生产实践中,为了解决亲本花期不一致或远距离杂交的困难,需要保存花粉的活力,因此花粉生活力测定是进行人工辅助授粉或杂交授粉的基础[18]。
由于目前国内引进的萱草品种较多,而且这些品种包括了二倍体、四倍体、六倍体等材料,萱草不同倍性的品种间,甚至相同倍性的品种间,开花授粉特性也各有差异,因此本研究选用目前国内栽培最为广泛的大花萱草新品种‘金娃娃’(Hemerocallishybridus‘Stella de Oro’)为试验材料,采用连续实地观察、显微观察、套袋授粉等方法,揭示该物种花器结构、花期物候、开花动态和开花授粉等特性,并对萱草的繁育系统类型进行了判断,另外就其结实率较低的原因进行了探讨,为萱草的种质资源创新和新品种选育提供理论依据。
1 材料与方法
1.1材料 试验地设于宁波市高新农业实验园区萱草种质资源圃内,海拔4.0~5.8 m,年最低气温-8.5 ℃,年最高气温39.5 ℃;年平均降水量1 440.37 mm。宁波地区春、秋和冬季雨水充足,气候湿润,夏季温度较高,光照较强且干旱少雨。金娃娃萱草试验小区采取作畦式栽培,畦长20 m,畦宽1.3 m,种植行距25 cm,株距25 cm,采取节约型栽培,全年不浇水,抽苔前和花后各施一次复合肥。以通过分株方式繁殖栽培2年的金娃娃萱草为试验材料,种质资源圃内种有丰富的萱草种植资源。
1.2花部特征、花期物候和开花动态观察 按照Dafni[19]的标准于2010年4―10月记录金娃娃萱草的群体花期和开花进程。在蕾期选择生长健壮的10株萱草,每株标记5朵花,记录各植株开花状况。
参照关文灵等[20]的方法观察记载花朵绽放进程、散粉时间、开放顺序和花期,以及花朵的组成颜色、形状、花冠直径。
1.3花粉活力和柱头可授性的测定 在盛花期10:00,选择开花前4、3、2、1 d的蕾和开放当天的花朵分别采集花药,将花粉撒在载玻片上,滴加0.1%的TTC进行染色,每个时期的花粉制2~3个玻片,每片取3个视野,分别统计不同时期花粉着色的百分率,求平均值作为该时期的花粉活力。
在盛花期10:00,选择开花前4、3、2、1 d的蕾和开放当天的花朵采集柱头,参照刘林德等[21]的方法检测柱头的可授性。
1.4杂交指数(Outcrossing Index,OCI)的估算 按照Dafni等[22]的标准进行花朵直径、花朵大小及开花行为的测定,由OCI值评判繁育系统类型。
1.5花粉/胚珠(Pollen-ovule Ratio,P/O)的测定 参照关文灵等[20]的方法测定单花花粉量和胚珠数,最后得到P/O值。依据Cruden[23]的标准评判繁育系统类型。
1.6坐果率的测定 参照关文灵等[20]的方法对金娃娃萱草分别进行6种处理,其中去雄套袋异株授粉(因为金娃娃萱草通过无性系繁殖,所有试验材料均为同一母本视为同株,所以异株授粉是授不同品种材料的花粉。每种处理取30朵花,在大蕾期去除雄蕊并套袋,开花当天10:00―10:30人工授粉。
2 结果与分析
2.1花部特征、花期物候和开花动态
2.1.1花部特征 金娃娃萱草为多年生宿根草本,具短根状茎和粗壮的纺锤形肉质根。叶基生、宽线形、对排成两列,叶宽0.96 cm,叶长28.42 cm,背面有龙骨突起,嫩绿色。花葶细长坚挺,高37.20 cm,着花4~8朵,呈顶生聚伞花序(图1A)。初夏开花,金黄色,花大,漏斗形,花筒长7.40 cm,花冠直径6.50 cm,花瓣6枚,分为内外两层,每层3枚花被片,围绕花柱和花丝呈镊合状排列(图1B),外花被裂片长圆形,顶端反卷,边缘无皱缩,长7.12 cm,宽1.41 cm(图1C a),内花被裂片卵形,顶端反卷,边缘皱缩,长7.12 cm,宽3.00 cm(图1C b),内外花被片基部合成花被筒。花柱长7.52 cm,柱头表面呈乳突状,成熟时分泌粘液。6枚雄蕊长短不一,花丝长4.52~5.10 cm,花药长0.52 cm,黄色(图1D)。子房上位,纺锤形(图1E),长0.52 cm,宽0.31 cm,绿色,坐果后果实呈嫩绿色,果长1.68 cm,宽1.56 cm(图1F),蒴果,背裂,内有亮黑色种子数粒(图1G)。
2.1.2花期物候 在人工栽培条件下,金娃娃萱草5-9月开花,一年开花3~4次(图1H),其中首次开花,花期最长,花量最大,其花期因每年气候变化略有差异。金娃娃萱草2010年首次抽苔期为5月1日(抽苔植株比例低于15%);盛抽期为5月4日(抽苔植株比例达50%以上);始花期为5月12日(开花植株比例低于15%);盛花期为5月17日(开花植株比例达50%以上),持续时间21 d;末花期为6月7日(开花植株比例低于15%);群体花期共持续32 d,开花高峰出现在5月中旬;单花花期为1 d,单个花苔的花序上花朵的开放无固定顺序。单个花苔从第1朵花开放到最后一朵花凋谢,持续9~16 d不等,这与植株的生长情况、单苔花朵数量以及天气情况有关。
2.1.3开花动态 晴天花朵的开放高峰在08:30―09:30,花朵开放时基部最外层花被裂片首先张开,之后花被裂片按由外向里的顺序依次张开,雌蕊首先显现出来,随着花被裂片不断向外展开,雄蕊也慢慢露出来,晴天时花冠未完全张开时花药就开裂散粉,如遇阴天花朵完全开放后也不会及时开裂散粉。在开花的整个过程中,雄蕊始终紧密围绕在花柱周围,花药上部距离柱头顶端约2.71 cm(图1B),雌蕊始终高于雄蕊,从而不利于自花传粉。金娃娃萱草花朵仅开放1 d,开花当天后期,首先花瓣开始失水变软,花冠部内外层花被片均向中心紧缩,花冠逐渐闭合并包裹花柱、花丝及花药,第2天早上花冠已经完全关闭(图1I);花瓣继续失水,颜色变浅,最后内外层花被裂片与花柱、花丝及花药缠绕在一起,花关闭1 d后,内轮花被片、花柱、外层花被片和子房渐渐褪色失水(图1J),萎蔫干燥,最后多数凋落。授粉花朵的闭合过程与之相同,只不过授粉后的花被裂片脱落的时间要推迟2~3 d(图1K),这也许和子房的生长发育有关。在栽培试验中观察到金娃娃萱草的传粉昆虫很少,在盛花期传粉昆虫主要有蜜蜂和菜粉蝶两种(图1M、N),但是其访花频率较低而且停留时间都很短,尤其是菜粉蝶,仅停留1~2 s。

图1 金娃娃萱草及该花昆虫Fig.1 Hemerocallis hybridus cv.‘Stella de oro’ and its insect
注:A,花序;B,花被片排列状态;C,内外花被片形状;D,花柱、花丝和花药;E,未受精子房;F,果实;G,种子;H,再次开花现象;I,第2天花朵闭合;J,花朵失水萎蔫脱落;K,子房膨大后花朵脱落;L,坐果后果实萎蔫失水;M,访花昆虫蜜蜂;N,访花昆虫菜粉蝶。
Note:A,Inflorescence; B,Tepals arrangement state; C,Shapes of internal and external tepals; D,Style,filament and anther; E,Unfertilized ovary; F,Fructification; G,Seed; H,Second flowering phenomenon; I,The next day of the flowers closed; J,The flower water loss,wilting then fall off; K,The flowers fall off after the ovary swollen; L,The fruit water loss and wilting; M,Bee; N,Cabbage butterfly.
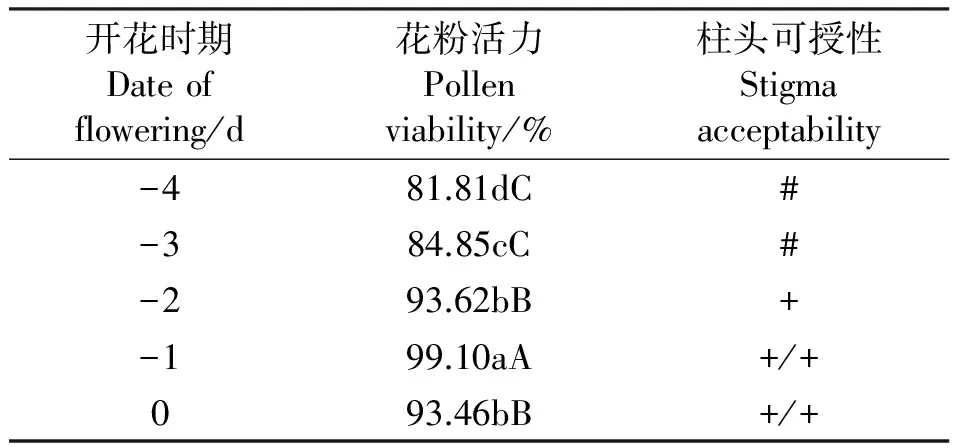
表1 金娃娃萱草的花粉活力和柱头可授性Table 1 Pollen viability and stigma acceptability of ‘Stella de oro’
注:数字前的“-”表示开花前; 0表示开花当天;“#”表示柱头不具有可授性;“+”表示柱头具可授性;“+/+”表示柱头具有强可授性;“+/#”表示柱头仅具部分可授性。同列不同小写和大写字母分别表示差异达0.05和0.01显著水平。
Note:“-” mean before blossomming ;“0” mean the blossoming day;“#” mean no stigma acceptability;“+” mean normal stigma acceptability;“+/+” mean high stigma acceptability;“+/#” mean part stigma acceptability. The same column with different letters mean, significant difference at 0.05 (lower case) and 0.01 (uppercase) levels.respectively.
2.2花粉活力和柱头可授性 金娃娃萱草在开花前大蕾期的花粉就已经具有活力(表1),开花前4 d花粉活力为81.81%,之后随着临近开放花粉活力逐渐增强,开花前1 d花粉活力最强,为99.10%,开花当天花粉活力为93.46%。由此可见,金娃娃萱草的花粉活力以开花前1 d为界呈现先逐渐增强后逐渐减弱的趋势,其中开花当天、开花前1 d和开花前2 d的花粉活力最强,是取花粉的最佳时期。金娃娃萱草开花前3、4 d的花蕾不具可授性,开花前2d的花蕾柱头具有可授性,柱头变蓝需2min左右;
开花前1 d和开花当天的柱头具有较强的可授性,柱头变蓝仅需5 s。由此可见,萱草开花前1 d和开花当天是给柱头授粉的最佳时期。
2.3杂交指数的估算 金娃娃萱草花朵直径为59.5 mm,大于6.0 mm,OCI值记为3,该物种属两性花,开花时柱头与花药在空间位置上是分离的,柱头位置比花药高,OCI值记为1;花药散粉初期雌蕊已经成熟,因此认为其雌、雄器官在成熟时间上是一致的,其OCI值记为0,所以金娃娃萱草的OCI值为4,根据Dafni等[22]的标准,其繁育系统为部分自交亲和,异交需要传粉者。
2.4花粉/胚珠 金娃娃萱草的单花花粉数平均为41 905 粒,胚珠数为36个,P/O值为1 164。根据Cruden[23]提出的标准,金娃娃萱草属于兼性异交类型。
2.5坐果率 授粉5 d后观察,坐果的果实呈浅绿色,子房开始膨大(图1F),未坐果的子房与花朵一起脱落(图1J);分别统计各处理的坐果率(表2)。试验结果表明,金娃娃萱草不存在无融合生殖现象;另外金娃娃萱草自然授粉中存在部分子房前期膨大,之后逐渐失水萎蔫的现象(图1L),造成这种现象的原因可能是自然授粉过程中接受的是异株花粉,两者杂交不亲和,但是据观察在9月份种质资源圃内只有金娃娃萱草处于花期,同样存在胚败育的现象。这说明金娃娃萱草存在一定的自交不亲和性,造成了后期不育。上述结果验证了该物种的繁育系统为兼性异交型,存在一定的自交不亲和性,属于部分自交亲和。

表2 金娃娃萱草授粉试验结果Table 2 The results of pollination of ‘Stella de Oro’
3 讨论
3.1金娃娃萱草繁育系统的判断 从萱草的花部特征来看其具有满足异交的诸多条件,如花朵硕大、数量繁多以及花色鲜艳等。依据Cruden[23]的评判标准,金娃娃萱草的繁育系统属于兼性异交类型;按照Dafni等[22]建议的标准测定杂交指数,金娃娃萱草的繁育系统为异交,部分自交亲和,需要传粉者;经人工授粉,金娃娃萱草为异交,需要传粉者,部分自交亲和。可见,由花粉/胚珠、杂交指数、授粉试验3种方法检测到的萱草繁育系统结果一致,因此认为金娃娃萱草的繁育系统类型属于兼性异交,需要传粉者,部分自交亲和。本试验坐果率研究结果表明自然授粉坐果率为14.0%,而且存在坐果后期部分胚败育的现象,这与崔贵梅等[24]研究结果金娃娃萱草平均每株结实率小于5%,存在大量无胚囊或胚囊内核发育紊乱及解体的败育现象一致。本研究中自然授粉坐果率高于5%,这也许和南北气候因素有关。
3.2金娃娃萱草的传粉系统 传粉系统由3个部分组成,即花粉、柱头和传粉媒介[25]。对于有性繁殖的植物,大量的花粉、有效的传媒和处于可授期的柱头是有效传粉的3个必要条件,因此要求植物花的生物学特征必须与其传粉机制相适应。本试验结果可知,金娃娃萱草的花粉活力和柱头可授性都没有问题。至于传粉媒介,由于在花朵开放的过程中柱头始终位于雄蕊之上,因此这种花部结构对花粉的传播与扩散十分不利。萱草的整个花期传粉昆虫很少,访花频率较低,停留时间也较短。可能因为萱草种质圃和松果菊、山桃草、金鸡菊等毗邻,传粉昆虫均被其他花吸引,较少在萱草上驻足停留。萱草种质圃和松果菊(Echinaceapurpurea)、山桃草(Gauralindheimeri)、金鸡菊(Coreopsisbasalis)等毗邻,传粉昆虫均被其他花吸引,较少在萱草上驻足停留。
自然界中两性花植物通常产生比果实或种子更多的花,结实率低的原因有几种:花粉限制、资源限制、缺少传粉昆虫,或者几种原因兼而有之[25]。该研究的试验材料为大田中栽培的园艺品种金娃娃萱草,种质圃内种质资源相当丰富,土壤、水肥等条件均较好,花粉量也十分充足,因此不存在花粉和资源的限制。综上分析得出,构成其坐果率低的原因如下:一是花器构造不利于花粉的传播与扩散;二是萱草花开于夏季,此时很多其他科属的植物同时开放,据观察发现一些传粉昆虫迷恋于周边菊科植物,而很少光顾萱草,分析可能是萱草本身花香味或花粉不受昆虫偏爱等原因造成,因此缺少传粉者;三是金娃娃萱草的单花花期短,仅有1 d,在传粉昆虫较少的情况下,这无疑进一步降低了其被授粉的机率。
3.3萱草的生殖补偿机制 为了保证后代的生存,生物往往发展出一整套必要的生殖补偿机制,保证物种繁衍的顺利进行[26]。萱草的主要繁殖方式为分株繁殖和播种繁殖,是一种无性繁殖和有性繁殖并存的植物。萱草自然结实率较低,因此以其根茎进行旺盛的无性繁殖,确保了其种群的繁衍。另外,萱草育种工作者,为了培育出更多综合性状优良观赏价值高的新品种,通过人工授粉不断创造新的种质。
[1] 汪发赞,唐进.中国植物志(第十四卷)[M].北京:科学出版杜,1980:52-62.
[2] 熊治廷,陈心启,洪德元.中国萱草属数量分类研究[J].植物分类学报,1997,35(4):311-316.
[3] Tomkins J P,Wood T C,Bames L S,etal.Evaluation of genetic variation in the daylily (Hemerocallisspp.) using AFLP markers[J].Theoretical and Applied Genetics,2001,102(4):489-496.
[4] 何琦,高亦珂,高淑滢.萱草育种研究进展[J].黑龙江农业科学,2011(3):137-139.
[5] 肖宜安,何平,李晓红.濒危植物长柄双花木的花部综合特征与繁育系统[J].植物生态学报,2004,28(3):333-340.
[6] 孔红,王庆瑞.甘肃萱草属植物的核型研究[J].广西植物,1993,13(3):247-251.
[7] 熊治廷,陈心启,洪德元.国产萱草属夜间开花类群的分类研究[J].植物分类学报,1996,34(6):586-591.
[8] 李洁,张少艾.萱草属HemerocallisL.若干野生种园艺品种染色体核型的比较研究[J].上海农学院学报,1995,13(3):208-217.
[9] 熊治延,陈心启,洪德元.萱草属中国特有种的细胞分类研究[J].植物分类学报,1997,35(3):215-218.
[10] 图力古尔,刘立波.吉林省3种萱草的核型研究[J].吉林农业大学学报,1995,17(3):50-55.
[11] 周朴华,何立珍,刘选明.组织培养中用秋水仙素诱发黄花菜同源四倍体的研究[J].中国农业科学,1995,28(1):49-55.
[12] 龙雅宜,龚维忠.多倍体萱草新品种的选育[J].园艺学报,1981,8(1):51-58.
[13] Panavas T,Pikula A,Reid P D,etal.Identification of senescence associated genes from daylily petals[J].Plant Molecular Biology,1999,40:237-248.
[14] Austin B.Engineering superior daylilies[J].American Nurseryman,1998,187(1):24-29.
[15] Li Z,Pinkham L,Campbell N F,etal.Development of triploid daylily (Hemerocallis) germplasm by embryo rescue[J].Euphytica,2009,169:313-318.
[16] Fogaca L A.Oliveira R A,Cuquel F L,etal.Heritability and genetic correlation in daylily selection[J].Euphytica,2011,7(22):1-10.
[17] 林辰壹,韩文娟,祁晨霞,等.新疆高葶韭雄蕊开花动态特征与花粉育性研究[J].草业科学,2012,29(4):561-566.
[18] 赵金花,李青丰,那仁图雅,等.3种野生葱属植物花粉活力和柱头可授性研究[J].草业科学,2010,27(4):93-96.
[19] Dafni A.Pollination Ecology:A Practical Approach[M].New York:Oxford University Press,1992:1-57.
[20] 关文灵,李叶芳,陈贤,等.蝴蝶花花器结构和开花授粉生物学特性[J].园艺学报,2009,36(10):1485-1490.
[21] 刘林德,张洪军,祝宁,等.刺五加花粉活力和柱头可授性研究[J].植物研究,2001,21(3):376-380.
[22] Dafni A,Hesse M,Pacini E.Polen and Pollination[M].New York:Springer-Verlag,2000.
[23] Cruden R W.Intraspecific variation in pollen-ovule ratios,nectar secretion-preliminary evidence of ecotypic variation[J].Annals of Missouri Botanic Garden,1976,63(2):277-289.
[24] 崔贵梅,杨丽莉,孙毅,等.大花萱草‘金娃娃’雌雄蕊发育进程及其与花蕾长度的相关性研究[J].西北植物学报,2008,28(10):1983-1988.
[25] 黄双全,郭友好.传粉生物学的研究进展[J].科学通报,2000,45(3):225-237.
[26] 张丙林,穆春生,王颖,等.五脉山黧豆开花动态及有性繁育系统的研究[J].草业学报,2006,15(2):68-73.
StudiesonflowerstructureandbreedingsystemofHemerocallishybriduscv. ‘Stelladeoro’
ZHAO Tian-rong, CAI Jian-gang, SHI Yong-tai, NI Jian-gang, SHEN Lan
(Ningbo Academy of Agricultural Sciences, Ningbo 315040, China)
A field investigation, including continuous observation, the use of outcrossing index, pollen and stigma viability; pollen-ovule ratio and fruit-set rate, was conducted on studing flower structure and breeding system ofHemerocallishybriduscv. ‘Stella de oro’. The main results were that under artificial culture conditions, the population bloomed three or four times a year from May to September. The first flowering span among populations was 32 days and for single scapes was 9-16 days. The life span of one single flower was 1 day. The stamen was close to the lateral side of the petaloid style and 2.71 cm shorter than the style. Relative position between the stamen and style was always unchangeable in the whole flowering process. The pollen viability was the strongest on the blooming day and the stigma had high stigma acceptability on the blooming day and 1 day before blooming. The outcrossing index was 4 and P/O was about 1 164. Combined those with the result of the bagging and artificial pollination experiment, the breeding system of the species was given priority to facultative outcrossing, part self-compatibility and insect pollinator dependent. The special floral structure, lack of pollinators and short florescence might be the reasons that the species had a low setting percentage.
Hemerocallishybriduscv. ‘Stella de oro’; floral syndrome; breeding system; outcrossing index; pollen-ovule ratio
ZHAO Tian-rong E-mail:rongronglily@163.com
2012-03-15接受日期:2012-05-18
宁波市科技攻关项目(2006C100035);宁波市农科教结合项目(2008NK31);宁波市农业科学研究院院长基金(2009-11)
赵天荣(1980-),女,河北沧州人,农艺师,硕士,主要从事观赏植物育种研究。E-mail:rongronglily@163.com
Q944.52
A
1001-0629(2013)01-0052-06
