亚洲小车蝗在不同生境中的群落动态研究
2013-09-28张未仲吴惠惠刘朝阳曹广春张泽华格西格
张未仲, 吴惠惠, 刘朝阳, 曹广春, 张泽华*, 格西格
(1.农业部作物有害生物综合治理重点实验室,北京 100081;2.内蒙古锡林郭勒盟镶黄旗草原工作站,镶黄旗 013250)
亚洲小车蝗(OedaleusdecorusasiaticusB.Bienko)隶属于直翅目,蝗总科,斑翅蝗科,是北方草原的主要危害种,主要取食禾本科植物,是内蒙古典型草原优势蝗虫种之一[1-2],而且亚洲小车蝗常选择放牧强度高且较为干旱的生境,是内蒙古草原草场退化的指示种[3]。
食性食量分析是研究蝗虫营养生态位的基础。营养生态位是指动物对其食物资源能够实际和潜在占据、利用或适应的部分[4]。通过对蝗虫的食性食量分析可以了解其取食特性,以及对植物资源的利用情况,进而分析草原蝗虫的营养生态位,从而可以了解蝗虫群落的结构以及多样性和稳定性,并且能够探讨环境因素的变化对蝗虫群落生态关系的影响,特别是草原植物群落结构的组成变化与蝗虫群落结构及数量之间的关系,这对于完善草原管理,指导蝗虫综合治理也有着重要的现实意义[5]。从20世纪60年代开始,已经用野外罩笼、模拟取食,嗉囊分析等方法对亚洲小车蝗食性食量进行了分析,确定了小车蝗食谱主要为禾本科植物,以及自然条件下个体寿命及食量[6-11]。近年来研究了亚洲小车蝗取食对植物补偿生长的影响及损失估计,并提出了防治阈值[12-13]。以前的研究,通过野外罩笼,来研究食性食量,可以近似模拟蝗虫野外自然条件下的取食动态,但是由于地理风貌,植被分布,以及气候(温度、湿度)等多种因素的影响,蝗虫在自然条件下的取食受外界影响较大[14],不利于控制试验条件,进行单因素分析。而室内观测结果与野外的实际草场观测相比,虽然有一定的差异,但可以方便地控制试验条件,有利于定量分析,并且对野外的观测数据有一定的修正作用。野外试验同室内观测相结合的研究方法尚未见报道。
本文希望通过在蝗虫发生时,采用剪样方的方法确定自然条件下不同草场的植被变化,用扫网及抛样框方法分析蝗虫种群动态,并结合室内亚洲小车蝗的食量分析,来反映亚洲小车蝗在不同生境中的取食情况。为野外防治提供理论依据。
1 材料与方法
1.1 样地选择
试验样地选在内蒙古锡林郭勒盟镶黄旗。经度113°83′,纬度42°25′,该地降水偏少且分布不均,在牧草生长的关键季节(5-8月份)的有效降雨量不足150 mm,全旗可利用的地表水基本没有。春季常有沙尘暴,冬季常有旱灾、雪灾发生。地区年日照时数3 031.6 h,平均日照百分率68%。地区年平均气温3.1℃,月平均气温从7月份20.4℃到1月份-16.5℃。无霜期较短,平均为120 d左右,最大冻土深度154 cm。
2010、2011年在亚洲小车蝗的发生季节,选择3块较为平坦但优势种不同的样地。1号地为围栏草地,以羊草为优势种,盖度约为45%,2号地和3号地为放牧草场,以针茅为优势种,退化较严重,盖度<35%。样地主要分布有克氏针茅(StipakryloviiRoshev.)羊草[Leymuschinensis(Trin.)Tzvel.],糙隐子草[Cleistogenessquarrosa(Trin.)Keng]、旋覆花(ConvolvulusammanniiDesr.)小叶锦鸡儿(CaraganamicrophyllaLam.)、冷 蒿 (Artemisia frigidaWilld.)等植物。
1.2 样地调查方法
在每块样地随机选取1m2样方,数码相机垂直摄像,用实验室软件分析样方的总盖度。记录样方内所有植物种类、每种植物的盖度、高度、密度。将样方内所有的植物地表部分分类剪下,分别保存于信封内,在室内烘干,称量植物干重,记录每种植物的生物量。每块样地5次重复。每隔15 d采样一次。
1.3 蝗虫调查方法
2010年用扫网的方法采集蝗虫样本。200次1个处理,重复10次。2011年用1 m×1 m×1 m无底样框采集蝗虫样本,重复10次。3块样地每隔10 d采样一次。先将网中的蝗虫放入无水乙醇中保存,回到室内记录蝗虫种类、数量、龄期。
1.4 亚洲小车蝗室内饲养方法
在野外采集羊草、针茅,在室内擦洗干净,取新鲜叶片组织,称取1~1.5 g,底端用棉花包裹,浸水,塞入2 m L离心管,放入塑料盒中。挑选相同龄期的小车蝗,雌、雄分别饲养。一个盒中放入3头,并在盒盖扎孔通气,以没有放虫的盒作为对照。重复5次。每隔24 h换虫,称量剩余的草重量,计算蝗虫的取食量。从3龄一直做到成虫。
1.5 数据分析方法
1.5.1 生物多样性比较
用两种常见的生物多样性指数分析植物和蝗虫多样性[5,24]。
辛普森多样性指数(Simpson′s diversity index)

香农-威纳指数(Shannon-Weiner index)
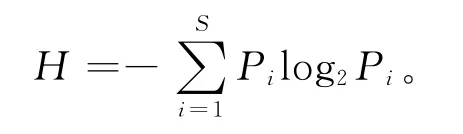
上述公式中,设种i的个体数占群落中总个体数的比例为P i,S为物种数目,并且分析植物与蝗虫多样性之间,以及植物生物量与蝗虫数量的相关性。
1.5.2 室内蝗虫取食量计算方法
取食量=对照植物生物量-取食后生物量。
1.5.3 数据处理分析
对蝗虫数量与植物盖度、植物生物量之间作相关及回归分析;蝗虫多样性与植物多样性之间作相关分析;蝗虫食量进行单因素方差分析。数据用统计软件SAS 8.0完成[10]。
2 结果与分析
2.1 三块样地植物与蝗虫概况
如表1、表2所示,在2010年和2011年两年的6-8月间,选取3块样地调查了植物盖度与生物量,蝗虫数量以及植物与蝗虫的多样性。其中,1号地草场的盖度、平均生物、蝗虫数量和亚洲小车蝗数量在蝗虫生长季明显区别于其他两块样地。总体上,2011年样地植物生物量均高于2010年。

表1 2010年室外试验调查结果1)Table 1 The data of outdoor experiments in 2010
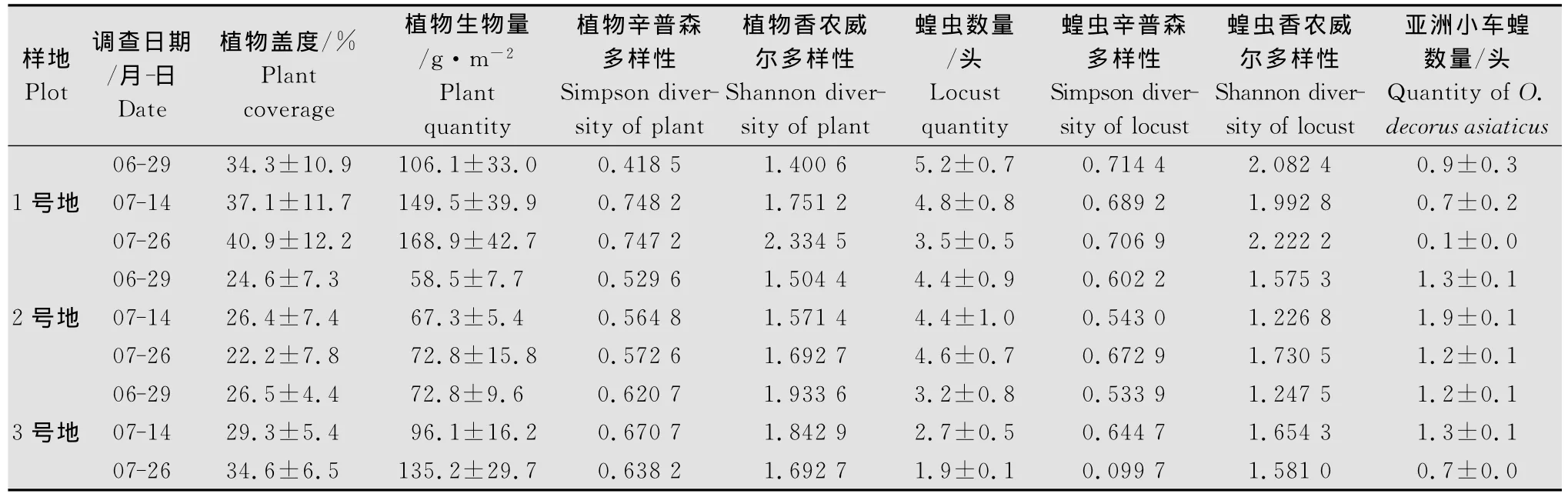
表2 2011年室外试验调查结果1)Table 2 The data of outdoor experiments in 2011
2.2 植物盖度与蝗虫数量的关系
为了研究亚洲小车蝗数量与植物盖度之间的关系,将每次调查的样地植物盖度同亚洲小车蝗数量做相关性分析。3块样地的植物盖度同亚洲小车蝗数量呈负相关(Y=233 42X-2.6052,R=-0.770 8,F=120.45,P<0.01,图1)。但是,当植物盖度大于25%时,亚洲小车蝗数量趋于稳定,当盖度小于15%时,蝗虫数量急剧增加。为了研究植物盖度同蝗虫总体数量之间的关系,将每次调查的样地植物盖度同蝗虫数量做相关性分析。3块样地的植物盖度同蝗虫总数呈负相关,蝗虫的数量随着植物盖度的增大而减 小(Y=666.77X-1.1136,R= -0.759 3,F=58.59,P<0.01,图2)。但是,当植物盖度大于25%时,蝗虫总数随盖度增加而减少且趋于稳定,当盖度小于15%时,蝗虫数量随着盖度的降低而迅速增加。

2.3 植物生物量与蝗虫数量的关系
3块样地的植物生物总量同亚洲小车蝗数量呈负相关,蝗虫的数量随着植物生物量的增大而减小(Y=355 41X-1.8893、R=-0.905 2、F=78.11、P<0.01、图3)。当植物生物量大于时70 g/m2时,亚洲小车蝗数量趋于稳定;当植物生物量小于70 g/m2时,小车蝗数量随生物量减小逐渐增大;当植物生物量小于30 g/m2时,随着生物量的减小亚洲小车蝗数量急剧增大。

图3 亚洲小车蝗数量同植物平均生物量关系Fig.3 The relationships between the average amount of O.decorus asiaticus and plant biomass
3块样地的植物生物总量同蝗虫总数呈负相关,蝗虫的数量随着植物生物量的增大而减小(Y=1 497.5X-0.87,R=-0.859 7,F=59.83,P<0.01,图4)。当植物生物量大于70 g/m2时,蝗虫数量趋于稳定;当植物生物量小于70 g/m2时,蝗虫数量随生物量减小逐渐增大;当植物生物量小于30 g/m2时,随着生物量的减小蝗虫数量急剧增大。
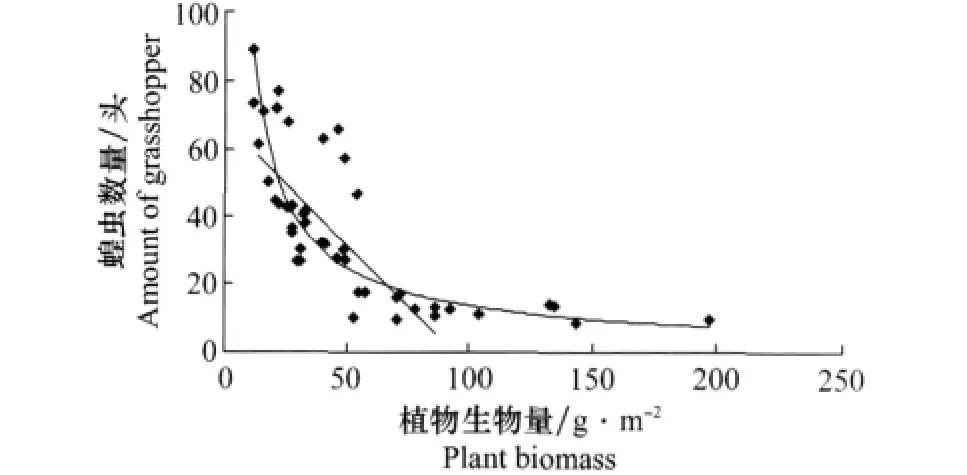
图4 蝗虫总数与植物平均生物量关系Fig.4 The relationships between the average amount of grasshopper and plant biomass
2.4 植物多样性与蝗虫多样性的相关性
相关分析可知,植物同蝗虫之间辛普森多样性呈正相关Y=0.702 6X+0.094 7(R=0.588 4,F=3.78,P<0.05,图5);植物同蝗虫的香农-威尔多样性指数呈正相关Y=0.579 4X+0.315 8(R=0.521 5,F=5.98,P<0.05,图6)。

2.5 亚洲小车蝗室内植物消耗量分析
从表3可知,亚洲小车蝗雌虫对针茅、羊草的消耗量都随时间推移而逐渐增大,且羊草消耗量高于针茅。两种植物的消耗量在7月14日达到一个高峰然后稍有下降,针茅消耗量在7月19日回升后有所下降,羊草在7月20日达到最大值后有所下降。7月14日前都是蝗蝻,14日以后都是成虫。7月1日3龄蝗蝻植物消耗量最低,6日、7日是4龄蝗蝻,8日、11日是5龄蝗蝻,4龄和5龄蝗蝻间、14日之前,6日、11日羊草消耗量明显高于针茅(P<0.05),14日以后,除19日外,每日雌虫对羊草消耗量均明显高于针茅(P<0.05),但是同种植物之间没有显著差异(P>0.05)。
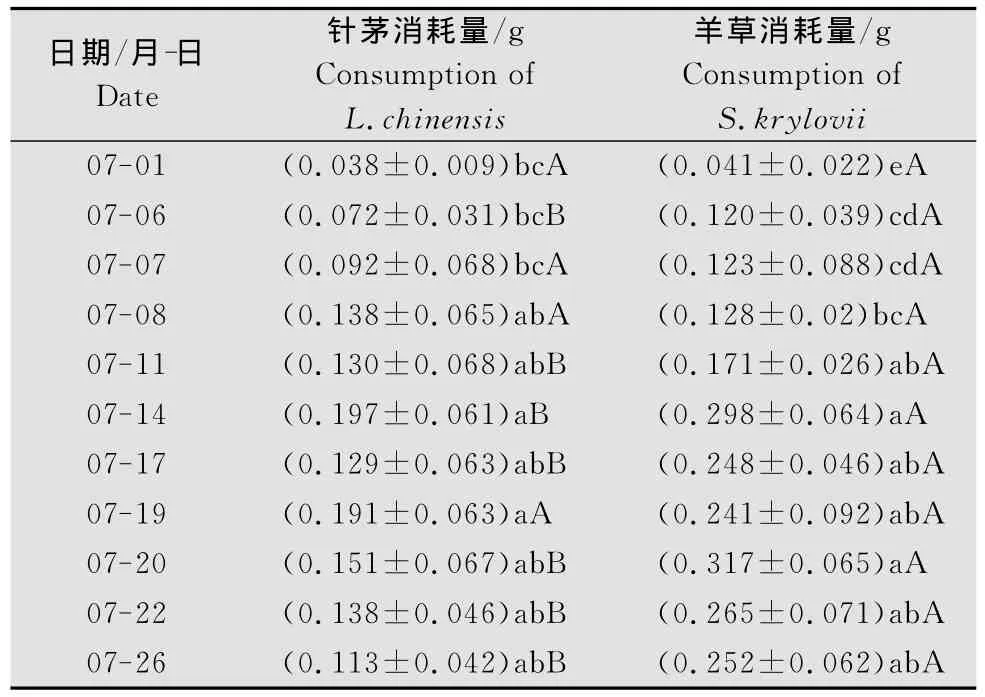
表3 亚洲小车蝗雌虫每日植物消耗量1)Table 3 Daily consumption of L.chinensis and S.krylovii by female O.decorus asiaticus
从表4可以看出亚洲小车蝗雄虫对针茅、羊草的消耗量都有一个缓慢增高然后逐渐降低的趋势,并且对羊草的消耗量大于针茅。两种植物消耗量在7月14日达到最大值,在17日、19日间有一个波动,然后逐渐降低,在蝗蝻期只有6日、11日对两种植物消耗量有显著差异(P<0.05);在成虫期21日后两种植物间消耗量差异显著(P<0.05),但是同种植物之间没有显著差异(P>0.05)。
3 结论与讨论
由于全球性气候变化和人类对草原自然资源过度开发利用,导致了草地生态环境逐渐恶化,蝗灾频繁发生且为害程度不断加剧[15]。由于草地面积锐减,草场沙漠化和盐碱化日益加重,造成草地生物量急剧减少及生物多样性的破坏,严重影响了草原畜牧业可持续发展和牧区的经济建设。研究蝗虫、草地生物量及生物多样性之间的关系,对于明确危害程度,提供防治策略,改善生态环境,保持草原牧区畜牧业的持续发展都有重要意义。

表4 亚洲小车蝗雄虫每日植物消耗量1)Table 4 Daily consumption of L.chinensis and S.krylovii by male O.decorus asiaticus
蝗虫的分布和种群结构同植物群落类型和结构密切相关[16-17]。本试验研究表明植物多样性同蝗虫多样性呈正相关。两种多样性指数中,植物与蝗虫之间辛普森多样性指数相关性略高于香农-威尔多样性指数相关性。1号地是围栏草地,草地植物均匀度好,多样性高,因此蝗虫种类最多。2号地、3号地是放牧草地,植被退化较为严重,植物物种均匀度较差,因此蝗虫种类较少。通过剪样方与扫网调查共同分析表明植物生物量、盖度与亚洲小车蝗数量以及蝗虫整体数量都呈负相关,且相关性均极显著。1号样地以羊草为优势种,生物量最大,蝗虫数量却最少。2号、3号两块样地以针茅为优势种,针茅生物量明显低于1号样地的羊草生物量,但是蝗虫数量却明显高于1号地。蝗虫与植物生物量的相关性大于蝗虫同盖度的相关性。由于亚洲小车蝗为优势种,植物生物量同亚洲小车蝗数量的相关性高于植物生物量同蝗虫整体数量相关性。野外试验在3块样地中,当盖度小于15%,或植物生物量小于70 g/m2时,蝗虫数量急剧增大。
蝗虫生态位研究,国内外已经做了大量的工作[18-22]。李鸿昌、卢辉等已经用野外罩笼试验研究表明,亚洲小车蝗喜食羊草,少食针茅,偶食冷蒿[1,23]。本试验在室内用两种植物饲养,结果显示亚洲小车蝗的取食量羊草大于针茅,同前人的研究相符。另外室内用冷蒿喂养蝗虫,取食量为0,而且发生自残现象,表明亚洲小车蝗不食冷蒿。同以前的研究相比,亚洲小车蝗对冷蒿的不喜程度更为明显。关敬群等对亚洲小车蝗雌雄间食量差异已作过研究[12],但是他们选用盆栽谷苗作为食料,在研究蝗虫对草场的危害时,没有明显的针对性。本试验用两种草原植物作为食物,研究表明,亚洲小车蝗对植物的消耗量随虫龄的增加而增大,在刚进入成虫期时达到最大值而后略有下降。在试验过程中,雌雄虫在成虫期对羊草的消耗量都高于针茅,而同种植物间植物的消耗量在高龄蝗蝻和成虫期没有明显差异。
室内外试验综合分析表明亚洲小车蝗喜食羊草,少食针茅,即在喜食的植物占优势的植物群落里,如果生境不适合,则蝗虫的数量依旧不高。而在非喜食植物占优势的草地草场,只要环境适宜,蝗虫的数量仍然很高。这表明蝗虫并不是根据对食料的喜好来选择栖息地,而是根据生境的适应与否来选择,环境因子对蝗虫群落的影响大于植物群落对蝗虫群落的影响。这个结论同贺达汉、康乐的研究结果,即蝗虫优先选择合适的栖境相吻合[10,24]。再结合生物多样性分析结果,1号地是围栏草地,2号地、3号地是放牧草地,围栏草地生物多样性高于放牧草地,放牧的重要地区是干旱和半干旱地区,同时也是蝗虫的多发区。过度放牧破坏了草地植被,同时诱发了蝗虫的发生,蝗虫的发生则加剧了草地的破坏,造成了恶性循环。本试验研究表明,过度放牧行为造成草地退化,易形成蝗虫合适的栖境,从而造成了蝗灾的发生。在生产中应实行合理放牧,提高植物覆盖度,降低蝗虫种群数量,达到防治蝗虫的目的。
[1]李鸿昌,席瑞华,陈永林.内蒙古典型草原蝗虫食性研究Ⅰ罩笼供食下的取食特性[J].生态学报,1983,3(3):214-228.
[2]李鸿昌,陈永林.内蒙古典型草原蝗虫食性的研究Ⅱ在自然植物群落内的取食特性[M]∥草原生态系统研究.北京:科学出版社,1985:154-165.
[3]Kang L,Chen Y L.Dynamics of grasshopper communities under different grazing intensities in Inner Mongolian steppes[J].Entomologia Sinica,1995(2):265-281.
[4]康乐,陈永林.草原蝗虫营养生态位的研究[J].昆虫学报,1994,37(2):178-189.
[5]贺达汉,郑哲民.环境因子对蝗虫群落生态效应的数值分析[J].草地学报,1996,4(3):213-220.
[6]钦俊德,郭郛,郑竺英.东亚飞蝗的食性和食物利用以及不同食料植物对其生长和生殖的影响[J].昆虫学报,1957,7(2):143-166.
[7]关敬群,魏增柱.亚洲小车蝗食量测定[J].昆虫知识,1989,26(1):8-11.
[8]冯光翰,樊树喜,刘秋芳,等.室外罩笼条件下几种草原蝗虫的食量测定[J].草地学报,1995,3(3):230-235.
[9]杨群芳,廖志昌,李庆,等.西藏飞蝗食性及防治指标[J].植物保护学报,2008,35(5):399-404.
[10]康乐,陈永林.草原蝗虫食料植物叶片表皮显微结构的研究[M]∥草原生态系统研究,第4集.北京,科学出版社,1992:125-139.
[11]Cangwere S K.A monograph on food selection inOrthoptera[J].Transactions of the American Entomological Society,1961,87:67-230.
[12]李广.亚洲小车蝗为害草场损失估计分析的研究[D].北京:中国农业科学院,2007.
[13]余鸣.蝗虫生态阈值初探[D].北京:中国农业科学院植物保护研究所,2006.
[14]Torell L A,Huddleston E W.Factors affecting the economic threshold for control of range-land grasshoppers[M]∥Capinera J L,Integrated Pest Management on Rangeland.Westview,Boulder,CO,1987:377-396.
[15]卢辉,韩建国,张泽华.锡林郭勒典型草原植物多样性和蝗虫种群的关系[J].草原与草坪,2008(3):21-28.
[16]卢辉,韩建国.典型草原三种蝗虫种群死亡率和竞争的研究[J].草地学报,2008,16(5):480-484.
[17]康乐,陈永林.草原蝗虫的时空异质性[M]∥草原生态系统研究,第4集.北京:科学出版社,1992:109-123.
[18]贺达汉,郑哲民.荒漠草原蝗虫营养生态位及种间食物竞争模型的研究[J].应用生态学报,1997,8(6):605-6110.
[19]Hansen R S,Ueckert D N.Dietary similarity of some primary consumers[J].Ecology,1970,51:640-648.
[20]Kang L,Chen Y L.Multimensional analysis of resource utilization in assemblages rangeland grasshoppers (Orthop.:Acrid.)[J].Entomologia Sinica,1994,1(3):264-282.
[21]Shelden J K,Rogers L E.Grasshopper food habits within a shrubsteppe community[J].Oecologia,1978,32:85-92.
[22]Ueekert D N.Seasonal dry weight composition in grasshopper diets on Colorado herbland[J].Annals of the Entomological Society of America,1968,61:1539-1544.
[23]卢辉,余鸣,张礼生.不同龄期及密度亚洲小车蝗取食对牧草产量的影响[J].植物保护,2005,31(4):55-58.
[24]康乐.放牧干扰下的蝗虫—植物相互作用关系[J].生态学报,1995,15(1):1-10.
