人类Boule基因在减数分裂时期的生物信息学分析
2013-11-14孙洋洋马玉珍石晶瑜
孙洋洋,马玉珍,石晶瑜
(1.内蒙古师范大学,内蒙古呼和浩特010000;2.内蒙古医院,内蒙古呼和浩特010000)
在精子发生这一复杂的生理过程中,减数分裂时期显得尤为重要。染色体的复制、分离,基因突变和缺失都可能使精子发生异常,进而导致男性不育。在与男性不育[1]相关的众多调控因子当中,DAZ家族就是其中非常重要的调控因子。它包括三个基因成员:DAZ、DAZL、Boule。DAZ 位于 Y 染色体上[2],主要在灵长类动物中发现该基因;DAZL(Deleted in azoospermia-like)位于常染色体,在哺乳动物、脊椎动物中存在;Boule[3]基因也是位于常染色体上,在线虫、哺乳动物、果蝇及脊椎动物中存在,并且在第一次减数分裂时期高表达。Boule是一个高度保守的基因,它编码精子发生减数分裂所需要的RNA结合蛋白。Boule基因全序列为编码区起始于序列的118,终止于322,存在CpG岛,长度为205bp。编码区两侧为254bp的5’非翻译区和308bp的3’非翻译区,在3’末端的polyA尾上游处有一个加尾信号AATAAA。Kostova[4]发现 Boule在睾丸组织中特异性表达的三个转录剪接体,其N末端又由三个选择性剪接外显子编码。尽管调节精子发生的基因发生着迅速的进化,但是从昆虫到人类的Boule基因仍维持着保守的功能域[5],果蝇的Boule突变会使精子发生在减数分裂时期受阻从而导致雄性不育。Eugene Yu jun Xu[5]等人在因 Boule缺失或突变导致的雄性不育苍蝇睾丸中导入人的Boule基因转录组,发现雄性不育果蝇能产生正常的精子。人类与果蝇Boule基因相似的功能表明人的Boule基因与果蝇的Boule基因直系同源,并且在调节减数分裂上,它在人类中的功能很可能与果蝇类似。人类精子发生过程中,减数分裂M期的起始是受M期启动因子MPF控制的,而MPF由细胞周期蛋白B和CDC2(Cell division cycle 2)组成。这个复合物是恢复减数分裂必需的,其活性受可逆磷酸化的调控,Wee1和 Myt1激酶使其磷酸化可致其失活,而CDC25磷酸酶可以使其活化,诱导细胞周期G2/M转换的完成[6]。目前已发现3种人类CDC25基因,其中CDC25A在睾丸中高表达,对精子发生至关重要[7],特别是在初级和次级精母细胞中表达,与Boule蛋白的初始表达阶段一致。在Boule缺乏的患者中,CDC25A磷酸酶基因mRNA仍然表达,有的甚至出现上调,但并没有翻译成具有功能的蛋白,而CDC25A磷酸酶的缺乏可能导致MPF超磷酸化,使MPF完全失活,这可能是患者精子发生阻滞的主要原因,Boule和CDC25A同时缺乏的患者减数分裂发生阻滞。Boule基因在哺乳动物其它组织器官中的功能,尤其是它在精子发生中的所起的作用尚不完全清楚[9],但Boule是精子生成所不可缺少的一个基因,它的表达在精子发生过程中起关键作用。
1 材料及其方法
1.1 材料
人 (Homo sapiens,AF272858)、斑 胸 草 雀(Taeniopygia guttata,XM-004175306.1)、虹鳟鱼(Oncorhynchus mykiss,HQ696915)、曼氏裂体吸虫(Schistosoma mansoni,XP-002575519.1)、拟南芥(Arabidopsis thaliana,NM-111126.3)、黑腹果蝇(Drosophila melanogaster,NM-079265)、小鼠(Mus musculus, AF272859)、牛 (BosTaurus, NM-001102115)、松鼠猴 (Saimiri sciureus,AJ717408)、倭黑猩猩(Pan paniscus,AJ717405)等10种Boule基因的mRNA序列及其相应的氨基酸序列,均来自美国国家生物信息中心(NCBI,National center for biotechnology information) 网 址:http://www.ncbi.nlm.nih.gov/。物种分类如图1所示。

图1 10个物种的分类图Fig.1 The ten species investigated in this study
1.2 方法及其使用的软件
1.2.1 Boule基因的mRNA序列和蛋白序列分析
通过NCBI网站的Gene数据库、Nucleotide数据库和Protein数据库查询人、家蚕等10种Boule基因的mRNA序列、氨基酸序列,基因所在的染色体及其蛋白质质量。
1.2.2 Boule蛋白质序列比对分析
利用 EBI(网址:http://www.ebi.ac.uk)网站的Clustalw Omega(网址:http://www.ebi.ac.uk/Tools/msa/clustalo/),对10种动物的Boule蛋白的氨基酸序列做比对分析。
1.2.3 Boule蛋白分子进化分析
利用分子进化遗传分析软件MEGA5.05(Molecular evolutionary genetics analysis)对10种动物的全长Boule蛋白做序列比对分析。系统进化树采用邻接算法,自检举5 000次,采用泊松校验计算距离。
1.2.4 Boule蛋白的氨基酸的理化性质分析
利用 Expasy(网址 http://www.expasy.org/)的ProtParam工具 (网 址:http://web.expasy.org/protparam/)对人的Boule蛋白氨基酸序列的相对分子质量、氨基酸组成、等电点(PI)、不稳定系数等物理化学性质进行分析。
1.2.5 疏水性/亲水性的预测和分析
利用Expasy网站的protscle(网址:http://web.expasy.org/protscale/)在线分析工具对人类Boule基因编码的蛋白氨基酸序列的疏水性亲水性进行分析。所有的参数按照软件的默认值。
1.2.6 信号肽分析
对人类Boule蛋白氨基酸序列,利用在线分析工具SignalP 4.0 Server(网址:http://www.cbs.dtu.dk/services/SignalP/)分析该氨基酸序列的信号肽的存在位置及序列。
1.2.7 跨膜结构的预测和分析
跨膜结构域常常是由跨膜蛋白的效应区域所展现,一般由20个左右的疏水性氨基酸残基组成,主要形成α-螺旋。利用在线工具TMHMM Server v.2.0(网 址: http://www.cbs.dtu.dk/services/TMHMM/)对人类Boule蛋白进行跨膜结构分析。
1.2.8 卷曲螺旋域分析
在很多的天然蛋白质中,都存在卷曲螺旋,该结构具有重要的生物学功能,比如分子识别、运动和调控代谢等。利用在线分析工具COILS(http://www.ch.embnet.org/software/COILS-form.html)对 Boule蛋白进行卷曲螺旋域分析。
1.2.9 亚细胞定位分析
利用在线亚细胞定位工具TargetP-1.1 Server(http://www.cbs.dtu.dk/services/TargetP/),输入Boule基因编码的蛋白序列,在序列来源选择Nonplant,其它选项用默认值,确认Boule基因编码的蛋白主要在细胞中发挥作用的部位。然后再用PSORTⅡserver(http://psort.hgc.jp/form2.html/) 工具进一步分析来验证上述工具分析结果的准确性。
1.2.10 与Boule蛋白相互作用的蛋白分析
在STRING数据库中(http://string-db.org/)查询与Boule蛋白相关作用的蛋白。主要研究与Boule蛋白直接或者间接作用的相关蛋白。之所以研究其蛋白之间的相互作用关系,是因为可以通过与其相关蛋白的功能信息,了解和预测该蛋白的功能及其它信息。
1.2.11 Boule蛋白的二级结构预测
利用基于 GOR算法的 GOR4(http://npsapbil.ibcp.fr/cgi- bin/npsa-automat.plpage=npsagor4.html)在线工具预测该蛋白是否具模体、无规则卷曲、β转角、β折叠及α螺旋等结构。进一步用PredictProtein软件对Boule的二级结构进行分析。
1.2.12 Boule蛋白的功能域预测和分析
分析蛋白质的组成及其中所包含的功能域是了解蛋白质功能的重要一步。蛋白质中的功能域是参加蛋白质之间相互作用的结构和功能单位。利用Pfam数据库(http://pfam.janelia.org/search/) 检测蛋白功能域的组成,主要对其进化过程中的高度保守的特异性区域进行分析。
2 结果分析
2.1 Boule基因的mRNA序列和蛋白序列分析
从NCBI网站相应的数据库中得到10种动物的全长Boule蛋白序列和 mRNA序列(见表1)。不同动物Boule基因的蛋白质大小不同,位于常染色体上。曼氏裂体吸虫的Boule蛋白最大,为509 aa;虹鳟鱼最小,为194 aa;小鼠的 Boule蛋白是297 aa;人、松鼠猴、倭黑猩猩的Boule蛋白质大小相等,均为283 aa;牛的Boule蛋白大小是295 aa;黑腹果蝇的Boule蛋白大小为228 aa;野猪则为293 aa。

表1 10种不同动物Boule基因查询信息Table 1 Information of Boule genes in 10 different animals
2.2 Boule蛋白质序列比对分析
对10种动物的Boule蛋白氨基酸序列做比对分析(见表2)。哺乳动物类人、鼠、牛、松鼠猴、倭黑猩猩之间的Boule蛋白序列的相似度很高,在90% ~99%之间。其中人与倭黑猩猩的Boule蛋白相似度很高,高达99%。哺乳动物与其它类动物Boule蛋白序列的相识度比较低,在31% ~52%之间,其中哺乳动物与黑腹果蝇的Boule蛋白序列相似度为31%~37%之间。曼氏裂体吸虫与拟南芥Boule蛋白序列相似度最低仅为1%,表明了Boule基因在进化中其功能的高度保守性。
表2 不同物种的Boule蛋白全长序列多序列比对的结果Table 2 Mutiple sequence alignment of Boule protein in different animals
2.3 Boule蛋白分子进化分析
由分子进化图(见图2)中可以看出,Boule蛋白在这10个物种中,主要分为两大类。拟南芥独自一类,其它9个物种为一大类;在哺乳动物中,很明显的又分为两类,小鼠、牛、松鼠猴为一类,人、黑猩猩为一类并且亲缘关系也最近。鸟类,鱼类与哺乳动物的亲缘关系也比较近。
2.4 Boule蛋白的氨基酸的理化性质分析
Boule蛋白分析表明:Boule蛋白含有283个氨基酸,分子质量为31 299.86 Daltons,含量最丰富的氨基酸为 Pro、Ser、Val等(见表 3)。
2.5 疏水性/亲水性的预测和分析
蛋白质能够折叠,主要因为其具有亲水性/疏水性的氨基酸。对其疏水性和亲水性进行分析可以更好地了解蛋白质。该蛋白的疏水区共有9处(图3),分别为 33~36,38~40,123~128,167~169,171~173,205 ~207,232 ~234,237~238,265 ~267。亲水区共有15处,分别为5~8,18~19,21~25,29~30,42~54,57 ~59,69 ~72,81 ~91,94 ~104,108 ~111,114 ~116,135 ~142,157~161,176 ~199,241 ~252。其中第168位最高,值为1.733;第98位最低,值为-2.622。P氨基酸分值最高表明其疏水性最强,Y氨基酸分值最低表明其亲水性最强。纵观Boule蛋白,氨基酸序列存在明显的亲水区和疏水区。

图2 用邻接法(NJ)构建的10种动物Boule蛋白的系统进化树Fig.2 Phylogenetic tree of Boule protein sequence from 10 different animals

表3 boule蛋白的氨基酸理化性质Table 3 Physicochemical properties of boule protein
2.6 信号肽分析
在核糖体合成蛋白后,往往需要到达细胞中制定的位置才能发挥其功能。而蛋白能否到达细胞中的特定位置,则完全由信号肽进行引导。经SingnalP 4.0预测Boule蛋白的信号肽序列,结果见图4。由图4可知,Boule蛋白不存在信号肽,属于非分泌型蛋白质。与 PredictProtein数据库(http://www.predicprotein.org)分析的结果一样。

图3 Boule蛋白氨基酸序列的疏水性/亲水性预测Fig.3 Hydrophobicity/phydrophilicity prediction of the Boule protein
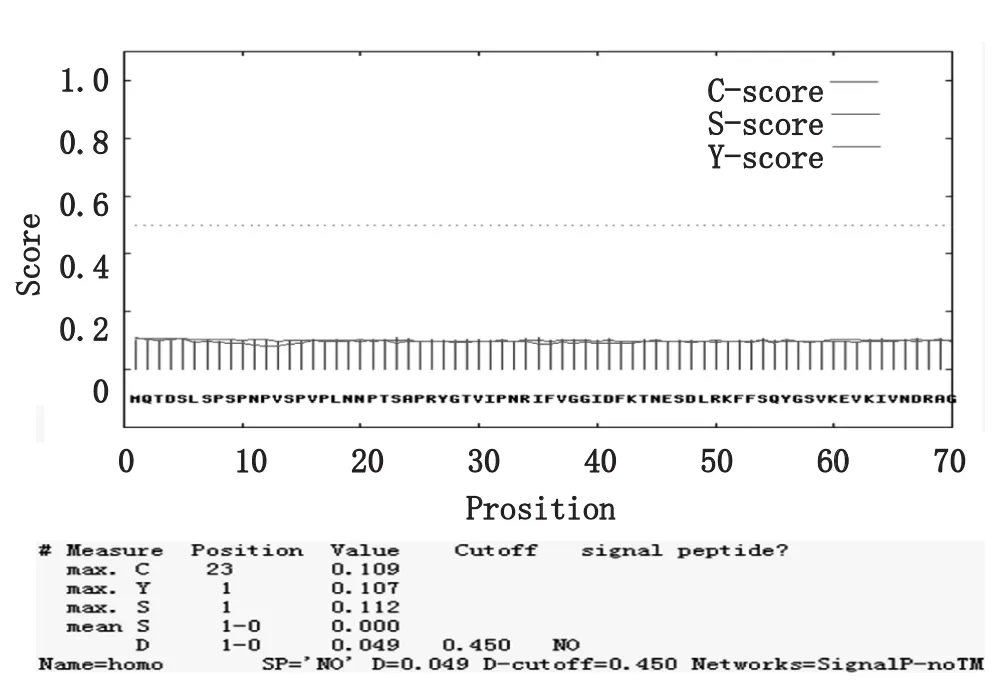
图4 Boule蛋白信号肽预测Fig.4 Signal peptide prediction of the Boule protein
2.7 跨膜结构的预测和分析
用在线工具TMHMM-2.0对Boule蛋白进行跨膜结构分析如图所示(见图5),结果表明Boule蛋白跨膜的可能性为0,所以Boule蛋白不存在跨膜结构域,属于非跨膜蛋白质类。

图5 Boule蛋白跨膜区域预测Fig.5 Transmembrane domain prediction of the Boule protein
2.8 卷曲螺旋分析
卷曲螺旋存在于很多天然的蛋白质当中,由图6所示,在窗口为14和21的时候,在75~100处可能形成卷曲螺旋。

图6 Boule蛋白的卷曲螺旋预测Fig.6 Coiled - coil prediction of the Boule protein
2.9 细胞定位分析
如图7所示,Boule蛋白基本上排除了在分泌途径上的功能,与信号肽分析的结果一致。与日本的PSORTⅡserver图8比较,Boule蛋白在细胞质、细胞核和线粒体内表达。文献报道,Boule蛋白在第一次减数分裂(前中期)的生殖细胞中表达,从细线期精母细胞(Leptotene spermatocytes)开始表达,到偶线期精母细胞(Zygotene spermatocytes)表达量增加,在粗线期精母细胞(Pachytene spermatocytes)表达量达到最高水平,到双线期(Diplotene)表达量显著下降,在圆形精子细胞(Round spermatids)、长形精子细胞(Elongating spermatids)等精子发生更高级阶段的细胞中Boule不表达,在胚胎早期、原始生殖细胞(Primordialgerm cells)和精原细胞(Spermatogonial cells)中均未检测到Boule蛋白的表达[9],Boule蛋白在细胞质中特异表达在细胞核或接近减数分裂的染色体中未见表达。人类精子发生过程中,减数分裂M期的启始受减数分裂启动因子MPF控制,而MPF是一个由细胞周期蛋白 B和Cdc2(Cell division cycle 2)组成的复合物,这个复合物对减数分裂的恢复是必需的,其活性受可逆磷酸化的调控,Wee1和Myt1激酶使MPF磷酸化可致其失活,而Cdc25磷酸酶可以使MPF活化,诱导G2/M转换的完成。

图7 Boule蛋白的亚细胞定位Fig.7 Subcellular localization prediction of Boule protein
图8 Boule蛋白的细胞定位Fig.8 Cellular localization prediction of Boule protein
2.10 与Boule蛋白相互作用蛋白分析
每一种蛋白质并不是孤立地存在于细胞中,而是与其他蛋白质相互作用形成复合体或功能模块,从而在细胞中进行各种不同的生理活动(见图9)。在果蝇中,PUM2蛋白与 Boule蛋白相互作用[10]。Boule激活并调控CDC25A的表达并且促进CDC25AmRNA的翻译,CDC25A蛋白进一步促使MPF中的Cdc2磷酸化,激活促成熟因子(Maturation-promoting factor,MPF),活化的MPF启动减数分裂过程G2PM的转换,完成减数分裂过程。Boule蛋白的缺失或突变可能导致雄性不育。
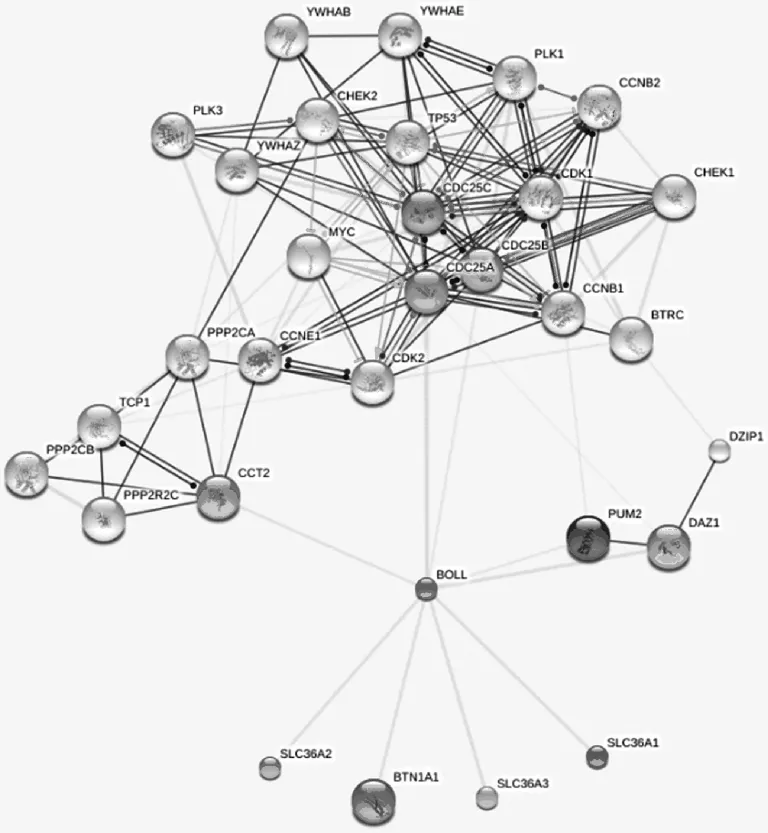
图9 Boule蛋白相互作用的蛋白分析Fig.9 Analysis of protein interaction with Boule
2.11 Boule蛋白的二级结构预测
该蛋白由 9.19% α-螺旋(Alpha helix)、22.26%延伸链(Extended strand)和68.55%无规则卷曲(Random coil)所组成。进一步用PredictProtein软件对Boule的二级结构进行分析,螺旋=7.4%,股=8.8%,环=83.7%,非正规二级结构区:92-283(见图10)。
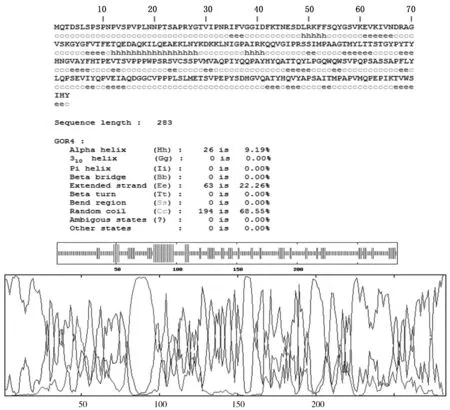
图10 Boule蛋白的二级结构预测Fig.10 Secondary structure prediction of the Boule protein
2.12 Boule蛋白的功能域预测和分析
在35位点开始,106位点结束,存在RRM(RNA recognition motif)保守域(见图11),说明Boule基因在哺乳动物中具有RRM基序、编码RNA结合蛋白的特性[11]。

图11 Boule蛋白结构域预测Fig.11 Predicted the structure domain of Boule protein
RRM是RNA结合蛋白中广泛存在的一种保守结构,它存在很多不同的RNA结合蛋白中,如异质性核糖蛋白(Heterogeneous nuclear ribonucleoproteins(hnRNPs)),在RRM基序中含有许多保守的氨基酸以保证对RNA的结合活性,但是这一家族的不同蛋白质却能特异地结合各种不同的RNA分子。
另外,运行blastp程序服务器对Boule蛋白进行搜索,得到图12。从图中看到,Boule蛋白属于RRM(RNA recognition motif)超家族,具有与 COG0724,PABP-1234,PLN03134,SF-CC1,U2AF_1g,ELAV_HUD_SF,half_pint,hnRNP-R-Q 相似的保守域。

图12 Boule蛋白结构域预测Fig.12 Predicted domain site of Boule protein
3 讨论
Boule是果蝇、哺乳动物不育和雄性生殖细胞在阻滞在减数分裂时期的一个高度保守的关键调控因子。在与精子发生障碍相关的不育病人当中,Boule蛋白的表达显著的下降。Boule是DAZ家族的一个成员,存在于灵长动物、哺乳动物和后生动物中。在果蝇中,研究表明,Boule表达在第一次减数分裂时期,调节Twine(phosphatase of Cdc25)的表达并开始促进成熟细胞因子(initiate MPF),比如Cdc2/cyclin B复合物,这些都是G2-M转换和第一次减数分裂时期的关键因子。所以在果蝇中,Boule调节Twine来控制减数分裂。调节生殖细胞的通路似乎很大程度上是保守的。本文对Boule基因及其编码蛋白信号肽、二级结构、亲疏水性、结构域及同其它物种的蛋白相似度同源比对等进行了生物信息学分析。结果表明,该蛋白无信号肽,无跨膜结构是亲水性蛋白,在75~100处可能形成卷曲螺旋。二级结构以α-螺旋为主。具有RRM功能域。Boule蛋白的同源性分析证明了Boule功能在动物中是高度保守的[12]。大量的实验已经表明Boule在精子形成减数分裂时期具有重要的调节作用。Xu等人发现Boule仅仅在精子发生时是必需的,但是在雌性的Boule基因缺失或者突变的时候仍可以正常生育。
References)
[1] Michael A.Crackower,Nadine K.Kolas,Junko Noguchi,Renu Sarao, Kazuhiro Kikuchi, Hiroyuki Kaneko, Eiji Kobayashi,Yasuhiro Kawai,Ivona Kozieradzki,Rushin Landers,Rong Mo,Chi-Chung Hui,Edward Nieves,Paula E.Cohen,Lucy R.Osborne,Teiji Wada,Tetsuo Kunieda,Peter B.Moens,Josef M.Penninger.Essential role of Fkbp6 in male fertility and homologous chromosome pairing in meiosis[J].Science,2003,300(5623):1291-1295.
[2] Renee Reijo,Tien-Yi Lee,Pia salo,Raaji Alagappan,Laura G.Brown,Michael Rosenberg,Steve Rozen,Tom Jafe,Donald Straus,Outi Hovatta,Albert de la Chapelle,SherMan Silber,David C.page.Diverse spermatogenic defects in humans caused by Ychromosome deletions encompassinganovelRNA2 binding protein gene[J].Human Reprouction,1996,11(4):27 -54.
[3] Eugene Yujun Xu,Frederick L.Moore,Renee A.Reijo Pera.A gene family required for human germ cell development evolved from an ancient meiotic gene conserved in metazoans[J].Proceedings of the National Academy of Sciences of the United States of America,2001,98(13):7414-7419.
[4] Yung Ming Lin,Yen Ni Teng,Chia Ling Chung,Wan Ching Tsai,Ying Hung Lin,Johnny Shinn Nan Lin,Pao Lin Kuo.Chung C L,et al.Decreased mRNA transcripts of M2phase promoting factor and its regulators in the testes of infertile men[J].Human Reproduction,2006,21(1):138-144.
[5] Eugene Yujun Xu,Douglas F.Lee,Ansgar Klebes,Paul J.Turek,Tom B.Kornberg,Renee A.Reijo Pera.Human boule gene rescues meiotic defects in infertile flies.Human Molecular Genetics..,2003,12(2),169 -175.
[6] Debra J.Wohlgemuth,Erika Laurion,Karen M.Lele.Regulation of the mitotic and meiotic cell cycles in the male germ line [J].Recent Progress in Hormone Research,2002,57(1):75 -101.
[7] Mei Shya Chen,Jonathan Hurov,Lynn S.White.Absence of apparent phenotype in mice lacking Cdc25C protein phosphatase[J].Molecular and Cellular Biology,2001,21(12):3853 -3861.
[8] Michael J.W.VanGompel,Eugene Yujun Xu,A novel requirement in mammalian spermatid differentiation for the DAZ-family protein Boule,Human Molecular Genetics[J],2010,Vol.19,No.12 2360 -2369.
[9] Nicola Reynolds,Howard J Cooke.Role of the DAZ genes in male fertility[J].Reproductive Biomedicine Online,2005,10(1):72-80.
[10] Kiran Padmanabhan,Joel Richter.Regulated Pumilio -2 binding controls RINGO/Spy mRNA translation and CPEB activation[J].Genes& development.2006;20(2):199-209.
[11] Douglas W.Houston,Jian Zhang,Jean Z.Maines,Steven A.Wasserman SA,Mary Lou King.Xenopus DAZ-like gene ecodes an RNA component of germ plasm and is a functional homologue of Drosophila Boule.Development,1998 125(2):171 -180.
[12] Gerald J.Wyckoff,Wen Wang,Chung-I Wu,Rapid evolution of male reproductive genes in the descent of man[J].Nature,2000,403(6767),304-309.
