山西省瓜类作物根茎部镰刀菌的分离与鉴定
2013-10-22袁微微李新凤高俊明王建明郝晓娟贺运春
袁微微,李新凤,高俊明,王建明,郝晓娟,贺运春
(山西农业大学农学院,山西太谷030801)
镰刀菌(Fusarium)是一类具有重要经济意义的真菌,广泛分布于自然界中,兼性寄生或腐生生活。其中,多种镰刀菌是重要的植物病原菌,由镰孢菌侵染引起的瓜类枯萎病、瓜类根腐病等瓜类作物萎蔫病,都是经土壤传播的系统性病害,且均为生产上最难防治的病害。通常情况下,其在黄瓜、西瓜、冬瓜上发病最为严重,一般年份发病率为20%,严重的田块可达80%~90%,是目前生产上制约瓜类作物生产的重要因素之一[1-2]。此外,一些弱致病镰刀菌和非致病镰刀菌还是植物的主要内生菌,不仅能使寄主植物快速生长,增强植物的抗逆性,而且能产生丰富多样的活性物质,从而对病原细菌、真菌和线虫产生拮抗作用,具有生物防治作用[3-10]。可见,镰孢菌属真菌对农业生产作用重大。明确一个地区某种作物上镰刀菌的种类,对于植物镰刀菌病害的检疫、防治和筛选拮抗作物病原菌的镰刀菌都具有重要意义。
笔者于2010年4月至2011年8月在山西省各地区采集了具有萎蔫症状与健康的瓜类作物标本共93份,进行了镰刀菌的分离、纯化,并依据其形态特点与培养性状进行了初步鉴定,以期了解山西省瓜类作物上镰刀菌的种类及分布特点,并有效防治其菌病害,提高瓜类作物产量及进一步筛选具有生防作用的镰刀菌资源提供理论依据。
1 材料与方法
1.1 试验材料
1.1.1 Armstrong液体培养基 葡萄糖20.0 g,MgSO4·7H2O 0.4 g,KCl 1.6 g,KH2PO41.1 g,Ca (NO3)25.9 g,FeSO4200 μg,MnSO4200 μg,ZnSO4200 μg,去离子水 1 000 mL。
1.1.2 康乃馨叶片培养基(CLA)[11]琼脂20 g,水1 L,康乃馨叶片适量。将3~5 mm2大小的无菌康乃馨叶片放入冷却到45℃的灭菌水琼脂平板表面(一般情况下,直径为9 cm的培养皿内加入10~12块即可)。接种时,菌块要靠近康乃馨叶片。
1.1.3 Spezieller Nährstoffarmer琼脂培养基(SNA)
KCl 0.5 g,MgSO4·7H2O 0.5 g,KNO31 g,KH2PO41 g,蔗糖 0.2 g,葡萄糖 0.2 g,琼脂 20 g,蒸馏水1 L。将灭菌培养基倒入培养皿中制成平板,待平板冷却后,在其表面放入1~2片滤纸条[12]。
1.1.4 2%水琼脂培养基(WA) 琼脂20 g,自来水1 L。
1.2 试验方法
1.2.1 样本采集 样本采集的时间和地点如表1所示。采集的样本种类主要为黄瓜、西瓜、西葫芦等瓜类作物的根茎组织。

表1 供试菌株及来源
1.2.2 镰刀菌的分离与纯化 采用常规组织分离法,对采集到的样本的根茎部镰刀菌进行分离、纯化。将供试材料用无菌刀片切成小块(0.5 cm2),置于70%酒精中浸泡几秒钟,取出后用无菌水冲洗干净,并用灭菌滤纸吸干表面的水分,然后置于2%的水琼脂平板表面,每平板4~5块,之后置于25℃的恒温培养箱内进行培养。待长出菌落后,用稀释平板法进行单孢分离。然后利用形态学分类方法对单孢分离物进行初步鉴定、编号,并记录不同种镰刀菌的分离频率。培养过程中,为避免细菌污染,可在培养基中加入适量抗菌素(如氯霉素、链霉素等),一般在培养基灭菌后冷却至45℃左右时加入。
挑取菌体置于显微镜下镜检,将明显混杂其他杂菌的镰刀菌菌落要先接入平板,经数次纯化后进行单孢分离;如果经镜检发现菌落没有污染其他杂菌,则可直接进行单孢分离。
1.2.3 镰刀菌的培养及形态观察 参考文献[13]的方法,将单孢分离后的培养物接种到CLA,SNA,PDA,WA培养基上适当培养,进行培养性状及显微形态观察,用于该镰孢菌种的鉴定。培养性状主要包括菌落在PDA培养基上的质地、颜色以及基质颜色和菌体生长速率。显微形态包括大、小型分生孢子及厚垣孢子、子实体的形态,产孢细胞、产孢梗的形态及形成方式等。
1.2.4 镰刀菌的鉴定 参考Leslie等[12,14-15]的分类系统,对分离到的镰刀菌菌株进行形态学鉴定。
2 结果与分析
2.1 样品分离、纯化结果
从山西省采集的93份瓜类作物样品中,共分离到353株镰刀菌,经形态学鉴定,其分属于8种镰刀菌(表2),分别是尖孢镰刀菌(F.oxysporum)、芬芳镰刀菌(F.redolens)、腐皮镰刀菌(F.solani)、轮枝镰刀菌(F.verticillioides)、层生镰刀菌(F.proliferatum)、禾谷镰刀菌(F.graminearum)半裸镰刀菌(F.semitectum)、锐顶镰刀菌(F.acuminatum)。其中,尖孢镰刀菌225株,占镰刀菌总量的65.03%,为优势菌群;其次是腐皮镰刀菌,共分离到60株,占总量的17.34%;芬芳镰刀菌和锐顶镰刀菌较少,分别只有2,3株。
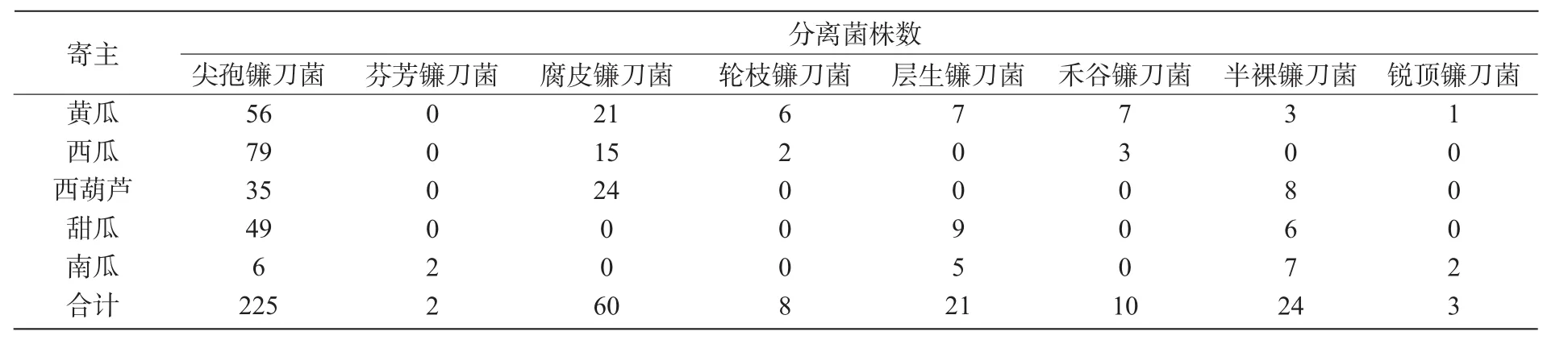
表2 镰刀菌分离结果
2.2 镰刀菌形态特点
2.2.1 尖孢镰刀菌 由图1可知,菌落表面常见菌丝扭结成束后凌乱地编织成类似网状的结构,菌落生长速度较快;PDA上25℃培养4 d,菌落直径为4.9~5.2 cm;气生菌丝羊毛状,白色、淡青莲至淡粉色;基物表面茄皮紫,基物不变色。由气生菌丝上侧生产孢细胞,在有稀疏分枝或无分枝的分生孢子梗上形成单瓶梗,产孢细胞安瓿瓶形、较短、领口明显。产生大量假头生的小型分生孢子,椭圆形、卵形或肾形,0~1个分隔,量度为(4.8~10.1)μm×(2.0~4.6)μm。易产生桔黄色黏孢团型分生孢子座。大型分生孢子美丽型,孢子较细、较弯曲,向两端比较均匀地逐渐变尖,足跟明显。细胞壁薄,2~6个分隔,多为3~5个分隔,量度为(20.2~36.6)μm×(3.3~4.5)μm。厚垣孢子球形,表面光滑,单生、双生或聚生于菌丝间。

2.2.2 芬芳镰刀菌 由图2可知,菌落蛛网状,PDA上25℃培养4 d,菌落直径为4.1~4.6 cm。

其气生菌丝羊毛状或毡状,苍白、米色至淡青莲色;基物表面青莲色,基物不变色。自气生菌丝上侧生或在短的分枝上形成瓶状小梗,单瓶梗,产孢梗长度介于尖孢镰刀菌与腐皮镰刀菌之间,分枝少。小型分生孢子数量多,假头生,椭圆形,卵形或肾形,0~1个分隔,量度为(6.72~18.34)μm×(2.30~4.13)μm。大型分生孢子介于美丽型与马特型之间,比尖孢镰刀菌稍宽,顶胞稍钝,基胞足跟不明显,分隔不清楚,量度为(31.2~46.2)μm×(3.7~6.3)μm。厚垣孢子球形,表面光滑,单个顶生或串生。
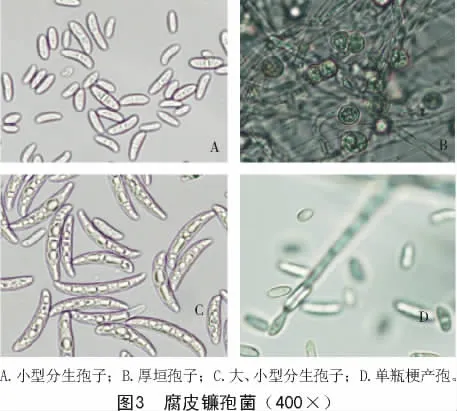
2.2.3 腐皮镰刀菌 由图3可知,PDA上25℃培养4 d,菌落直径为2.8~4.9 cm。气生菌丝羊绒状,白色至浅灰色,大多数菌株呈奶油色,中央有土黄色黏孢团,少数菌株菌落背面可产生蓝紫色色素;基物表面肉色,基物不变色。前期在气生菌丝上长出的产孢细胞为菌丝形的单瓶梗,产孢梗长,大于 50 μm,长可达到 200 μm,在成熟培养物上形成分生孢子座,其上可成簇产生短而多分枝的产孢梗,顶端领口明显。大型分生孢子马特型,即大型分生孢子最宽处在孢子中线上部,两端较钝,顶细胞稍弯;基细胞钝圆形或足跟不明显,整个孢子形态较短、较胖,2~7个分隔,多数 3~5个分隔,量度为(23.7~37.7)μm×(3.7~6.4)μm。有大孢子融合现象。在幼培养中会产生大量假头生小型分生孢子,椭圆形、卵形或肾形,比较宽,0~1个分隔,多为1个分隔,量度为(7.6~17.7)μm×(2.1~5.4)μm,明显比其他种类镰刀菌的小型分生孢子粗胖。
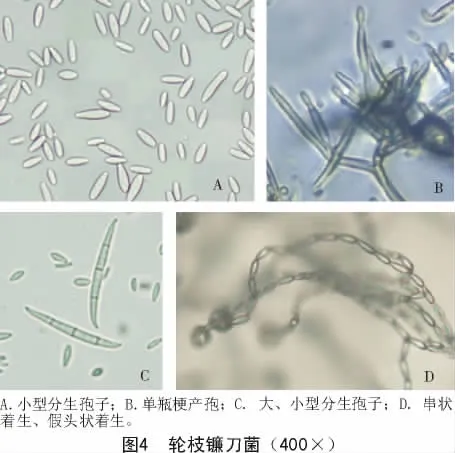
2.2.4 轮枝镰刀菌 由图4可知,气生菌丝羊毛状至粉状,粉色至紫色,后产生深紫色色素,基物表面及基物为暗紫色。在PDA平板上25℃培养4 d后,菌落直径为35~46 mm。产孢细胞单瓶梗,瓶形,分枝或不分枝。小型分生孢子多,呈链状,分生孢子链较长,一般都大于50 μm,有时聚集形成假头状。小孢子一端平截,为典型的棒槌形,没有或有1个分隔,量度为(4.1~8.2)μm×(1.7~3.9)μm。菌体培养后期可产生桔黄色分生孢子座。大型分生孢子细长、较直,孢子的背腹两侧近似平行,两端微收缩。顶细胞为渐细的锥形,略弯曲,基细胞为不明显的足跟状,大孢子壁较薄,分隔明显,一般为3~5个分隔,量度为(24.3~41.4)μm×(3.7~4.9)μm。
2.2.5 层生镰刀菌 由图5可知,菌落蛛网状,生长速度中等,PDA上25℃培养4 d后,菌落直径为4.7~4.9 cm。气生菌丝羊毛状,菌落初期白色或淡紫色,基物紫色。气生菌丝上有分枝较多的产孢梗,单瓶梗或复瓶梗(层出梗),产孢细胞瓶形。常见较长的产孢细胞旁边有一个很短的产孢点,或形成Z字形,层生的复瓶梗,为典型的层出复瓶梗。小型分生孢子串生或假头生,卵形、椭圆形或棒槌形,通常无分隔,量度为(6.9~19.9)μm×(2.3~4.7)μm。在气生菌丝的产孢细胞上形成大量较长的分生孢子链。有些菌株培养后期可产生桔黄色分生孢子座。大型分生孢子镰刀形,较直而细长,细胞壁薄,且通常不易产生,顶细胞尖而弯曲,足细胞明显,3~5个分隔,量度为(27.3~49.2)μm×(3.3~4.6)μm。缺乏厚垣孢子。

2.2.6 禾谷镰刀菌 由图6可知,在PDA培养基上生长迅速,菌落生长初期呈放射状。气生菌丝呈羊毛状旺盛生长,白色至紫红色,菌落中央常呈浅驼色至黄褐色,菌落背面为紫红色。在25℃下培养4 d后,菌落直径为2.6~3.3 cm。产孢细胞单瓶梗,小型分生孢子缺乏。在SNA或CLA上产生桔黄色分生孢子座。大型分生孢子较直或镰刀形,顶细胞略弯曲、渐尖,短喙状,基部足跟明显,一般3~6个分隔,多为5个分隔,量度为(39.13~78.73)μm×(3.99~6.37)μm。在培养后期大孢子数量明显减少。厚垣孢子聚生或串生,间生或顶生,球形或近球形,浅褐色。

2.2.7 半裸镰刀菌 由图7可知,在PDA培养基上25℃培养4 d后,菌落直径为4.2~5.6 cm。气生菌丝羊毛状或羊绒状,初期白色至淡粉红,菌落背面粉色,后期菌落变为浅驼色至褐色,基物表面茶褐色,菌落背面深褐色。产孢细胞单瓶梗、复瓶梗和多芽产孢共存。气生菌丝上产生的大、小孢子间没有明显界限,存在大量的中型分生孢子。分生孢子多数为倒卵形或纺锤形,稍弯或直。分生孢子座上产生的孢子镰状、稍弯,顶细胞楔形,基细胞突起,足跟有或无,多为3~5个分隔,量度为(28.72~39.99)μm×(4.61~5.93)μm。厚垣孢子极少,球形,单顶生或串生于菌丝中间,褐色。

2.2.8 锐顶镰刀菌 由图8可知,在PDA上培养基上25℃培养4d后,菌落直径为4.4~5.0cm。气生菌丝棉絮状,淡粉红至淡豆沙色,有橙色黏孢团。基物表面茶褐色,基物不变色。通常为单瓶梗产孢,小型分生孢子少,锥形或镰状。橙色黏孢团上上易形成分生孢子座。大孢子整齐典型,镰刀形,腹背明显弯曲,顶胞呈锐角、细长。基胞足跟明显,有2~4个分隔,多为3个分隔,量度为(19.7~36.6)μm×(2.7~4.7)μm。厚垣孢子球形,表面不光滑,单顶生或串生于菌丝。

3 结论与讨论
通过对山西省黄瓜、西瓜、西葫芦等瓜类作物上镰刀菌的分离、纯化、鉴定,初步明确了山西省瓜类作物根茎部镰刀菌有8种,分别为尖孢镰刀菌(F.oxysporum)、芬芳镰刀菌(F.redolens)、腐皮镰刀菌(F.solani)、轮枝镰刀菌(F.verticillioides)、层生镰刀菌(F.proliferatum)、禾谷镰刀菌(F.graminearum)半裸镰刀菌(F.semitectum)、锐顶镰刀菌(F.acuminatum)。其中,尖孢镰刀菌为优势菌群,其次是腐皮镰刀菌。这一结果为进一步研究山西省瓜类作物镰刀菌病害、镰刀菌的生物防治等方面奠定了理论基础。
寄生于植物体内的镰刀菌,多数为植物病原菌,如瓜类枯萎病是由寄生于瓜类作物的尖孢镰刀菌引起,腐皮镰刀菌是引起瓜类作物根腐病的主要病原菌。本研究所选材料有些为健康植株,而有些则是具有萎蔫症状的病株,鉴定结果表明,尖孢镰刀菌为优势菌群,腐皮镰孢菌也较其他菌种多,所以可以确定本试验分离到的这2种镰刀菌中部分菌株为致病菌。此外,在枯萎病和根腐病的病株中,还分离到与报道的病原菌不同的其他菌株。如西葫芦根腐病病株中除分离到腐皮镰刀菌外,还分离到半裸镰刀菌;西瓜枯萎病病株中除分离到尖孢镰刀菌还分离到禾谷镰刀菌,但半裸镰刀菌和禾谷镰刀菌对于这2种病害的作用及其与寄主植物的关系还有待进一步研究。
植物内生真菌与寄主植物关系复杂,研究表明,许多内生镰刀菌中的非致病性菌株和弱毒菌株对作物的微管束病害具有交叉保护和诱导抗性作用。据文献报道,有些可用来防治甘蓝、番茄、黄瓜、西瓜、甘薯等作物的枯萎病[5,7-8],有些则用来控制植物寄生线虫[16-17]。本研究中,部分菌株分离自枯萎病或根腐病严重发生地块的健康植株,这为进一步从这些菌株中开发对枯萎病或根腐病等根部病害具有生防作用的菌株奠定了基础。
[1]韩金星,洪日新,周林,等.西瓜、黄瓜、甜瓜等瓜类枯萎病研究进展[J].中国瓜菜,2009(2):32-35.
[2]王萍,肖艳,曾敬富,等.瓜类枯萎病的识别与诊断及综合防控技术初报[J].现代园艺,2010(2):46-47.
[3]王拱辰,叶琪铭.镰刀菌在生物防治中的作用[J].生物防治通报,1990,6(2):80-84.
[4]杨涛,张贵锋,关天舒,等.黄瓜、甜菜专化型尖孢镰刀菌人工诱变非致病菌系筛选及其生物防治应用研究 [J].辽宁农业科学,2001(4):11-13.
[5]关营昌,王守正.利用弱致病菌株诱导黄瓜抗枯萎病研究[J].河南农业大学学报,1991,25(4):433-437.
[6]Larkin R P.Suppression of Fusarium wilt of watermelon Fusarium oxysporum and other microorganisms recorvered from a disease suppressive soil[J].Phypathology,1996,86:812-819.
[7]Mandeel Q,Bank er R.Mechanisms involved in biological control of Fusarium wilt of cucumber with strains of nonpathogenic Fusarium oxysporum[J].Phytopathology,1991,81:462-469.
[8]Park C S,Paulitz T C,Barker R.Biocontrol of Fusarium wilt of cucumber resulting from interactions between Pseudomonas putida and nonpat hogenic isolates of Fusarium oxysporum[J].Phytopathology,1998,78:190-194.
[9]Dababat A A,Sikora R A.Induced resistance by the mutualistic endophyte,Fusariumoxysporum 162,towardMeloidogyne incog-nita on tomato[J].Biocontrol Science and Technology,2007,17(6):969-975.
[10]Vu T,Hauschild R,Sikora R A.Fusarium oxysporum endophytes induced systemic resistance against Radopholus similis on banana[J].Nematology,2006,8(6):847-852.
[11]Fisher N L,Burgess L W,Toussoun T A,et al.Carnation leaves as a substrate and for preserving cultures of Fusarium species[J].Phytopathology,1982,72:151-153.
[12]Leslie J F,Summerell BA.The Fusarium laboratorymanual[M].USA:Blackwell Publishing,2006.
[13]李新凤.山西省玉米穗腐病病原镰孢菌的分离与鉴定[J].山西农业大学学报:自然科学版,2012,32(3):218-223.
[14]Nelson P E,Toussoun T A,Marasas W F O.Fusarium species:Anillustrated manual for identification[M].London:The Pennsylvania State UniversityPark,1983.
[15]Booth C.The genus Fusarium[M].London:Commonwealth Mycological Institute,1971.
[16]Hallmann J,Sikora R A.Influence of Fusarium oxysporum,a mutualistic fungal endophyte,on Meloidogyne incognita infection of tomato[J].Zeitschrift fur Pflanzenkrankheiten und Pflanzenschutz,1994,101(5):475-481.
[17]Le H T T,Padgham J L,Sikora R A.Biological control of the rice root-knot nematode Meloidogyne graminicola on rice,using endophytic and rhizosphere fungi[J].International Journal ofPest Management,2009,55(1):31-36.
