不同光照和温度下白花鬼针草和金盏银盘种子萌发研究
2013-10-24韦春强潘玉梅唐赛春林春华刘明超
韦春强, 潘玉梅, 唐赛春, 林春华,2, 刘明超,2
(1.广西壮族自治区中国科学院广西植物研究所,广西桂林 541006; 2.广西师范大学,广西桂林 541004)
不同光照和温度下白花鬼针草和金盏银盘种子萌发研究
韦春强1, 潘玉梅1, 唐赛春1, 林春华1,2, 刘明超1,2
(1.广西壮族自治区中国科学院广西植物研究所,广西桂林 541006; 2.广西师范大学,广西桂林 541004)
通过在人工培养箱内模拟环境条件,探讨了不同光照和温度对白花鬼针草和金盏银盘种子萌发的影响,结果表明:光照和温度不是白花鬼针草种子萌发的必要条件,白花鬼针草种子萌发的温度适应范围比金盏银盘宽,在15 ℃/10 ℃、20 ℃/15 ℃、25 ℃/20 ℃、30 ℃/25 ℃、35 ℃/30 ℃、40 ℃/35 ℃下均能萌发,在 15 ℃/10 ℃、20 ℃/15 ℃、25 ℃/20 ℃、30 ℃/25 ℃时萌发率均达94%左右,金盏银盘种子在40 ℃/35 ℃时不能萌发;温度影响白花鬼针草和金盏银盘种子萌发高峰的出现时间及峰值大小,而光照不影响。
白花鬼针草;金盏银盘;种子萌发;光照;温度
白花鬼针草(BidensalbaL. )为菊科(Compositae)鬼针草属(BidensL. )一年生草本,原产于热带美洲,现在广泛分布于世界各地的热带和亚热带地区,在我国主要分布于广西、广东、海南、云南和贵州等地[1-2]。白花鬼针草在美国被认为是危害最严重的世界性杂草之一[3],在我国也是分布范围较广、危害较大的外来入侵杂草之一,主要入侵农田、果园、林地等生境,容易形成单一群落,对农林业生产以及生物多样性带来极大危害。目前对白花鬼针草的研究较少,主要有形态区分[4]、特征特性[1]、化感作用[5]和生理特性[6]方面的研究,关于其种子萌发方面的研究未见报道。金盏银盘(B.biternataL.)是白花鬼针草近缘同属本地种,秋季开花,一般花期较短,生境也较为广泛,路边、田边、果园甚至林下均可见,但数量不多,一般是单株或几株零星分布,未造成危害[7]。
将入侵种与近缘本地种进行比较研究是研究植物入侵性的一个重要方法,可以排除亲缘关系相差较远或进化水平不同等因素的影响,使结果更加具有说服力[8-10]。种子萌发是某一物种从潜在种群转变为现实种群的关键,对种群个体的定居、繁殖、扩散和抵御不良环境有着非常重要的意义。本研究通过比较不同光照和温度条件下入侵植物白花鬼针草与其近缘本地种金盏银盘的种子萌发,为评价白花鬼针草扩散到新生环境下是否能定居并最终形成入侵提供了可靠的依据。
1 材料与方法
1.1 材料
白花鬼针草种子于2010年11月采自广西壮族自治区横县,金盏银盘的种子于2010年8月采自广西壮族自治区桂林地区,成熟种子采集后装入纸袋,常温保存。
1.2 方法
2011年3月开始进行白花鬼针草和金盏银盘种子在不同光照和温度条件下的萌发试验。参照Etejere等的研究[11-12],设置2个光照条件,各6种不同变温,共12个处理,每个处理4次重复。每个处理选取40粒饱满种子,置于底部垫有2层滤纸的培养皿中进行试验,并保持培养皿中滤纸湿润,将其置于人工培养箱中萌发。萌发过程中每天观测1次,连续观察20 d,如果20 d后仍有萌发则观察至萌发结束为止。萌发率计算:萌发率=发芽种子数/试验种子数×100%。
光照设置周期性光照和全黑暗2种。周期性光照设置时间为13 h/11 h(昼/夜),光照强度为 8 000 lx,萌发温度设置为随昼/夜(13 h/11 h)而变换,分别是40 ℃/35 ℃、35 ℃/30 ℃、30 ℃/25 ℃、25 ℃/20 ℃、20 ℃/15 ℃、15 ℃/10 ℃等6种不同变温。全黑暗用2层黑布盖住培养皿,使其保持 24 h 持续黑暗,萌发温度设置同周期性光照。
1.3 数据处理
采用Excel软件对数据进行整理、计算、绘图,用平均数代表样本各性状的整体水平;用标准差反映样本的绝对变异度,采用SPSS统计软件的One-way ANOVA分析各参数的差异。
2 结果与分析
2.1 不同光照和温度对白花鬼针草和金盏银盘种子萌发率的影响
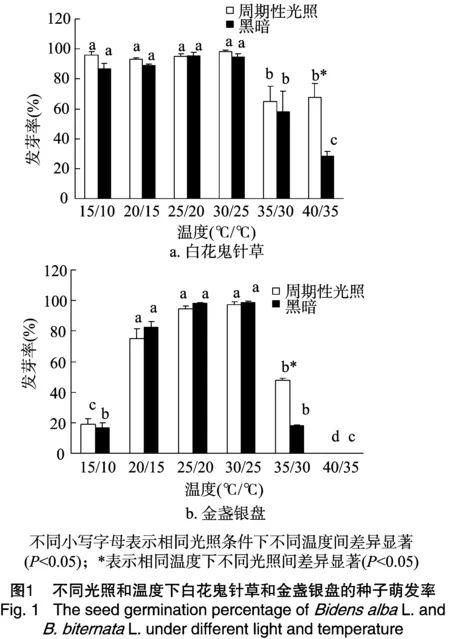
从图1可以看出,在周期性光照和黑暗条件下,白花鬼针草种子在15 ℃/10 ℃、20 ℃/15 ℃、25 ℃/20 ℃、30 ℃/25 ℃时均有较高的萌发率,达94%左右;当温度为35 ℃/30 ℃时,2种条件下白花鬼针草种子萌发率为60%左右,均出现显著下降(P<0.05);当温度到达40 ℃/35 ℃时,周期性光照条件下白花鬼针草种子萌发率与35 ℃/30 ℃时无显著差异(P>0.05),但是黑暗条件下与35 ℃/30 ℃时存在显著差异(P<0.05)。在周期性光照和黑暗条件下,在15 ℃/10 ℃、20 ℃/15 ℃、25 ℃/20 ℃、30 ℃/25 ℃ 范围内,金盏银盘种子萌发率随着温度的升高而升高,最高时达到98%左右;当温度为 35 ℃/30 ℃ 时,2种条件下金盏银盘种子萌发率均出现显著下降(P<0.05),且2种条件下金盏银盘种子萌发率也存在显著差异(P<0.05);当温度到达40 ℃/35 ℃时,2种条件下金盏银盘种子萌发率均为0。
2.2 不同光照和温度对白花鬼针草和金盏银盘种子萌发进程的影响
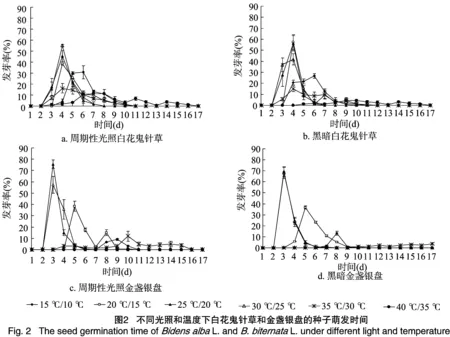
如图2所示,温度影响白花鬼针草和金盏银盘种子萌发高峰的出现时间及峰值大小,而光照不影响。在周期性光照和黑暗条件下,白花鬼针草种子萌发在温度为35 ℃/30 ℃、30 ℃/25 ℃、25 ℃/20 ℃、20 ℃/15 ℃下4 d时达到高峰期;温度为15 ℃/10 ℃时,处理6 d达到发芽高峰;而温度为40 ℃/35 ℃时种子萌发无明显高峰期,且其萌发持续时间较长,为16 d,其他温度条件下仅为8 d左右。在周期性光照和黑暗条件下,金盏银盘种子在温度为30 ℃/25 ℃、25 ℃/20 ℃时萌发迅速,而且发芽整齐,处理3 d时萌发达到最高峰,萌发率达55%以上;在温度为20 ℃/15 ℃时处理5 d萌发达到最高峰;在温度为15 ℃/10 ℃时,种子仅在处理7~10 d时萌发,且萌发率低;35 ℃/30 ℃时种子萌发无明显高峰期,而40 ℃/35 ℃时种子不萌发。
3 讨论
外来种的入侵过程一般认为包括引入、建群、扩散和危害等阶段[13],种群的建立及扩散是外来种是否成功入侵过程中很重要的阶段,种子是其中最为关键的因素之一,因而种子的萌发又是一个非常重要的环节。种子的萌发受所多种因素的影响,而光照和温度则是其中比较重要的因素。
通过研究发现,白花鬼针草在温度为15 ℃/10 ℃、20 ℃/15 ℃、25 ℃/20 ℃、30 ℃/25 ℃时均有较高的种子萌发率,达94%左右,而在温度为35 ℃/30 ℃时,仍有60%左右的萌发率。金盏银盘在温度为20 ℃/15 ℃、25 ℃/20 ℃、30 ℃/25 ℃时,种子萌发率达75%以上。洪岚等对外来入侵植物三叶鬼针草(BidenspilosaL.)种子萌发进行研究,发现三叶鬼针草种子萌发适宜温度为15~30 ℃[14]。杨期和等对分布于滇南热带的256种热带植物的种子萌发的影响因素进行了研究,结果表明在能萌发的125种植物中,有80%以上的植物种子萌发的适宜温度为18~30 ℃[15]。韦春强等研究发现飞机草(EupatoriumodoratumL.)种子萌发的适宜温度为20~30 ℃[16]。本研究的白花鬼针草和金盏银盘的种子萌发适宜温度与前人研究相似。当温度低于或高于白花鬼针草和金盏银盘的种子萌发的最适范围时,其萌发时间延长,这是它们对环境温度的适应策略之一。通过比较发现,白花鬼针草种子萌发的温度适应范围比金盏银盘广,在15~40 ℃时均能萌发,这可能是白花鬼针草能成为入侵种并广泛分布的一个重要原因。
不同植物种子的萌发对光的要求不同,可分为需光、忌光和中性3种[17],如飞机草、假苍耳(Ivaxanthifolia)、紫茎泽兰(Eupatoriumadenophorum)、薇甘菊(Mikaniamicrantha)等种子在没有光照的条件下萌发率在20%以下,而在有光的条件下其萌发率均在40%以上,差异多达到极显著水平(P<0.01)[16,18];而三叶鬼针草、望江南(Cassiaoccidentalis)、马唐(Digitariaadscendens)、毛龙葵(Solanumsarrachoides)等在周期性光照和全黑暗下种子萌发率并无显著差异[14,19-20];麦仁珠(Galiumtricornutum)种子在有光时萌发会受到抑制[21]。本试验研究发现,在35 ℃/30 ℃以下温度时,白花鬼针草在周期性光照和全黑暗2种条件下的种子萌发率并无显著差异,在30 ℃/25 ℃以下温度时,金盏银盘在2种光条件下的种子萌发率也无显著差异,但白花鬼针草在40 ℃/35 ℃、金盏银盘在35 ℃/30 ℃时周期性光照和全黑暗下种子萌发率差异显著,可能是因为当温度成为限制条件时,适当的光照能促进种子的萌发。这跟前人研究的其他物种有一定的差别,这也是其对环境的一种适应策略之一。白花鬼针草种子的宽温度适应范围、快速萌发、高萌发率及萌发的不需光性为其在新环境中种群的建立及扩散并最终形成入侵提供了很好的保障。
[1]田兴山,岳茂峰,冯 莉,等. 外来入侵杂草白花鬼针草的特征特性[J]. 江苏农业科学,2010(5):174-175.
[2]邢福武,曾庆文,谢左章. 广州野生植物[M]. 贵阳:贵州科学技术出版社,2007:224-225.
[3]Linda C D.Bidensalba[EB/OL]. (2009-12-16). http://www.floridata.com/ref/B/bide_alb.cfm.
[4]何报作,曾 静,韦 郃,等. 鬼针草与易淆品白花鬼针草的叶形态——脉序图谱鉴别特征[J]. 中国中药杂志,2009,34(20):2559-2563.
[5]陈志云,梁水凤,李东文,等. 假臭草等12种植物对白花鬼针草幼苗的化感作用[J]. 热带亚热带植物学报,2011,19(5):454-462.
[6]刘明超,韦春强,唐赛春,等. 不同土壤养分水平下2种外来鬼针草和近缘本地种的比较研究[J]. 生物安全学报,2012,21(1):32-40.
[7]王岸英,张玉茹. 菊科8种鬼针草属(BidensL.)杂草种子的鉴别[J]. 吉林农业大学学报,2002,24(3):57-59,64.
[8]Feng Y L,Fu G L. Nitrogen allocation,partitioning and use efficiency in three invasive plant species in comparison with their native congeners[J]. Biological Invasions,2008(10):891-902.
[9]Gerlach J D,Rice K J. Testing life history correlates of invasiveness using congenetic plant species[J]. Ecological Applications,2003,13:167-179.
[10]王 坤,杨 继,陈家宽. 近缘种比较研究在植物入侵生态学中的应用[J]. 生物多样性,2009,17(4):353-361.
[11]Etejere E O. Viability of herbicide-treated seeds ofEupatoriumodoratumL.[J]. Weed Research,1980,20(6):361-363.
[12]全国明,毛丹鹃,章家恩,等. 飞机草的繁殖能力与种子的萌发特性[J]. 生态环境学报,2011,20(1):72-78.
[13]Stanton L E. The estblishment,expansion and ecosystem effects ofPhragmitesaustralis,an invasive species in coastal Louisiana[D]. Louisiana:Louisiana State University,2005.
[14]洪 岚,沈 浩,杨期和,等. 外来入侵植物三叶鬼针草种子萌发与贮藏特性研究[J]. 武汉植物学研究,2004,22(5):433-437.
[15]杨期和,杨威,李秀荣. 热带植物种子萌发影响因素初探[J]. 种子,2001(5):45-48.
[16]韦春强,刘明超,唐赛春,等. 光照和温度对飞机草种子萌发的影响[J]. 广西植物,2012,32(4):527-530.
[17]Fenner M. Seeds:the ecology of regeneration in plant communities[M]. 2nd ed. Oxford:CABI Publishing,2000:259-284.
[18]许慧男,王文杰,于兴洋,等. 菊科几种入侵和非入侵植物种子需光发芽特性差异[J]. 生态学报,2010,30(13):3433-3440.
[19]Norsworthy J K,Oliveira M J. Coffee senna (Cassiaoccidentalis) germination and emergence is affected by environmental factors and seeding depth[J]. Weed Science,2005,53(5):657-662.
[20]Zhou J K,Deckard E L,Ahrens W H. Factors affecting germination of hairy nightshade (Solanumsarrachoides) seeds[J]. Weed Science,2005,53(1):41-45.
[21]Chauhan B S,Gill G,Preston C. Factors affecting seed germination of threehorn bedstraw (Galiumtricornutum) in Australia[J]. Weed Science,2006,54:471-477.
SeedGerminationofBidensalbaL.andB.biternataL.underDifferentLightandTemperatureRegimes
WEI Chun-qiang1,PAN Yu-mei1,TANG Sai-chun1,LIN Chun-hua1,2,LIU Ming-chao1,2
(1.Guangxi Institute of Botany,Guangxi Zhuang Autonomous Region and Chinese Academy of Sciences,Guilin541006,China; 2.Guangxi Normal University,Guilin 541004,China)
BidensalbaL. is an invasive alien weed,andB.biternataL. is its congener. The effects of different light and temperature conditions on seed germination ofB.albaandB.biternatawere studied by simulating environmental conditions in climatic chambers.B.albaadaptive temperature range for seed germination was wider than that ofB.biternata.B.albawas able to germinate at temperatures from 15 ℃/10 ℃ to 40 ℃/35 ℃,with its germination percentage being about 94% under temperatures from 15 ℃/10 ℃ to 30 ℃/25 ℃.B.biternataseed did not germinate at 40 ℃/35 ℃. Temperature could affect the time of germination peak and height of the seedlings of both species but light had no differential effect.
BidensalbaL.;BidensbiternataL.; seed germination; light; temperature
Q945.34
A
1003-935X(2013)04-0001-04-
韦春强,潘玉梅,唐赛春,等. 不同光照和温度下白花鬼针草和金盏银盘种子萌发研究[J]. 杂草科学,2013,31(4):1-4.
2013-09-06
国家自然科学基金(编号:31260154);广西壮族自治区自然科学基金(编号:2011GXNSFE018005)。
韦春强(1982—),男,壮族,广西宾阳人,硕士,从事外来入侵植物研究。E-mail:weichun007@163.com。
唐赛春。E-mail:tangs@gxib.cn。
