长江流域清江胡家溪四种蜉蝣优势种的周年生产量及其动态*
2013-09-25温芳妮王利肖李晓宇闫云君
江 晶,温芳妮,王利肖,邱 爽,李晓宇,闫云君
(华中科技大学生命科学与技术学院,武汉430074)
大型底栖动物是河流生态系统中重要的生物类群之一,能加速水底碎屑分解,促进泥水界面物质交换和水体自净,是河流生态系统物质循环和能量代谢的重要参与者,也是人们了解河流生态系统结构和功能的关键类群之一.此外,由于底栖动物的生命周期相对较长、行动缓慢、物种丰富,且不同种类底栖动物对环境条件的适应性和对污染等不利因素的耐受力及敏感程度不同,因此作为重要的指示生物已广泛应用于水质及环境监测中,起着水下哨兵的作用[1].因此,河流大型底栖动物群落结构与功能研究已成为生态学的热点领域之一[2].然而,在我国,河流生态学研究严重滞后,与国际先进水平存在较大差距,目前仅有少许对河流功能研究的报道[3-5].
蜉蝣目昆虫广泛存在于各级河流中,在河流生态系统中起着重要作用[1-2].目前有关该类群的研究工作多见于国外报道,国内有关工作主要集中在分类学和水质监测方面[6],而有关蜉蝣在河流生态系统中的生态功能及营养动态则鲜见报道.由于蜉蝣是河流生态系统食物链中极为重要的环节,对维系群落食物网的复杂性具有十分重要作用.因此,研究蜉蝣类群的生态学功能对了解和掌握我国河流生态系统的结构与功能特点具有重要意义.本文于2006-2007年对清江上游流域一二级河流(胡家溪)大型底栖动物群落进行了为期一周年的调查研究,探讨4种蜉蝣优势种的种群动态和周年生产量及动态变化.
1 工作方法
1.1 采样点和采样时间
胡家溪是长江支流清江的一条二级河流,位于长阳土家族自治县境内(30°27'~30°28'N,111°03'~111°35'E),流域面积约50 km2,于胡家窝附近注入清江,距离隔河岩水电站约8 km.胡家溪两侧为山脉,植被覆盖率较高,郁闭度较高,河两岸着生灌木和杂草.流域地区为典型亚热带气候,多年平均降水量约1460 mm,全年水温变化范围在10~28℃之间.

图1 胡家溪及采样点分布Fig.1 Distribution of sampling sites at Hujiaxi Stream
根据河流微生境的类型(底质、水流速度、水生植物等),从上游至下游选择了6个采样点(图1),分别是:S1:河边生长着茂密的灌木和杂草,河面较窄,约1~2 m宽,且部分被遮蔽着,水流较缓,底质为粗沙;S2:有一支流汇入其中,河面较窄,约2 m宽,水流较缓,河底以细沙为主;S3:在一个落差约0.5 m的小瀑布下方,水流较急,河底以布满了苔藓的巨石为主,附近10 m处有一片小竹林;S4:位于一堰塘下游浅水区,地势平缓,水流缓慢,底质为石灰岩,河边杂草较少;S5:河面变宽,河宽约3 m,河底以卵石为主,腐殖质较多,水流较急;S6:位于河口,一排污口的下游,河宽约4 m,水流较急,河底碎石较多.胡家溪河水的理化性质见表1.采样时间一般为每月的5日左右,为期一年,即2006年4月至2007年3月.
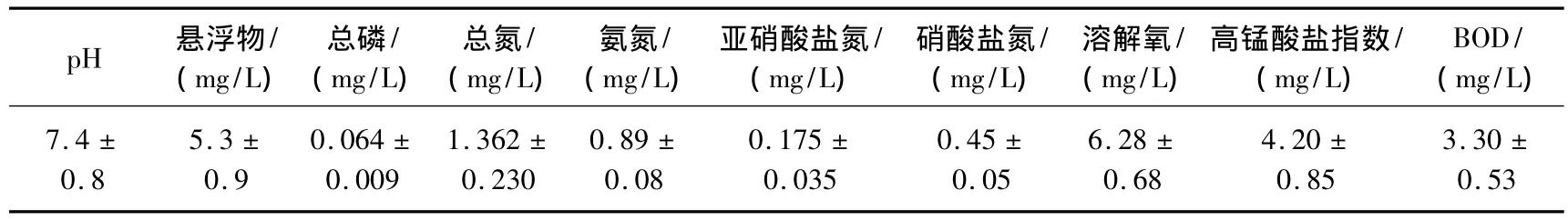
表1 胡家溪的理化性质Tab.1 Physico-chemical characteristics of Hujiaxi Stream
1.2 标本采集及处理
定量采集使用60目的D型网和Surber网,每个采样点面积约为30 cm×30 cm,每样点1次,1个重复.样品经60目铜筛筛洗后,置于白色解剖盘中分检,标本用10%的福尔马林固定保存.
1.3 生活史
根据各月种群的龄期分布频率了解其发育动态,从而推知种群生活史状况[7-8].
1.4 生产量的测算
采用体长(龄期)频率法(size(instar)-frequency method)测算其周年生产量[9-10],其计算公式为:
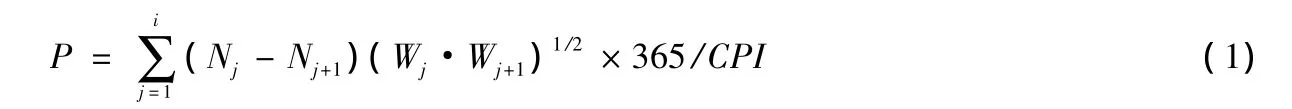
式中,CPI(Cohort Production Interval)为一个同龄组完成生活史所需时间.在此,忽略矫正系数i(Pe/P),令其等于1,原因是Pe/P在实际计算中非常困难,而且造成的误差极小[11].
1.5 生产量的动态
按照Benke等[8]的方法,测算各同龄组的P/B(生产量/生物量)系数,然后计算出日P/B系数,再根据各采样时段的现存量,求出各时段的生产量动态.生产量的时间重叠采用比例相似系数(Proportional Similarity index,PS)[12]表示为:

式中,PSab表示a、b两物种比例相似性,n表示采样间隔数,Pa,i表示 a物种第 i间隔生产量所占比例,Pb,i表示b物种第i间隔生产量所占比例.
2 结果
2.1 种群动态
小裳蜉种群密度和生物量(湿重,以下同)的变化趋势基本相同,周年平均值分别为407 ind./m2和1.00 g/m2.密度在2006年4月达到峰值(1156 ind./m2),生物量则在2006年5月达到峰值(2.40 g/m2),密度和生物量在其它各月均保持较低水平(图2a).
扁蜉种群密度和生物量的变化趋势较为不同,周年平均值分别为150 ind./m2和0.37 g/m2.密度在2007年3月出现峰值(256 ind./m2),明显高于其它各月;生物量则出现两次峰值,分别在4月(0.94 g/m2)和次年3月(1.05 g/m2),其中次年3月为主峰值(图2b).

图2 胡家溪小裳蜉(a)、扁蜉(b)、四节蜉(c)和细蜉(d)的现存量周年动态Fig.2 Annual variations of standing stock of Leptophlebia sp.(a),Electrogena sp.(b),Indobaetis sp.(c)and Caenis sp.(d)in Hujiaxi Stream
四节蜉种群密度和生物量的变化较为一致.密度出现两次峰值,即在11月达到主峰值(404 ind./m2),次年2月达到次峰值(385 ind./m2);生物量也出现两次峰值,分别在次年1月(0.56 g/m2)和次年2月(0.61 g/m2).四节蜉密度和生物量的周年平均值分别为 232 ind./m2,0.30 g/m2(图 2c).
细蜉种群密度和生物量的变化几乎完全同步,密度出现三次峰值,即在4月达到主峰值(319 ind./m2),8月达到次峰值(150 ind./m2),次年1月份达到第三个峰值(76 ind./m2);生物量的三次峰值分别为0.53、0.26、0.20 g/m2.细蜉密度和生物量的周年平均值分别为 91 ind./m2、0.17 g/m2(图 2d).
2.2 生活史
小裳蜉成虫羽化主要发生在8、10月和次年的1、2、3月,随后交配产卵.繁殖发生在4、9月和10、11月,11月出生的个体由于冬天气温较低生长发育缓慢,至次年2、3月开始生长较快.因此,可判断小裳蜉一周年有三个世代:4-8月、7-10月为一个世代,11月至次年3月为一个世代,世代有重叠(图3a).
扁蜉成虫羽化时间为4月和次年2、3月,随后交配产卵,繁殖主要发生在5、6月和8、9、11月,因此亦可划分为三个世代,即5-7月为一个世代、8-10月为一个世代、11月至次年4月为一个世代(图3b).
四节蜉成虫羽化时间为4月、10月和次年2、3月,繁殖主要发生在5、11、12月和次年1、2月,因此可以粗略划分三个世代,即5-8月为一个世代,8-10月,11月至次年4月为一个世代,但世代间存在重叠现象(图3c).
细蜉成虫的羽化主要发生在4月、5月和次年1月,随后交配产卵.繁殖主要发生4、7和12月.由此可粗略划分出两个世代,即6-11月一个世代,12月至次年5月一个世代(图3d).
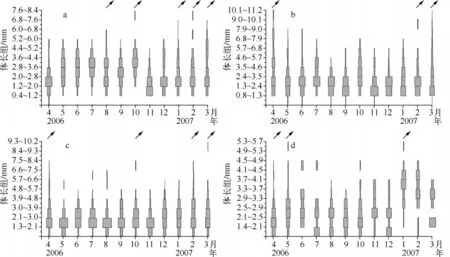
图3 胡家溪小裳蜉(a)、扁蜉(b)、四节蜉(c)和细蜉(d)的种群体长频率分布逐月动态(表示幼虫羽化)Fig.3 Monthly size-frequency distributions of Leptophlebia sp.(a),Electrogena sp.(b),Indobaetis sp.(c)and Caenis sp.(d)in Hujiaxi Stream
2.3 周年生产量
2.3.1 小裳蜉的周年生产量 小裳蜉一年完成三个世代,其周年生产量为其同龄群的3倍.小裳蜉的同龄群生产量为147.14 g/m2,现存量为30.87 g/m2,周年 P/B 系数为14.3.周年生产量湿重为441.42 g/m2,换算成干重[13]为 88.28 g/m2(表 2).
2.3.2 扁蜉的周年生产量 扁蜉一年亦完成三个世代,周年生产量为其同龄群生产量的3倍.扁蜉同龄群生产量为 144.96 g/m2,现存量为 24.77 g/m2,周年 P/B 系数为 17.6;周年生产量湿重为 434.88 g/m2,换算成干重为86.98 g/m2(表3).

表2 胡家溪小裳蜉的周年生产量Tab.2 Annual production of Leptophlebia sp.in Hujiaxi Stream

表3 胡家溪扁蜉的周年生产量Tab.3 Annual production of Electrogena sp.in Hujiaxi Stream
2.3.3 四节蜉的周年生产量 四节蜉一年完成三个世代,周年生产量为其同龄群生产量的3倍.四节蜉同龄群生产量为 249.07 g/m2,现存量为 49.72 g/m2,周年 P/B 系数为 15.03.则周年生产量湿重为747.21 g/m2,换算成干重为149.44 g/m2(表4).
2.3.4 细蜉的周年生产量 细蜉一年完成两个世代,周年生产量为其同龄群生产量的2倍.细蜉同龄群生产量为 20.26 g/m2,现存量为 5.66 g/m2,周年 P/B 系数为 7.20,周年生产量湿重为40.52 g/m2,换算成干重为8.10 g/m2(表5).
2.4 生产量动态
4种蜉蝣优势种小裳蜉、扁蜉、四节蜉、细蜉的生产量动态如图10所示.4种蜉蝣的生产量时间重叠比例相似系数均较高,除细蜉和四节蜉为0.55以外,其余均大于0.60,细蜉和小裳蜉、扁蜉和小裳蜉的相似性系数甚至分别高达0.81和0.79(表6).可见,4种蜉蝣的生产量时间重叠性较高,这可能与4种蜉蝣的生境以及食物资源较为相近有关.

表4 胡家溪四节蜉的周年生产量Tab.4 Annual production of Indobaetis sp.in Hujiaxi Stream

表5 胡家溪细蜉的周年生产量Tab.5 Annual production of Caenis sp.in Hujiaxi Stream
3 讨论
本研究地点位于亚热带地区,4种蜉蝣中,细蜉为一年两代,小裳蜉、扁蜉和四节蜉均为一年三代,与文献报道的亚热带地区蜉蝣一般为一年1~3个世代的规律基本吻合[14-17].
就生产量而言,本研究中的4种蜉蝣平均周年生产量换算成干重分别为:小裳蜉88.28 g/m2、扁蜉86.98g/m2、四节蜉 149.44 g/m2、细蜉 8.10 g/m2;4 种蜉蝣平均周年 P/B 系数分别为 14.3、17.6、15.0、7.2.4种蜉蝣的生产量与已报道的其他河流蜉蝣的生产量相比较为适中[14-27](表 7).但按照 Huryn 等[28]对溪流底栖动物生产量的统计,这些蜉蝣的生产量除细蜉小于100 g/m2,四节蜉的生产量大于100 g/m2,其余两种接近于“高生产量”的范围(100~300 g/m2(DW)),这与胡家溪的总磷含量较高(0.064 mg/L),藻类生长较多、较快,且胡家溪的郁闭度较高,植物碎屑充足有关.另一方面,蜉蝣的捕食者(鱼类、蟹类)相对较少,捕食压力也小.4种蜉蝣的P/B系数与其他河流的相比较为适中,特别与同地区的叹气沟河蜉蝣的P/B系数较为接近,这也说明了在环境条件相似的区域,物种的P/B系数比较稳定.

表6 胡家溪蜉蝣优势种间的相似性系数Tab.6 Proportional similarity between mayfly dominant species in Hujiaxi Stream

表7 不同水体蜉蝣生产力和P/B系数比较Tab.7 Secondary production and P/B ratio of mayfly species
本研究的4种蜉蝣优势种之间比较,二翼蜉的周年生产量最大,这可能与二翼蜉的种群密度、生物量最大有关,且其P/B系数也达到了15.0,仅次于扁蜉的P/B系数17.6.细蜉的周年生产量最小,明显小于其余三种,可能与其种群密度较小有关.就P/B系数而言,扁蜉的最大,这可能与胡家溪扁蜉个体相对较小(75%以上的个体体长在1.3~4.6 mm范围内),保持较快生长速率(周转率)能使得其更加适应胡家溪的生态环境,从而在竞争资源中占据优势有关.小裳蜉有80%以上的个体的体长在2.0~5.2 mm范围内,在胡家溪的几种蜉蝣中个体中相对较大,P/B系数在几种蜉蝣中最小.这与Huryn等报道的溪流水生昆虫P/B系数通常与其个体大小呈反相关关系相一致[28].
从生产量动态来看,4种蜉蝣的生产量除细蜉在2007年3月的日均生产量较低,可能与采样误差有关以外,其余3种蜉蝣在3、4、5月生产量均维持在较高水平(图4),这可能由于越冬的一代蜉蝣在春季的大量繁殖,种群密度较高,故导致其生产量较高.而在8-10月,4种蜉蝣的生产量均较低,这可能与圆顶华溪蟹等捕食者在8-10月的大量繁殖有关.有研究表明四节蜉科Baetidae的幼虫在有捕食者存在时基本不生长,但在没有捕食者存在时,生物量一周就能增加50%[28].4种蜉蝣之间的时间重叠相似性系数除细蜉和四节蜉的为0.55以外,其余均大于0.60,细蜉和小裳蜉、扁蜉和小裳蜉的相似性系数甚至分别高达0.81、0.79.但这仍然在Benke报道的范围之内[21].4种蜉蝣之间缺乏时间维度的分离,可能与这4种蜉蝣在全年均有出现以及它们的多化性、食物资源丰富有关.

图4 胡家溪小裳蜉(a)、扁蜉(b)、四节蜉(c)、细蜉(d)的日均产量及生产量动态Fig.4 Temporal pattern of interval biomass and mean daily production for Leptophlebia sp.(a),Electrogena sp.(b),Indobaetis sp.(c)and Caenis sp.(d)in Hujiaxi Stream
[1]Benke AC,Huryn AD.Benthic invertebrate production-facilitating answers to ecological riddles in freshwater ecosystems.J N Am Benthol Soc,2010,29(1):264-285.
[2]Salas M,Dudgeon D.Life histories,production dynamics and resource utilisation of mayflies(Ephemeroptera)in two tropical Asian forest streams.Freshwater Biol,2003,48:485-499.
[3]Yan Y,Li X.Secondary production and its trophic basis of two species of mayfly in a subtropical stream of China.Int Rev Hydrobiol,2006,91(2):136-147.
[4]Huang Y,Yan Y,Li X.Food web structure of benthic macroinvertebrates in a second order stream of the Hanjiang River basin in middle China.J Freshwater Ecol,2008,23(3):421-427.
[5]Wen F,Jiang J,Deng S et al.Food web and trophic basis of production of the benthic macroinvertebrate community of a subtropical stream(Yangtze River Basin,Middle China).Int Rev Hydrobiol,2009,95(4/5):395-409.
[6]王备新,杨莲芳.我国东部底栖无脊椎动物主要分类单元耐污值.生态学报,2004,24(12):2768-2775.
[7]Georgian T,Wallace JB.Seasonal production dynamics in a guild of periphyton-grazing insects in a southern Appalachian stream.Ecology,1983,64:1236-1248.
[8]Benke AC,Wallace JB.Trophic basis of production among riverine caddisflies:implications for food web analysis.Ecology,1997,78(4):1132-1145.
[9]Hynes HBN,Coleman MJ.A simple method of assessing the annual production of stream benthos.Limnol& Oceanogr,1968,13:569-573.
[10]Hamilton AL.On estimating annual production.Limnol& Oceanogr,1969,14:771-782.
[11]Menzie CA.A note on the Hynes method of estimating secondary production.Limnol& Oceanogr,1980,25(4):770-773.
[12]Whittaker RH.Communities and ecosystems.New York:Macmillian,1975:103.
[13]闫云君.浅水湖泊大型底栖动物生态能量学及生产量的研究[学位论文].武汉:中国科学院水生生物研究所,1998.
[14]Yan Y,Li X.Production and its trophic basis of two mayflies(Leptophlebia sp.and Ephemera sp.)in Heizhuchong Stream in China.J Freshwater Ecol,2006,21(2):301-306.
[15]闫云君,李晓宇.黑竹冲河小裳蜉(Leptophlebia sp.)和蜉蝣(Ephemera sp.)周年生产量.生态科学,2006,25(2):97-101.
[16]Perán A,Velasco J,Millán A.Life cycle and secondary production of Caenis luctuosa(Ephemeroptera)in a semiarid stream(Southeast Spain).Hydrobiologia,1999,400:187-194.
[17]González JM,Basaguren A,Pozo J.Life history and production of Caenis luctuosa(Burmeister)(Ephemeroptera,Caenidae)in two nearby reaches along a small stream.Hydrobiologia,2001,452:209-215.
[18]Christman VD,Voshell JR.Life history,growth and production of Ephemeroptera in experimental ponds.Ann Entomol Soc Am,1992,85:705-712.
[19]Rodgers EB.Production of Caenis(Ephemeroptera:Caenidae)in elevated water temperatures.Freshwat Invertebr Biol,1982,1:2-16.
[20]Benke AC,Jacobi DI.Production dynamics and resource utilization of snag-dwelling mayflies in a blackwater river.Ecology,1994,75:1219-1232.
[21]Welton JS,Ladle M,Bass JAB.Growth and production of five species of Ephemeraptera larvae from an experimental recirculating stream.Freshwater Biol,1982,12:103-122.
[22]Oertli B.Leaf litter processing and energy flow through macroinvertebrates in a woodland pond(Switzerland).Oecologia,1993,96:466-477.
[23]Macfarlane MB,Waters TF.Annual production by caddisflies and mayflies in a western Minnesota plains stream.Can J Fish Aquat Sci,1982,39:1628-1635.
[24]Perán A.Ciclos de vida y producción secundaria en un río de características semiáridas(Río Chícamo,SE de la Peninsula Ibérica):Elcaso de Caenis luctuosa y Sigara scripta[Dissertation].Murcia:Universidad de Murcia,1997.
[25]González JM,Basaguren A,Pozo J.Life history,production and coexistence of two Leptophlebiid mayflies in three sites along a Northern Spain stream.Archiv Hydrobiol,2003,158(3):303-316.
[26]Lobinske RJ,Ali A,Stout IJ.Life history and productivity of Hexagenia limbata(Ephemeroptera:Ephemeridae)and selected physicochemical parameters in two tributaries of the Wekiva river,central Florida.Florida Entomol,1996,79(4):543-551.
[27]苏华武.长江流域二级河流大型底栖动物群落结构和生产量的研究[学位论文].武汉:华中科技大学,2007.
[28]Huryn AD,Wallace JB.Life history and productionof stream insects.Ann Rev Entomol,2000,45:83-110.
