棉蚜抗药性及其化学防治
2013-09-28邵振润高希武
梁 彦, 张 帅, 邵振润, 高希武*
(1.中国农业大学昆虫学系,北京 100193;2.农业部全国农业技术推广服务中心,北京 100125)
棉蚜[Aphisgossypii(Glover)]又称瓜蚜,属半翅目蚜科,是一种世界性的杂食害虫,主要通过取食作物汁液和传播病毒病造成危害。棉蚜寄主范围广泛,据记载有116科900多种;棉蚜具有孤雌生殖和两性生殖两种方式,在北方露地、保护地和南方亚热带地区孤雌生殖是主要的繁殖方式,在寒冷季节和地区,可以迁飞至木槿、花椒等越冬寄主上进行繁殖,故其一年可以繁殖10~30代。目前,棉蚜的防治主要依赖于化学防治,随着杀虫药剂的广泛使用,棉蚜对有机氯类、有机磷类、拟除虫菊酯类、氨基甲酸酯类等杀虫药剂逐渐产生抗性。由于棉蚜发生量大,世代周期短,以及田间用药不合理,使得其抗药性发展迅速[1],已成为抗药性严重和难以治理的害虫之一。本文主要从棉蚜抗药性现状、抗药性机制以及棉蚜的化学防治等三个方面进行阐述。
1 棉蚜抗药性的发展历史与现状
1.1 棉蚜对有机磷类药剂的抗性
20世纪30年代,德国Schradev首先发现有机磷类杀虫药剂,50年代,美国氰胺公司推出了一个重要的低毒杀虫药剂——马拉硫磷。有机磷杀虫药剂多数属高毒或中等毒性,少数为低毒性;常用的有机磷杀虫药剂有敌百虫、马拉硫磷、毒死蜱等。该类杀虫药剂杀虫谱广,兼有触杀、胃毒和熏蒸3种不同的杀虫作用方式,并且具有高效速杀的性能,但由于大量单一的使用,在国内外棉蚜对不少有机磷类杀虫药剂都产生了严重的抗性。
1983年,在泰国棉蚜田间种群对乐果产生了抗性[2]。1992年在美国阿拉巴马地区,棉蚜对多种有机磷杀虫药剂均产生了抗性,其中包括磷铵、甲胺磷、砜吸磷、硫丙磷等,但抗性水平均不高[3];同时在密西西比地区也发现了棉蚜对有机磷药剂产生了抗性[4]。1994年,美国夏威夷地区棉蚜田间种群对乙酰甲胺磷产生了高达2 000倍以上的抗性[5]。1995年,美国加利福尼亚州出现了毒死蜱、杀扑磷的抗性,且抗性水平较高[6]。2000年,澳大利亚对马拉硫磷的抗性倍数为41~56倍,对丙溴磷抗性倍数为4.8~6.3倍[7]。2001年,欧洲的西班牙、荷兰、法国、希腊、意大利等国家,棉蚜对甲胺磷和砜吸磷产生了不同程度的抗性,抗性水平较高[8]。1996-2004年,巴基斯坦田间棉蚜种群对8种有机磷类药剂的抗性发生不同程度的变化,棉蚜对久效磷一直保持相对比较敏感的水平,在1998年抗性增长了9.8倍,到2004年降低到0.5倍;对乐果抗性也比较低,处于5.8~41倍;对丙溴磷抗性倍数为0.3~7.9倍;对毒死蜱抗性倍数为0.2~6.4倍;对喹硫磷抗性倍数为0.07~3.1倍;对甲基对硫磷抗性倍数为4.4~80倍,抗性呈上升趋势;对甲基嘧啶磷抗性倍数为15~1.4倍,抗性水平下降,对乙硫磷抗性1998年和1999年分别为215和921倍,到2004年降为4.3倍[9]。2008年,喀麦隆北部地区棉蚜对乐果产生了一定水平的抗性[10]。
1964年,国内首次报道了棉蚜对内吸磷(1059)、对硫磷(1605)分别产生了148和23倍的抗性[11]。1963-1964年,对河南各棉区棉蚜进行测定发现:棉蚜对内吸磷的抗性分别达到150.85、88.86、25.07、15.78倍[12]。1977年,山东高密县棉区的棉蚜对内吸磷和对硫磷的抗性从1963年的148倍和23倍上升到1 400倍和800倍[13]。1980年,国内棉田的棉蚜开始对甲基对硫磷产生了一定的抗性[14];1985-1999年,山东的滨州、菏泽、聊城、德州四个地区对氧化乐果的抗性倍数在5~80[15]。1988年,河北、北京等地棉蚜对马拉硫磷的抗性达357.8倍,对乐果抗性达128.6倍,敌敌畏83.5倍,乙酰甲胺磷72.1倍[16];同时,棉蚜对国内用量不大的磷铵也表现出抗性[17]。1989年,河南省棉区棉蚜对久效磷和马拉硫磷等有机磷类药剂则处于低水平的抗性[18];到1992年抗性监测显示河南省棉区棉蚜对久效磷和马拉硫磷的抗性水平明显增加。1995年棉蚜对久效磷抗性为34.16倍;对马拉硫磷的抗性则为55.36倍[19]。1990-1995年,陕西省棉区棉蚜伏蚜仅对久效磷产生5.2~9.8倍的抗性,而对其他有机磷杀虫药剂均处于敏感阶段[20]。1993年至1996年四川省棉蚜对马拉硫磷产生中等抗性,1996年抗性水平为17.9~45.0倍;1997年抗性有所下降,1998年回升,比1997年上升了18倍[21]。1999年,新疆北疆棉蚜种群对有机磷类杀虫药剂的抗性差异较大,抗性倍数为0.3~6.8,对甲基对硫磷和马拉硫磷处于敏感水平,抗性倍数分别为1.0和0.3倍,对马拉硫磷和久效磷处于低抗水平,抗性倍数分别为5.9和6.8倍;与1988年相比,到1999年这12年间棉蚜对有机磷类杀虫药剂的相对抗性上升了3.23~3.65倍[22]。2008年,我国山东地区棉蚜对马拉硫磷、毒死蜱、乐果、乙酰甲胺磷产生了不同程度的抗性[23]。
1.2 棉蚜对拟除虫菊酯类药剂抗性
拟除虫菊酯类药剂是依据天然除虫菊素的化学结构而仿制成的一类高效杀虫药剂。自20世纪七八十年代问世以来,由于其具有高效、广谱、低毒、低残留等特点而在世界各国得到了广泛的应用,但在使用后很快就出现了抗性问题,并且抗药性发展速度明显快于有机磷、氨基甲酸酯类药剂。
1992-1994年,美国德克萨斯州、阿拉巴马州、密西西比州、夏威夷州的棉蚜分别对氯氟氰菊酯、联苯菊酯、S-氰戊菊酯产生了不同程度的抗性,其中夏威夷州棉田的棉蚜对S-氰戊菊酯的抗性倍数达到390倍[3,5,24]。1997-2000年,巴基斯坦中部地区出现了对联苯菊酯、氟氯氰菊酯、高效氯氟氰菊酯、氯氰菊酯、溴氰菊酯、甲氰菊酯等拟除虫菊酯类药剂的抗性[25]。1998年,在澳大利亚新南威尔士州出现了联苯菊酯、高效氯氟氰菊酯抗性[26];2001年,该地区棉蚜对溴氰菊酯的抗性倍数为19倍,对联苯菊酯的抗性倍数为48倍[7]。2001年,欧洲的法国、意大利、希腊、荷兰、葡萄牙、西班牙等国棉蚜种群均出现了对氟氯氰菊酯的抗药性[8]。2003年,巴基斯坦棉蚜田间种群几乎对所有拟除虫菊酯类农药均产生了高水平抗性[27]。2008年,喀麦隆北部棉区出现了氯氰菊酯的抗药性,抗性倍数为41.2~473倍[10]。
1985年,我国山东菏泽、聊城、滨州、德州4个棉区棉蚜对氰戊菊酯抗性倍数达到30~370倍[15]。1988年,我国棉蚜表现出了对溴氰菊酯的抗性[17];北京、河北棉蚜对溴氰菊酯的抗性为401.2倍,对氰戊菊酯抗性为519.7倍[29]。1990年,山东德州的棉蚜种群对溴氰菊酯、氰戊菊酯的抗性分别达到6 113.0、12 576.8倍[30]。1995年陕西棉蚜对溴氰菊酯、氰戊菊酯及氯氟氰菊酯的抗性分别为75 127.91、8 341.25 和 3 122.45 倍[20]。1988-1999年间,新疆北疆棉蚜对菊酯类杀虫药剂抗性发展呈现不同的发展速度,氯氰菊酯的抗药性上升速度快于溴氰菊酯,相对上升了20.48倍[22]。2000年四川省棉区棉蚜对溴氰菊酯抗性最高(16.6~108.5倍),但是存在地区和年度间的差异[21]。1999-2004年,山东菏泽、聊城、滨州、德州地区对氰戊菊酯的抗性倍数为370~2 150倍[15]。2008年,山东地区出现了高效氰戊菊酯的抗药性[23]。
1.3 棉蚜对氨基甲酸酯类药剂的抗性
毒扁豆碱是最早发现的天然氨基甲酸酯类化合物,直到19世纪50年代,许多农药及相关企业相继推出了氨基甲酸酯类杀虫药剂产品,例如甲萘威等;这类药剂具有杀虫广谱、低毒、廉价、合成简单等特点,在随后短短的几年中得到了迅速开发,涕灭威、灭多威、克百威、丁硫克百威等一批氨基甲酸酯类杀虫药剂相继问世,并成为当时杀虫药剂领域的第二大类产品。但是,这些氨基甲酸酯类药剂都已受到了抗药性的影响。
1980年,英国首先报道了棉蚜对抗蚜威的抗药性[31];1988年,日本科学家也报道了棉蚜对抗蚜威的抗性[32];1991年,希腊棉田棉蚜种群对抗蚜威、唑蚜威、灭多威等分别产生了380、31和3倍的抗性;1992年报道了津巴布韦的棉蚜种群对抗蚜威和唑蚜威产生了74和57倍的抗性,对灭多威没有发现抗性[33]。但是1994年,美国夏威夷州发现了棉蚜对灭多威的抗药性,抗性倍数为9.2倍[5]。2001年,在欧洲的法国、西班牙、希腊、荷兰、意大利出现了针对抗蚜威的抗性[8]。1999-2003年,巴基斯坦田间棉蚜种群对甲萘威抗性为15~64.5倍;对灭多威抗性水平为1.0~9.1倍,逐年下降;1997-1999年对硫双威抗性为26~101倍,到2004年降为1.9倍;对呋线威抗性为0.5~1.5倍,处于敏感水平;对丁硫克百威抗性为0.4~1.2倍,很敏感[9]。2004-2008年,室内棉蚜抗性品系对抗蚜威的抗性倍数从227 倍上升到48 000 倍[34-35]。
1988年,克百威、甲萘威在国内棉蚜上出现了抗药性[17];1988年,河北、北京等地棉蚜对抗蚜威的抗性达7 887.7倍,表现出极高抗性[16]。1990-1995年陕西棉蚜对灭多威产生20.4倍的抗药性[20]。1996年四川地区棉蚜对灭多威的抗性已增长了7.0倍[21]。1988至1999年间,新疆北疆棉蚜对灭多威的抗药性发展较快,相对上升了20.48倍[22]。1999年,山东滨州、菏泽、聊城、德州出现了对丁硫克百威的抗药性,抗性倍数分别为9.05、9.77、8.81、9.26倍,到2004年,各地抗性倍数均有所下降[15];2008年,山东出现了对灭多威、克百威的交互抗性,抗性水平分别为22.4、33.2倍[23]。
1.4 棉蚜对新烟碱类药剂的抗性
新烟碱类杀虫药剂是防治刺吸式害虫如蚜虫等最有效的药剂之一。拜耳公司于1991年首先推出了新烟碱类化合物吡虫啉,是防治刺吸式口器害虫药剂发展中的一个“里程碑”。随后开发出了啶虫脒、噻虫嗪、噻虫啉等一系列的药剂。但是2000年前后,就发现了棉蚜对新烟碱类药剂的抗性。
2001年,在意大利的罗马、葡萄牙Poroa Isenta的棉蚜上,首先出现了吡虫啉抗性的报道[8];一年后,在我国山东泰山的棉蚜上也出现了相似的抗性[36];随后,棉蚜对吡虫啉的抗性迅速扩展,目前抗性已经扩展到山东省的德州、聊城、菏泽、滨州、泰安等地区[15,37]。除此之外,2004年,在我国山东聊城采集的棉蚜上也产生了对啶虫脒的抗性[15];最近几年,在澳大利亚的棉蚜种群对噻虫嗪产生抗药性[38]。2012年郭天凤等[39]对我国新疆主要植棉区的棉蚜种群进行了吡虫啉和啶虫脒2种药剂的抗性评价,结果发现,整个新疆植棉区棉蚜对吡虫啉的敏感性好于啶虫脒,部分地区棉蚜对啶虫脒已经产生了低水平的抗性。
1.5 棉蚜对有机氯类、环戊二烯类药剂的抗性
从20世纪30年代末开始,有机氯杀虫药剂由于其价格便宜、制造方便、对人畜口服毒性小,而得到快速的发展;但是在长期使用中发现,该类药剂在土壤、作物上残留较久,污染生态环境,威胁人类健康,为此世界上一些国家开始对如六六六、DDT等有机氯杀虫药剂进行限制和禁用。我国于1983年起,也正式停止了六六六、DDT的生产,因此之后未见其药剂产生抗性的报道;而在国外,2008年,喀麦隆的诺尔省棉蚜对DDT仍有抗性[10]。此外,在90年代初,美国密西西比州和夏威夷州出现了棉蚜对硫丹的抗药性,其中在夏威夷州的抗性倍数为3.6倍[5,24]。
2 棉蚜抗药性机制
随着杀虫药剂在田间的广泛使用,昆虫抗药性已成为限制化学防治有效性的重要因素,而对其抗药性机制的研究则是解决抗性问题的基础。通常害虫产生的抗药性策略主要有药剂对体壁的穿透性降低、害虫对药剂的代谢能力增加、杀虫药剂与害虫体内分子靶标结合能力降低、行为逃避减少药剂的接触剂量以及脂肪组织的储藏代谢、结合蛋白的隔离等。
2.1 文献报道的棉蚜抗药性机制类型
棉蚜的抗药性机制研究报道比较多。主要涉及表皮穿透性的降低、对药剂代谢能力增加(羧酸酯酶、P450等)、靶标敏感度降低(钠离子通道点突变、乙酰胆碱酯酶敏感度降低等)[40]。棉蚜中已经报道的涉及乙酰胆碱酯酶敏感度降低产生抗性的药剂有毒死蜱、内吸磷、乐果、甲胺磷、杀扑磷、氧化乐果、对硫磷、甲基对硫磷、磷胺、硫丙磷、甲萘威、丁硫克百威、抗蚜威、灭多威等;涉及乙酰胆碱受体变异产生抗性的药剂有啶虫脒、吡虫啉、噻虫嗪等;涉及钠离子通道点突变产生抗性的药剂有联苯菊酯、氟氯氰菊酯、氯氟氰菊酯、氯氰菊酯、溴氰菊酯、高效氰戊菊酯、甲氰菊酯;与GABA-氯离子通道复合体变异产生抗性的药剂有硫丹;与P450代谢增加产生抗性有关的药剂有乙酰甲胺磷、毒死蜱、马拉硫磷、氧化乐果、高效氰戊菊酯、克百威、灭多威等;与羧酸酯酶(酯酶)代谢增加或作为结合蛋白产生抗性的药剂有有机磷、氨基甲酸酯、拟除虫菊酯类药剂等(详见表1)。

表1 棉蚜在全世界抗性发生概况1)Table 1 Profile of A.gossypii resistance to insecticides in the world

续表1 Table 1(Continued)

续表1 Table 1(Continued)
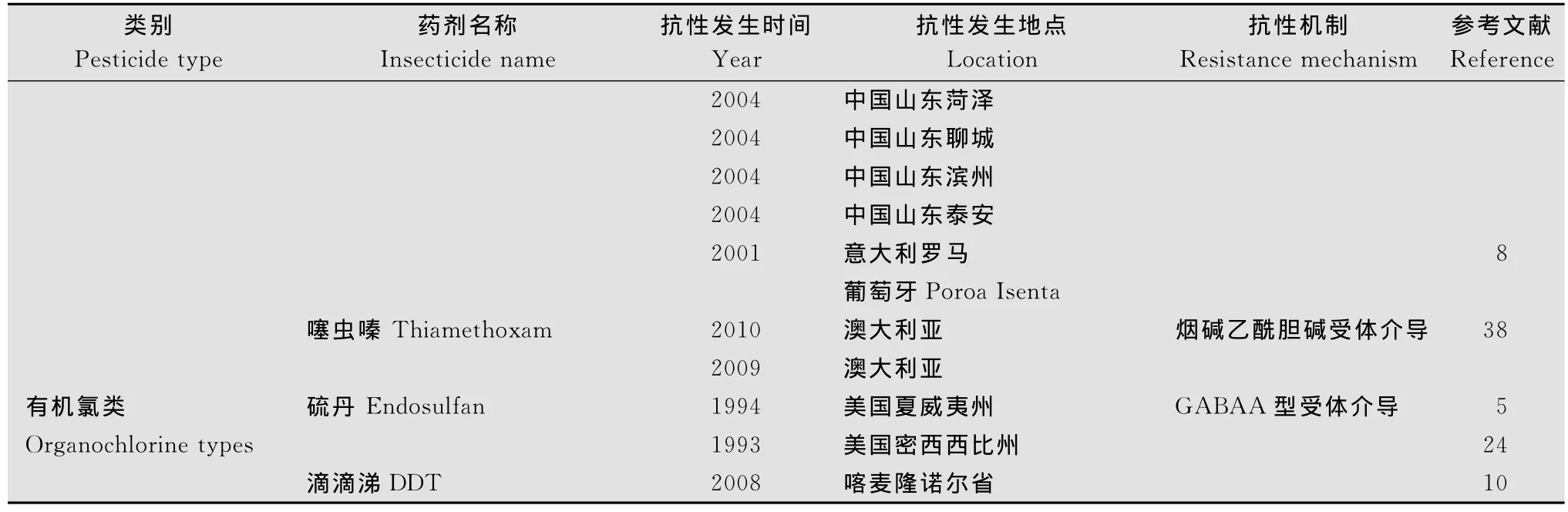
续表1 Table 1(Continued)
2.2 羧酸酯酶介导的棉蚜抗药性
羧酸酯酶(Car E)是昆虫体内重要的解毒酶系之一,能够催化水解脂族羧酸酯、芳族酯、芳族胺类及硫酯等化合物,在昆虫对杀虫剂的抗性中起着极为重要的作用[41]。
高希武等[42]通过乐果、氧化乐果、抗蚜威、氰戊菊酯、溴氰菊酯和氯氰菊酯等6种常用杀虫剂对8种蚜虫选择毒性的研究,发现棉蚜(瓜蚜)对药剂的敏感度明显低于其他蚜虫种。氰戊菊酯和溴氰菊酯对蚜虫的选择毒性与α-乙酸萘酯羧酸酯酶的活性具有明显的相关性,而与β-乙酸萘酯羧酸酯酶的活性则没有显著相关性。氯氰菊酯的选择毒性与上述两种酯酶的活性没有相关性。
研究发现久效磷、倍硫磷、甲萘威、马拉氧磷、虫螨磷对拟除虫菊酯类杀虫剂具有明显的增效作用,甲萘威、倍硫磷或马拉氧磷与氰戊菊酯混用(3∶1)对瓜蚜(棉蚜)的共毒系数分别为166.8、276.7和411.1;久效磷、倍硫磷或虫螨磷与溴氰菊酯以3∶1混用对瓜蚜的共毒系数分别为149.9、488.2和592.7。这些有机磷或氨基甲酸酯类杀虫剂对瓜蚜羧酸酯酶的活性具有明显的抑制作用,其抑制能力与对氰戊菊酯和溴氰菊酯的增效程度呈显著正相关。说明对蚜虫体内水解拟除虫菊酯的羧酸酯酶的抑制是主要的增效机制之一[43]。
高希武等用分光光度计终点测定法和酶标仪动力学测定法检测了3个不同抗性水平的棉蚜品系(R1,R2,R3)和1个敏感品系(S)的羧酸酯酶活性。发现S品系对β-乙酸萘酯(β-NA)的水解活性明显高于α-乙酸萘酯(α-NA),而抗性品系相反,对α-NA的活性明显高于β-NA。对α-NA水解的活性在不同品系间差异较大(近60倍),而对β-NA水解的活性差异小于对α-NA,最大约为14倍。用酶标仪动力学测定法研究表明,4个棉蚜品系间羧酸酯酶活性也存在明显的差异,S、R1、R2和R3分别为38、85、198和762 m OD/(min·头);其他研究中均证明酶动力学方法测定结果的可靠性[44-46]。
孙鲁娟等[47-48]用α-乙酸萘酯(α-NA)、α-丁酸萘酯(α-NB)、α-磷酸萘酯(α-NP)和β-磷酸萘酯(β-NP)作底物比较研究了氧化乐果抗性和敏感品系棉蚜羧酸酯酶的比活力、米氏常数(Km)和最大反应速度(Vmax)等有关的动力学常数。以α-NA和α-NB作底物时,抗性品系棉蚜的比活力显著低于敏感品系;以α-NP和β-NP作底物时,两个品系棉蚜的比活力、Km和Vmax没有明显差异。酯酶同工酶分析同样表明了抗性品系棉蚜羧酸酯酶活力低于敏感品系棉蚜。利用RT-PCR扩增的方法克隆了氧化乐果抗性(RR)和敏感(WS和SS品系)品系棉蚜的羧酸酯酶基因,抗性品系与WS敏感品系比较,有8个核苷酸的突变,其中产生了4个氨基酸的突变;与新疆敏感品系(SS)比较有6个核苷酸的突变,其中产生了4个与WS品系相同的氨基酸突变(苏氨酸→甲硫氨酸,天冬酰胺→赖氨酸,甘氨酸→天冬氨酸,丝氨酸→苯丙氨酸)。SS品系和WS品系之间有6个核苷酸的差异,但都没有引起氨基酸的突变。以羧酸酯酶基因为探针,用限制性内切酶(HpaII、Msp I)完全酶解氧化乐果抗性和敏感品系棉蚜总DNA进行杂交。杂交图谱没有显著不同,表明抗性品系棉蚜没有基因扩增现象,而且在棉蚜的羧酸酯酶基因中没有甲基化现象。在另一个研究中以α-乙酸萘酯(α-NA)为底物,比较氧化乐果敏感和抗性品系棉蚜羧酸酯酶的比活力,发现抗性品系羧酸酯酶比活力明显小于敏感品系。对这两个品系的羧酸酯酶基因进行了克隆,通过对抗性和敏感品系羧酸酯酶基因核苷酸序列及推导的氨基酸序列比较,发现抗性品系有4个氨基酸发生了替代,对其蛋白质三维结构分析推测His104—Arg的替代位于其活性中心[49]。
用α-乙酸萘酯作底物,测定羧酸酯酶活性,分析其活性与有机磷抗性的关系。研究发现:两种品系棉蚜的羧酸酯酶对杀螟硫磷的水解能力很小,其中抗性品系的酶活力越高,其结合能力也越高;此外,杀螟硫磷的结合能力也会由于过量底物的存在而降低。因此,推测α-乙酸萘酯与杀螟硫磷很可能具有相同的作用位点[50]。
曹传旺等[51]利用溴氰菊酯和高效氯氰菊酯的荧光类似物作为底物,研究了抗性和敏感棉蚜品系对菊酯类药剂代谢的差异。应用这两种底物水解后生成具有荧光化合物的特性,测定了不同品系棉蚜羧酸酯酶的代谢活性。结果表明,氧化乐果棉蚜抗性和敏感品系羧酸酯酶对溴氰菊酯报告荧光底物的代谢活性分别为 10.0 pmol/min·mg pro.和3.4 pmol/min·mg pro.;对高效氯氰菊酯报告荧光底物的代谢活性分别为4.0 pmol/min·mg pro.和2.4 pmol/min·mg pro.,抗性品系羧酸酯酶对溴氰菊酯和高效氯氰菊酯报告荧光底物的代谢活性分别为敏感品系的2.9和1.7倍;溴氰菊酯棉蚜抗性和敏感品系羧酸酯酶对溴氰菊酯报告荧光底物的代谢活性分别为7.6和6.2 pmol/min·mg pro.;对高效氯氰菊酯报告荧光底物的代谢活性分别为9.3和5.2 pmol/min·mg pro.,抗性品系羧酸酯酶对溴氰菊酯和高效氯氰菊酯报告荧光底物的代谢活性分别为敏感品系的1.2和1.8倍。这种衍生的报告荧光底物能够用来检测抗性棉蚜羧酸酯酶的水解活性,表明羧酸酯酶可能参与棉蚜对溴氰菊酯和氧化乐果抗性的形成。
Cao等[52]运用实时荧光定量RT-PCR对氧化乐果抗性和敏感品系羧酸酯酶m RNA表达水平进行了研究,抗性品系棉蚜羧酸酯酶基因mRNA相对表达量为敏感品系的4.54倍,表明羧酸酯酶基因的过量表达可能参与氧化乐果抗性机制的形成。溴氰菊酯抗性品系(DRR)棉蚜羧酸酯酶基因mRNA相对表达量为敏感品系的6.28倍。阿瓦提抗性品系(AWT)羧酸酯酶基因m RNA相对表达量为敏感品系的6.61倍。RT-PCR克隆获得的阿瓦提抗性和敏感品系羧酸酯酶基因cDNA均为1 581 bp,编码526个氨基酸,两品系在核苷酸水平上具有99.94%的同源性,氨基酸水平上具有99.81%的同源性;与敏感品系相比,抗性品系中仅有一个碱基发生了变化,导致一个氨基酸的变化(Ala87-Val),但该突变位点不位于活性中心,两品系羧酸酯酶活性中心和信号域均未发生变化,由此推断棉蚜溴氰菊酯抗性主要由羧酸酯酶基因转录水平的增加引起,Ala87-Val突变可能不参与抗性机制[53]。
Pan等[54]应用实时荧光定量PCR研究棉蚜马拉硫磷抗性和敏感品系体内羧酸酯酶mRNA相对表达量和DNA相对拷贝数的变化。结果表明抗性品系羧酸酯酶基因m RNA表达量是敏感品系的1.99倍,而抗性品系羧酸酯酶基因DNA拷贝数是敏感品系的4.42倍。这说明在棉蚜对马拉硫磷产生抗性过程中,体内羧酸酯酶基因DNA水平拷贝数的增加和mRNA转录水平的提高与马拉硫磷的代谢有关。同时通过RT-PCR扩增得到羧酸酯酶全长基因序列,对比分析抗性和敏感品系羧酸酯酶基因序列,抗性品系氨基酸序列发现2个突变:K(Lys)14Q(Gln);N(Asn)354D(Asp)。在所检测的9个抗性品系样品中其中8个样品均发生了这两个突变,F14Q突变导致了MRR的亲水性大于MSS的亲水性,N354D突变导致了MRR的亲酯性大于MSS的亲酯性。这可能与棉蚜对马拉硫磷抗性有关。同时发现羧酸酯酶mRNA存在选择性剪切现象,其中剪切变异型羧酸酯酶和正常羧酸酯酶基因相比较,变异型剪切了60 bp碱基,少20个氨基酸。
Chang等[55]利用溴氰菊酯抗性品系(DRR)和敏感品系(DSS),研究了DEF(酯酶抑制剂)的增效作用以及其对溴氰菊酯抗性品系羧酸酯酶的抑制作用。结果发现:DRR是DSS抗性的23 900倍,对联苯菊酯和醚菊酯的交互抗性分别是7 560和99倍;当DEF预处理15 h时,DEF增加了溴氰菊酯和联苯菊酯的毒性,而醚菊酯的毒性没有增加;DEF显著抑制了DRR中羧酸酯酶的活性,而DSS没有明显变化;对DRR活体抑制试验在15 h时达到最大抑制,24 h后又恢复到原来的活性。证明了棉蚜体内的羧酸酯酶是其对溴氰菊酯产生抗药性的重要机制之一。
为了明确植物次生物质对昆虫解毒酶的影响作用,利用饲喂法和2-十三烷酮熏蒸法研究了植物次生物质2-十三烷酮和槲皮素对棉蚜羧酸酯酶的诱导作用。结果表明,取食含0.01%槲皮素的饲料棉蚜羧酸酯酶的活性最大,均与对照差异显著。用2-十三烷酮熏蒸处理24 h,以α-NA为底物测定棉蚜羧酸酯酶的活性在处理后48 h达到最大,为对照的2.5倍。饲喂法处理槲皮素对棉蚜羧酸酯酶的诱导作用具有显著的剂量效应;亚致死剂量2-十三烷酮熏蒸对棉蚜羧酸酯酶的诱导作用具有显著的时间效应[56]。
不同寄主植物对棉蚜羧酸酯酶活性有一定的影响。高希武[57]对不同棉花品种间棉蚜种群羧酸酯酶活性进行了测定。结果表明,在试验的7个棉花品种中,取食‘中棉12’叶片的棉蚜α-乙酸萘酯(α-NA)羧酸酯酶活性是取食‘泾阳鸡脚棉’种群的5.7倍。β-乙酸萘酯(β-NA)羧酸酯酶的活性以取食‘方渑双无’的棉蚜最高,是取食‘BR-S-10’棉蚜的4.5倍。在取食‘方渑双无’叶片的棉蚜中,羧酸酯酶活性大于2.5μmol/min·mg pro.的个体占28.3%(α-NA)和67.1%(β-NA),而在取食‘BR-S-10’的种群中仅占2.6%(α-NA)和1.3%(β-NA)。取食‘中棉12’的棉蚜对β-NA与α-NA活性的比值为0.79,而取食其他棉花品种的种群β-NA与α-NA活性比值均大于1。用‘中棉12’饲养的棉蚜对α-NA和β-NA活性均低于种群平均值的个体(CE1型)占26%;均高于平均值的个体(CE2型)占19%;对α-NA活性高于平均值,对β-NA活性低于平均值的个体(CE3型)占31%;对α-NA活性低于平均值,对β-NA活性高于平均值的个体(CE4型)占24%。取食其他6个棉花品种的棉蚜中,CE1和CE2型的比例分别为49.4%~64.0%和23.4%~47.2%,明显高于取食‘中棉12’的种群,而CE3和CE4型个体仅占0~7.6%和3.4%~10.4%,明显低于取食‘中棉12’的种群。说明寄主植物对棉蚜羧酸酯酶的量和质均有影响。
聚丙烯酰胺凝胶电泳图谱显示出,棉蚜种群中大多数个体都具有6条基本的酯酶活性带(E-2—E-7),但是,在北京海淀区东北旺抗性种群中发现了具有9条带的个体,其中 E-1染色最浅,E-8、E-9染色较深。由于该蚜对拟除虫菊酯的抗性水平与其酯酶活性密切相关,确定每毫克蛋白质15 min水解α-乙酸萘酯大于40μmol的蚜虫个体频率在种群中大于11%时为抗性种群[60]。
3 棉蚜的化学防治展望
棉蚜的化学防治是否奏效受到许多因子的影响,其中抗药性机制的差异影响比较大。在我国棉蚜田间种群中除了上述介绍的羧酸酯酶介导的抗性以外,药剂的穿透性降低、ACh E敏感度降低[61-62]、钠离子通道点突变[63]等都有报道。另一方面随着转Bt基因棉的种植,非Bt棉靶标害虫种群数量有上升趋势,棉蚜是转Bt基因棉田发生危害的刺吸性害虫之一。在实验室研究表明,棉蚜分别用转Bt基因棉和亲本常规棉饲养40代后,饲养于转Bt基因棉上的棉蚜体内能检测到痕量的Bt蛋白。与亲本常规棉相比,转Bt基因棉上棉蚜寿命有延长的趋势,但是生命表参数无显著差异[59]。另外也发现不同体色的棉蚜对杀虫药剂的敏感度存在差异。黄瓜上体色为绿色的棉蚜种群对氧乐果、抗蚜威和氰戊菊酯的耐性明显高于黄色棉蚜种群[58]。
在我国棉蚜田间种群对药剂的抗性发展速度较快,到目前为止几乎对所有使用的药剂均产生了抗性(表2)。棉蚜的化学防治除了合理地选择药剂品种外,重要的方面是掌握施药时间和施药方法,做到科学用药。例如,棉蚜一般选择在叶片的背部、嫩梢和嫩茎进行为害,喷药时喷头朝上,自下而上,喷匀打透;交替用药,降低用药频率,避免同一种药连续使用(尤其是菊酯类药剂),以防止或减缓抗药性的产生。棉蚜发生初期应采取点片防治与全面防治相结合,尽量选用高效低毒、对天敌杀伤力小、环境友好型的化学药剂。具体来说,当田间棉蚜呈零星或点片发生时,可结合田间管理,进行挑治,不必全田喷药,发生区域可选用吡虫啉、啶虫脒等新烟碱类或联苯菊酯类等药剂进行防治。目前新烟碱类药剂对棉蚜活性比较高,菊酯类药剂中联苯菊酯、三氟氯氰菊酯、溴氰菊酯等活性仍高于其他菊酯类药剂。

表2 我国棉蚜抗药性发展历史Table 2 History of A.gossypii resistance to insecticides in China
另一方面就是根据作物的生长发育状况、棉蚜种群数量以及天敌的影响等方面的因素,因地制宜地合理制定棉蚜田间化学防治的指标。早在20世纪80年代,我国就已经建立了一个针对化学防治的阈值(防治指标),可以减少50%杀虫药剂的使用[64]。
棉蚜的防治策略应以农业防治为主,化学防治为辅,兼以生物防治和物理防治。运用有害生物综合治理(Integrated Pest Management,IPM)的原则和方法,将棉蚜的种群数量控制在经济阈值以内,做到统筹安排、合理防治,从而建立一个良好的田间生态系统。
[1]李飞,韩召军,吴智锋,等.我国棉蚜抗药性研究现状[J].棉花学报,2001,13(2):121-124.
[2]Wangboonkong S.The cotton aphid insecticides[C].First Conference of Entomology and Zoology.Thailand,1983:290-296.
[3]Kerns D L,Gaylor M J.Insecticide resistance in field populations of the cotton aphid(Homoptera:Aphididae)[J].Journal of Economic Entomology,1992,85:1-8.
[4]O’Brien D J,Abdel-Aal M A,Ottea J A,et al.Relationship of insecticide resistance to carboxylesterases inAphisgossypii(Homoptera:Aphididae)from midsouth cotton[J].Journal of Economic Entomology,1992,85:651-657.
[5]Hollingsworth R G,Tabashnik B E,Ullman D E,et al.Resistance ofAphisgossypii(Homoptera:Aphididae)to insecticides in Hawaii:spatial patterns and relation to insecticide use[J].Journal of Economic Entomology,1994,87:293-300.
[6]Grafton-Cardwell E E,Vehrs S C.Monitoring for organophosphate-and carbamate-resistant armored scale(Homoptera:Diaspididae)in San Joaquin Valley Citrus[J].Journal of Economic Entomology,1995,88:495-504.
[7]Herron G A,Powis K,Rophail J,et al.Insecticide resistance inAphisgossypiiGlover(Hemiptera:Aphididae),a serious threat to Australian cotton[J].Australian Journal of Entomology,2001(40):85-91.
[8]Nauen R,Elbert A.European monitoring of resistance to insecticides inMyzuspersicaeandAphisgossypii(Hemiptera:Aphidiae)with special reference to imidacloprid[J].Bulletin of Entomological Research,2003,93(1):47-54.
[9]Ahmad M,Arif M I.Susceptibility of Pakistani populations of cotton aphidAphisgossypii(Homoptera:Aphididae)to endosulfan,organophosphorus and carbamate insecticides[J].Crop Protection,2008,27:523-531.
[10]Carletto J,Martin T,Vanlerberghe-Masutti F,et al.Insecticide resistance traits differ among and within host races inAphisgossypii[J].Pest Management Science,2010,66:301-307.
[11]龚坤元,张桂林,翟桂英.棉蚜对“1059”抗药性的测定[J].昆虫学报,1964,13(1):1-8.
[12]石键,崔光先,郑港庆.河北棉区棉蚜对内吸磷抗药性的调查[J].昆虫知识,1965(2):329-330.
[13]孙耘芹,冯国蕾,张桂林,等.棉蚜对有机磷杀虫剂抗性的生化机制[J].昆虫学报,1987,30(1):13-19.
[14]Gong K Y,Ahng G L,Sun Y Q.The chemical cotton aphids and the development of resistance in northern cotton areas in China during the last thirty years[C].International Congress of Entomology,Kyoto,1980.
[15]Wang K Y,Guo Q L,Xia X M,et al.Resistance ofAphis gossypii(Homoptera:Aphididae)to selected insecticides on cotton from five cotton production regions in Shandong,China[J].Journal of Pesticide Science,2007,32:372-378.
[16]郑炳宗,高希武,王政国,等.瓜-棉蚜对有机磷及氨基甲酸酯杀虫剂抗性机制研究[J].植物保护学报,1989,16(2):131-138.
[17]Tang Z H,Gong K Y,You Z P.Present status and countermeasures of insecticide resistance in agricultural pests in China[J].Pesticide Science,1988,23:189-198.
[18]吴孔明.河南省棉区棉蚜对三种杀虫药剂抗性测定[J].中国棉花,1989(3):40-42.
[19]胡冠芳,陈明,张新瑞,等.甘肃敦煌棉蚜抗药性测定[J].西北农业学报,2000,9(2):45-48.
[20]王兴林,冯俊涛,杨崇珍,等.陕西棉蚜抗药性监测[J].西北农业学报,1997,6(3):57-60.
[21]彭丽年,何树林,杨庭权,等.四川地区棉铃虫和棉蚜抗药性监测及分析[J].棉花学报,2002,14(2):62-64.
[22]马志卿,江志利,陈安良,等.新疆北疆棉蚜抗药性测定及发展动态分析[J].西北农林科大学学报,2001,29(2):80-82.
[23]Shang Q,Pan Y,Fang K,et al.Biochemical characterization of acetylcholinesterase,cytochrome P450 and cross-resistance in an omethoate-resistant strain ofAphisgossypiiGlover[J].Crop Protection,2012,31:15-20.
[24]Hardee D D,AinsworthJ M.Cotton aphid(Homoptera:Aphididae):effect of in-furrow insecticides on pesticide resistance[J]Journal of Economic Entomology,1993,86:1026-1029.
[25]Ahmad M,Arif M I,Denholm I.High resistance of field populations of the cotton aphidAphisgossypiiGlover (Homoptera:Aphididae)to pyrethroid insecticides in Pakistan[J].Journal of Economic Entomology,2003,96:875-878.
[26]Herron G A,Powis K,Rophail J.Baseline studies and preliminary resistance survey of Australian populations of cotton aphidAphisgossypiiGlover(Hemiptera:Aphididae)[J].Australian Journal of Entomology,2000,39:33-38.
[27]Ahmad M,Arif M I,Denholm I.High resistance of field populations of the cotton aphidAphisgossypiiGlover (Homoptera:Aphididae)to pyrethroid insecticides in Pakistan[J].Journal of Economic Entomology,2003,96:875-878.
[28]慕立义,王开运.我国棉花蚜虫对菊酯类农药及呋喃丹抗药性调查与研究[J].农药,1986(2):1-6.
[29]郑炳宗,高希武,王政国,等.北京及河北地区北部瓜-棉蚜对拟除虫菊酯抗药性的研究初报[J].植物保护学报,1988,15(1):55-61.
[30]王开运,慕立义,张世安,等.六种蚜虫对溴氰菊酯等杀虫剂的抗性及毒力选择性[J].农药,1992,31(3):2-5.
[31]Furk C,Powell D F,Heyd S.Pirimicarb resistance in the mel-on and cotton aphid,AphisgossypiiGlover[J].Plant Pathology,1980,29:191-96.
[32]Takada H,Murakami Y.Esterase variation and insecticide resistance in JapaneseAphisgossypii[J].Entomologia Experimentalis et Applicata,1988,48:37-41.
[33]Moores G D,Gao X W,Denholm I,et al.Characterisation of insensitive acetylcholinesterase in insecticide resistant cotton aphids,AphisgossypiiGlover(Homoptera:Aphididae)[J].Pesticide Biochemistry and Physiology,1996,56:102-110.
[34]Benting J,Nauen R.Biochemical evidence that an S431F mutation in acetylcholinesterase-1 ofAphisgossypiimediates resistance to pirimicarb and malathion[J].Pest Management Science,2004,60:1051-1055.
[35]Bingham G,Gunning R V,Delogu G,et al.Temporal synergism can enhance carbamate and neonicotinoid insecticidal activity against resistant crop pests[J].Pest Management Science,2008,64:81-85.
[36]Wang K Y,Liu T X,Yu C H,et al.Resistance ofAphisgossypii(Homoptera:Aphididae)to fenvalerate and imidacloprid and activities of detoxification enzymes on cotton and cucumber[J].Journal of Economic Entomology,2002,95:402-407.
[37]Shi X,Jiang L,Wang H,et al.Toxicities and sublethal effects of seven neonicotinoid insecticides on survival,growth and reproduction of imidacloprid-risistant cotton aphid,Aphis gossypii[J].Pest Management Science,2011,67:1528-1533.
[38]Herron G,Wilson L.New findings of insecticide resistance inTriatomainfestans(Heteroptera:Reduviidae)from the Gran Chaco[J].Australian Journal of Entomology,2011,50:93-98.
[39]郭天凤,马野萍,丁荣荣,等.新疆主要植棉区棉蚜对吡虫啉和啶虫脒的抗性评价[J].中国棉花,2012,39(12):4-5.
[40]Liu Nannan,Zhu Fang,Xu Qiang,et al.Behavioral change,physiological modification,and metabolic detoxification:mechanisms of insecticide resistance[J].昆虫学报,2006,49(4):671-679.
[41]孙鲁娟,高希武,郑炳宗.棉蚜抗药性现状及其分子机制研究[M]∥中国植物保护学会.面向21世纪的植物保护发展战略.北京:中国科学技术出版社,2001:481-484.
[42]高希武,王政国,郑炳宗,等.六种常用杀虫剂对八种蚜虫的选择毒性[J].昆虫学报,1990,33(3):274-279.
[43]高希武,郑炳宗.几种农药对蚜虫羧酸酯酶的抑制和拟除虫菊酯的增效[J].北京农业大学学报,1991,17(4):89-94.
[44]高希武,郑润勇,宁世民,等.棉蚜不同品系羧酸酯酶的酶标仪动力学测定研究[J].北京农业大学学报,1997,2(5):59-63.
[45]高希武,胡熳华.应用酶标仪动力学方法监测棉蚜的抗药性[J].昆虫知识,1998,35(1):17-19.
[46]赵颖,高希武.棉蚜不同抗性品系羧酸酯酶比较[J].植物保护学报,1997,24(4):351-355.
[47]Sun L,Zhou X,Zhang J,et al.Polymorphisms in a carboxylesterase gene between organophosphate-resistant and-susceptibleAphisgossypii(Homoptera:Aphididae)[J].Journal of Economic Entomology,2005,98:1325-1332.
[48]孙鲁娟,高希武,郑炳宗.棉蚜抗氧化乐果品系及敏感品系羧酸酯酶性质的比较[J].昆虫学报,2002,45(6):724-727.
[49]郭惠琳,高希武.棉蚜抗氧化乐果品系的羧酸酯酶基因突变[J].昆虫学报,2005,48(2):194-202.
[50]Suzuki K,Hama H,Konno Y.Carboxylesterase of the cotton Aphid,AphisgossypiiGlover(Homoptera:Aphididae).responsible for fenitrothion resistance as a sequestering protein[J].Applied Entomology and Zoology,1993,28:439-439.
[51]曹传旺,史雪岩,姜辉,等.运用α-氰代酯荧光底物检测抗性棉蚜羧酸酯酶代谢活性[J].昆虫学报,2009,52(3):261-266.
[52]Cao Chuanwang,Zhang Jing,Gao Xiwu,et al.Overexpression of carboxylesterase gene associated with organophosphorous insecticide resistance in cotton aphids,Aphisgossypii(Glover)[J].Pesticide Biochemistry and Physiology,2008,90:175-180.
[53]Cao Chuanwang,Zhang Jing,Gao Xiwu,et al.Differential mRNA expression levels and gene sequences of carboxylesterase in both deltamethrin resistant and susceptible strains of the cotton aphid,Aphisgossypii[J].Insect Science,2008,15(3):209-216.
[54]Pan Yiou,Guo Huilin,Gao Xiwu.Carboxylesterase activity,c DNA sequence,and gene expression in malathion susceptible and resistant strains of the cotton aphid,Aphisgossypii[J].Comparative Biochemistry and Physiology Part B:Biochemistry and Molecular Biology,2009,152(3):266-270.
[55]Chang J,Cao C W,Gao X W.The effect of pretreatment with S,S,S-tributyl phosphorotrithioate on deltamethrin resistance and carboxylesterase activity inAphisgossypii(Glover)(Homoptera:Aphididae)[J].Pesticide Biochemistry and Physiology,2010,98:296-299.
[56]吕敏,孙婳婳,王丽红,等.植物次生物质对棉蚜谷胱甘肽S-转移酶和羧酸酯酶活性的诱导作用[J].中国农学通报,2012,28(3):253-256.
[57]高希武.寄主植物对棉蚜羧酸酯酶活性的影响[J].昆虫学报,1992,35(3):267-272.
[58]高希武,郑炳宗.不同体色瓜蚜耐药性差异及其生化机制[J].植物保护学报,1991,18(2):181-185.
[59]薛丽,肖达,卢延,等.转基因棉对棉蚜生长发育及其主要代谢物质含量的影响[J].华北农学报,2011,26(4):115-120.
[60]高希武,郑炳宗.生物化学法监测瓜-棉蚜田间种群的抗药性[J].植物保护学报,1990,17(4):373-377.
[61]高希武,郑炳宗.北京地区蚜虫乙酰胆碱酯酶(ACh E)的研究[J].北京农业大学学报,1990,16(1):85-89.
[62]高希武,王荣京,郑炳宗.四种蚜虫AChE的活性及其对抑制剂的敏感度[J].昆虫知识,1990,27(4):217-218.
[63]孙鲁娟,高希武,郑炳宗.氰戊菊酯抗性和敏感品系棉蚜部分钠离子通道基因的克隆[J].中国农业科学,2003,36(11):1301-1305.
[64]张永孝,赵之刚,曹赤阳.棉蚜为害损失与防治指标的研究[J].植物保护学报,1982(4):229-236.
[65]钦俊德.中国昆虫毒理学、昆虫病理学三十年[J].昆虫学报,1979,22(3):249-256.
[66]上海昆虫研究所昆虫抗药性研究组,上海川沙县张江公社植物保护站,安徽霍山县植物保护站.氧乐果防治抗性棉蚜的试验[J].昆虫学报,1977,20(4):479-481.
