贵州百花湖麦西河河口后生浮游动物群落结构及与环境因子的关系*1
2013-09-25李秋华温远志杨卫诚高廷进
李秋华,温远志,杨卫诚,欧 腾,高廷进,张 垒,魏 刚
(1:贵州师范大学贵州省山地环境信息系统和生态环境保护重点实验室,贵阳550001)
(2:贵州师范大学生命科学学院,贵阳550001)
(3:贵阳学院生态研究中心,贵阳550005)
河口是面源污染和点源污染流入水库较集中的地方,也是水体富营养化程度和水华发生频率较高的地段,河口水质的好坏直接或间接地影响着水库水质状况.改善入库河口的水质是控制外源污染进入水体的最后一道屏障,也是相对容易实施水质改善工程的地段.后生浮游动物作为水域生态系统重要的组成部分,在水域生态系统营养结构中是一类极为重要的消费者,扮演着非常重要的角色,能作为指示生物较好地反映水质状况[1-3].后生浮游动物的群落结构与水体富营养化之间存在着密切关系,富营养化是后生浮游动物群落结构变化的主要动因之一[4-6].通过对后生浮游动物群落结构及与环境因子之间的关系研究,能够反映出该区域的水环境状况.
百花湖(水库)位于贵州省贵阳市西北郊,是红枫湖(水库)的下游湖库,由红枫湖下泄水以及长冲河、东门桥河、麦城河和麦西河四源汇成.麦西河为百花湖四大入库河流之一,也是百花湖主要污染源流入河流之一,因而麦西河河口水质恶化相对严重.近年来,贵州省贵阳市对水资源的保护非常重视,对贵阳市的“两湖一库”即“红枫湖、百花湖和阿哈水库”开展了生态修复工程和水质改善工程.目前,有关贵州省百花湖麦西河河口后生浮游动物群落结构及其与环境因子关系的研究较少,缺乏基础数据.本文通过对贵州百花湖麦西河入库河口后生浮游动物种类分布状况进行2周年的动态调查研究,结合后生浮游动物密度变化、指示种与环境因子之间的关系分析,初步了解百花湖麦西河库湾的后生浮游动物群落结构动态状况及引起动态变化的主要环境因子,为百花湖麦西河库湾的水体修复、水质评价、水生生态系统的健康以及采取相应的管理措施提供一定的基础资料和生物学依据.
1 材料与方法
1.1 采样点和频率
图1 百花湖麦西河口采样点Fig.1 Sampling sites at the estuary of Maixi River in Lake Baihua
百花湖(水库)(26°35'~26°42'N,106°27'~106°34'E)位于贵州高原的中部乌江的一级支流——猫跳河上,在红枫湖的下游,百花湖与红枫湖合称贵阳市“两湖”,是贵阳市重要饮用水源地.麦西河是百花湖主要入库河流之一.在麦西河河口设有2个采样点,采样点S1位于麦西河的支流上,而采样点S2在河口与水库开阔处(图1).S1与S2采样点相差2 km左右.于2009年8月至2011年8月,每月1次、每月2次或每两月1次的频率在S1和S2点进行采样.
1.2 后生浮游动物的鉴定与计数
后生浮游动物按照全国内陆水域水生生物和环境调查规范中的方法进行[7-10].定量样品:取20 L水样置于25#浮游生物网中过滤,浓缩至50 ml,加福尔马林溶液固定后存于100 ml标本瓶中,带回实验室镜检.各类后生浮游动物种类按照文献[7-10]鉴定.透明度(SD)和pH分别采用透明度盘(即塞氏盘)和pHS-3C型酸度计测定.根据《水和废水监测分析方法》[11]测定总氮(TN)、总磷(TP)、硝态氮(-N)等水化指标.叶绿素a(Chl.a)浓度测定:取200~500 ml样品过0.45 μm的纤维滤膜抽滤,反复冻融后用丙酮萃取法进行测定[12].
2 结果与分析
2.1 轮虫的种类组成与动态变化
调查期间共鉴定出轮虫29种,其中S1、S2样点分别鉴定出28、29种,常见的检出种有螺形龟甲轮虫、热带龟甲轮虫、萼花臂尾轮、裂足臂尾轮虫、角突臂尾轮虫、前节晶囊轮虫、卜氏晶囊轮虫、长三肢轮虫、长肢多肢轮虫(表1).S1、S2样点的轮虫种类组成具有较大的相似度.在调查期间鉴定出的29种轮虫中,臂尾轮科出现的种类最多,其次是异尾轮科;从种类的季节变化来看,均为冬季种类较少,春、夏季种类较多;从种类组成上看,大部分都属于污染种,主要属于β中污染型指示种类.

表1 百花湖麦西河入库河口轮虫的种类组成Tab.1 Composition of rotifer at the estuary of Maixi River in Lake Baihua
百花湖麦西河入库河口轮虫的丰度在0.16~837.80 ind./L之间,最大值和最小值均在S2采样点中,分别出现在2010年11月和7月(图1).通过独立样本的t检验,S1和S2采样点轮虫的季节动态变化无明显差异(n=27,P>0.05).从轮虫的季节动态看,在2010年8月份之后,轮虫丰度明显高于2009年同期轮虫丰度,每年的3-4月份轮虫的丰度有一个高峰值,但是轮虫的最高值出现在11月份.
2.2 桡足类的种类组成及动态变化
百花湖麦西河入库河口桡足类浮游动物种类相对较少,仅检测到了温中剑水蚤(Mesocyclops thermocyclopoides)和猛水蚤(Harpacticella sp.),以无节幼体(Nauplius)和桡足幼体(Copepodites)为主.百花湖麦西河入库河口桡足类的丰度在0~642.75 ind./L之间,最大值和最小值均在S2采样点中,分别出现在2010年5月和7月(图2).通过独立样本的t检验,S1和S2采样点桡足类的季节动态变化无显著差异(n=27,P>0.05).从桡足类的季节动态可以看出,每年的3-4月份桡足类的丰度有一个高峰值.
2.3 枝角类的种类组成及动态变化
调查期间共监测到枝角类浮游动物8种,包括长额象鼻溞(Bosmina longirostris)、短腹锐额溞(Alonella exigua)、短尾秀体溞(Diaphanosoma brachyurum)、短腹平直溞(Pleuroxus aduncus)、微型裸腹溞(Moina micrura)、蚤状溞(Daphnia pulex Leydig)、僧帽溞(Daphnia cucullata)、奥氏秀体溞(Diaphanosoma orghidani)等.其中,长额象鼻溞和奥氏秀体溞为麦西河库湾常见种类,而以长额象鼻溞为主要优势种类.枝角类丰度在0~31.20 ind./L之间,主要出现在11月至次年4月份.
2.4 后生浮游动物丰度组成
后生浮游动物的丰度主要由轮虫和桡足类组成,枝角类丰度的比例相对较少,轮虫丰度比例在4.09%~100%之间,桡足类丰度比例在0~95.64%之间,轮虫丰度比例最高出现在2010年7月的S2采样点,而桡足类最高值出现在2010年3月的S2采样点(图3).对比S1和S2采样点可以看出,S1采样点轮虫丰度比例高于S2采样点;从季节来看,2009年冬季桡足类丰度比例明显高于其他时段,但是在2010年之后,轮虫丰度一直维持较高的比例.
2.5 优势后生浮游动物动态变化特征
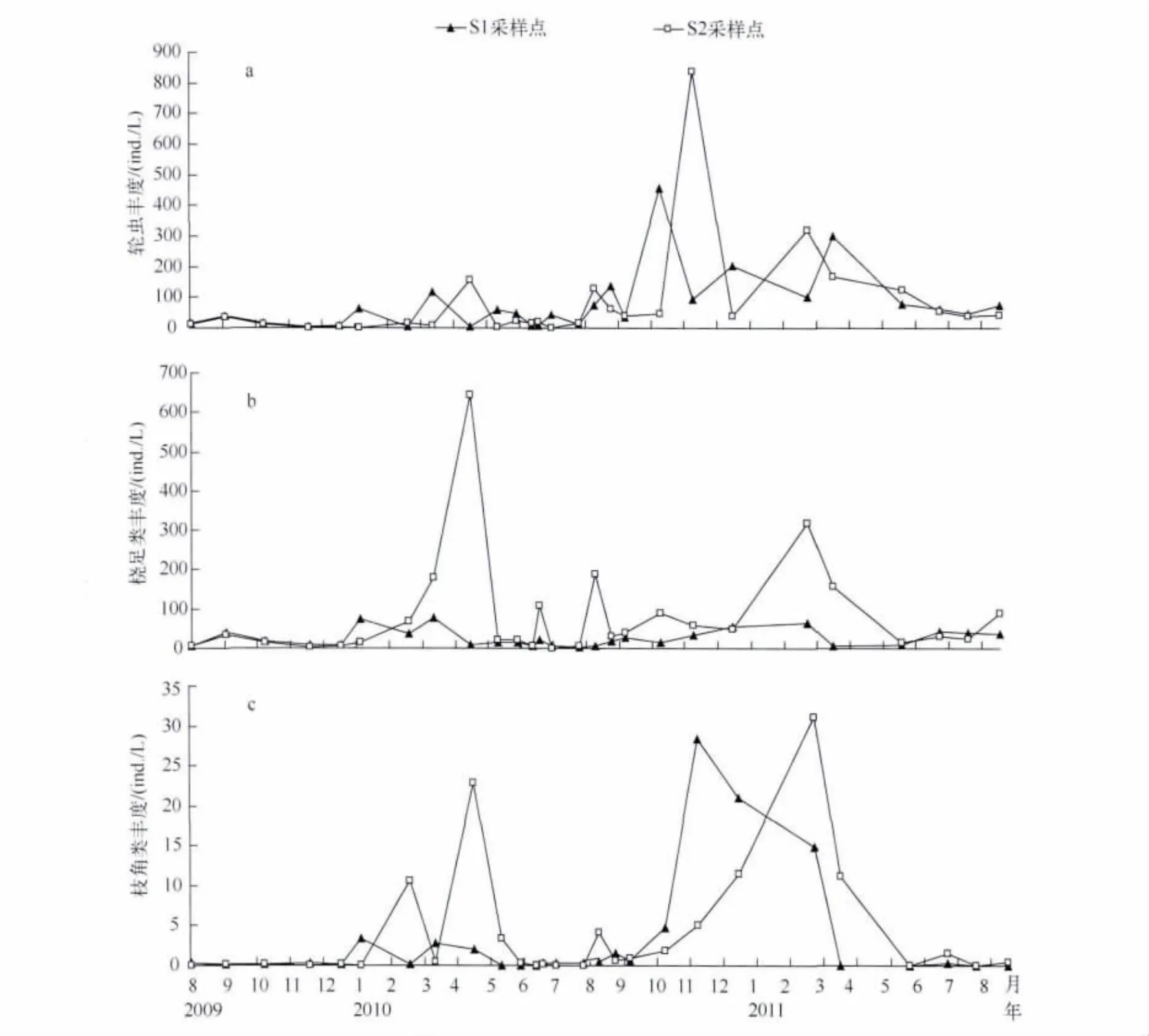
图2 百花湖麦西河入库河口轮虫(a)、桡足类(b)和枝角类(c)丰度动态变化Fig.2 Dynamics of rotifer(a),copepod(b)and cladocera(c)abundance at the estuary of Maixi River in Lake Baihua
S1和S2采样点主要优势后生浮游动物的季节动态变化表明,萼花臂尾轮虫、前节晶囊轮虫、卜氏晶囊轮虫和裂足臂尾轮虫为轮虫类的主要优势种,而长额象鼻溞为枝角类的主要优势种,无节幼体为桡足类的主要优势种.前节晶囊轮虫、卜氏晶囊轮虫在10-11月和5-6月为主要优势种,其最高丰度出现在10-11月,分别达到424.88 ind./L 和392.20 ind./L.裂足臂尾轮虫在8-9月为主要优势种,达到61.99 ind./L.长额象鼻溞为冬、春季主要优势种,最高丰度达到27.93 ind./L;无节幼体在12月至次年2月份占优势,最高丰度达到67.68 ind./L(图4).
2.6 后生浮游动物与环境因子之间的关系
2.6.1 主要环境指标 百花湖麦西河入库河口透明度在0.50~2.60 m之间变化,最大值出现在2010年12月S2采样点,最小值出现在2010年5月S1采样点.S1和S2采样点透明度相差不大,S1采样点平均透明度略低于S2采样点(图5).从季节变化来看,透明度在冬季最高、夏季最低.叶绿素a浓度在9.00~100.58 μg/L之间变化,最大值出现在2009年10月S1采样点,而最小值出现在2010年12月S2采样点,S2采样点叶绿素a浓度明显低于S1采样点.叶绿素a浓度季节变化明显,冬季最低、夏季(8-10月)最高.总氮浓度在0.73~2.85 mg/L之间变化,最小值和最大值都出现在S2采样点,总氮的季节变化明显,冬季较高、夏季较低;总磷浓度在0.03~0.12 mg/L之间变化,其季节变化不如总氮明显,但总磷浓度有逐年降低的趋势,2009年总磷浓度最高(图5).从富营养化参数可以看出,麦西河河口富营养化程度较高,大部分时期处于富营养化状态或中 富营养状态.百花湖麦西河河口pH值在7.15~9.15之间,反映出了高pH值的特征;采样期间温度在3.0~25.3℃之间,具有明显的季节性;降雨量在3~180 mm之间,主要集中在4-10月,而冬季12月至次年3月降雨相对较少.

图3 百花湖麦西河入库河口后生浮游动物丰度组成Fig.3 Percents of metazooplankton abundance at the estuary of Maixi River in Lake Baihua
2.6.2 后生浮游动物与环境因子之间的关系 选择了13个对后生浮游动物群落结构动态变化影响较大的环境因子,包括水体的透明度、叶绿素a、总氮、硝态氮、亚硝态氮、氨态氮、总磷、磷酸根、悬浮物、浊度、pH、温度和降雨量进行相关分析,表2仅列出了与后生浮游动物丰度相关性较好的环境因子.温度、透明度、总氮和硝态氮与轮虫和枝角类显著相关,桡足类与这些环境因子相关性不显著(表2).其中,轮虫丰度与透明度、总氮、硝态氮(rSD=0.487,rTN=0.419,rNO--N=0.501,P <0.01;n=50)呈极显著正相关,而与浊度、温度(r浊度=3-0.345,r温度= -0.332,P <0.05;n=50)呈显著负相关.枝角类丰度与透明度、硝态氮(rSD=0.528,rNO--N=30.409,P <0.01;n=50)呈极显著正相关,与温度(r温度= -0.427,P <0.01;n=50)呈极显著负相关.
3 讨论
3.1 后生浮游动物群落结构
百花湖麦西河河口后生浮游动物群落结构特点为轮虫的种类和丰度在后生浮游动物中所占的比例较高,大型浮游动物(枝角类、桡足类)种类和丰度所占的比例较少,这与许多的湖泊和水库浮游动物各类群的组成比例情况相似[13-16].百花湖麦西河河口富营养化程度较高,大部分时期处于富营养化状态,尤其以夏、秋季富营养化更为严重[17].轮虫作为水生生态系统的重要组成部分,其种类组成及丰度消长与水质密切相关.通常情况下,在清洁型水体中轮虫呈现出种类多、数量少的特点,而在中污染型的水体中,往往是耐污染类轮虫为优势种(往往以较高数量出现)[14-16].百花湖麦西河河口在监测期内共监测到29种轮虫,以螺形龟甲轮虫、热带龟甲轮虫、萼花臂尾轮、裂足臂尾轮虫、角突臂轮虫、前节晶囊轮虫、卜氏晶囊轮虫、长三肢轮虫、多肢轮虫常见,这些种类都是富营养化指示种,后生浮游动物种类的组成反映出百花湖麦西河河口的水质状况,轮虫指示出了河口富营养化程度较高.在调查中,枝角类和桡足类的种类较少,季节变化明显,优势种类明显,枝角类主要以长额象鼻溞为优势种,在冬、春季更明显.

图4 百花湖麦西河入库河口优势后生浮游动物丰度动态变化Fig.4 Dynamics of dominant metazooplankton species abundance at the estuary of Maixi River in Lake Baihua
3.2 主要理化因子对后生浮游动物群落结构的影响
河流作为地球陆地表层生态系统的脉络,联结着陆地生态系统与湖泊.一方面,河流接纳了来自陆地生态系统的各种营养组分,其中碳组分包括溶解有机碳(DOC)、颗粒有机碳(POC)、溶解无机碳(DIC)以及颗粒无机碳(PIC),其中DIC包括、、H2CO3、CO2等组分,导致水体有机质积累.另一方面,在河流内部水生生物光合作用与呼吸作用、有机质的氧化分解和转化利用、碳酸盐的溶解和沉淀等地球化学过程,导致水体以河流DIC为核心发生一系列的水环境变异,对水体富营养化产生重要影响[18].河口由于人口密集,大量工业废水和生活污水的排放使河口水域生态环境发生了一定程度的改变,河口水域水文环境复杂多变,其浮游动物动态在很大程度与水动力学有关[18-19],但不同水体具有不同的影响因子.百花湖麦西河河口具有典型的喀斯特地貌特征.在喀斯特地区,以碳酸盐岩化学溶蚀为特色的喀斯特流域河流DIC浓度较非喀斯特流域高出好几倍.在高钙、高碱度的喀斯特水体中,由于溶解二氧化碳的施肥效益,水生生物利用光合作用合成有机质的效率大大增加,沉水植物与浮游植物迅速繁殖,较易导致富营养进程速度加快[18].轮虫主要以细菌等为食,与S2采样点相比,S1采样点要远离河口开阔区,S1采样点轮虫丰度较高也指示了该点富营养化程度要高于S2采样点.温度被认为是影响浮游动物发生并引起其种类季节演替的关键因子之一,水温的变化可导致被捕食的压力和浮游动物的食物发生改变,进而直接或间接地改变浮游动物的群落结构[1,15].本研究得出,在百花湖麦西河河口,温度、透明度、总氮和硝态氮与轮虫和枝角类丰度呈显著相关,温度是影响轮虫和枝角类丰度的主要环境因子,这也与许多研究结果一致[1,5].在对百花湖麦西河河口浮游植物群落结构动态变化研究中,温度也是影响该区域浮游植物群落结构动态变化的主要环境因子[17],温度的季节变化引起了该区域的水环境状态发生改变,同时也引起了浮游动物群落结构的动态变化.

图5 百花湖麦西河入库河口透明度和叶绿素a浓度(a),总氮和总磷浓度(b),pH值、温度和降雨量(c)的动态变化Fig.5 Dynamics of secchi depth and concentration of chlorophyll-a(a);concentration of total nitrogen and total phosphorus(b);pH,temperature and rainfall(c)at the estuary of Maixi River in Lake Baihua

表2 后生浮游动物丰度与环境因子之间的关系Tab.2 The correlation between metazooplankton abundance and environmental factors
3.3 水生生物对环境因子的响应
在对麦西河河口浮游植物与环境因子之间的关系研究中得出温度是影响浮游植物群落结构的主要环境因子,营养盐浓度成为影响浮游植物群落结构动态变化的次要环境因子[17],与本文对后生浮游动物与环境因子关系研究的结果一致.这是因为:一方面,贵州高原地区季节变化明显,温度年际变化显著,温度是影响浮游生物动态变化的主要因子;另一方面,麦西河河口富营养状态较高,营养盐已经能够满足浮游生物生长的需求,相对高的营养盐浓度变化对浮游生物群落结构动态的变化影响不大,营养盐不是影响浮游生物群落结构变化的主要环境因子.因此,在贫营养水体,营养盐是影响浮游生物群落结构动态变化的主要环境因子,而在富营养化程度较高水体,水文因素的动态变化是影响浮游生物群落结构动态变化的主要环境因子[20].
[1]王晓辉,望 甜,韩博平.南亚热带富营养化抽水型水库轮虫的组成与动态.湖泊科学,2009,21(1):101-109.
[2]Chen FZ,Xie P.The effects of fresh and decomposed Microcystis aeruginosa on cladocerans from a subtropic Chinese lake.Journal of Freshwater Ecology,2003,18:97-104.
[3]Lin QQ,Duan SS,Hu R et al.Zooplankton distribution in tropical reservoirs,South China.Internat Rev Hydrobiol,2003,88(6):602-613.
[4]Guevara G,Lozano P,Reinoso G et al.Horizontal and seasonal patterns of tropical zooplankton from the eutrophic Prado Reservoir(Colombia).Limnologica,2009,39(2):128-139.
[5]郭 凯,赵 文,殷守仁等.北京官厅水库轮虫群落结构与水体富营养化状况.湖泊科学,2010,22(2):256-264.
[6]温新利,席贻龙,张 雷等.芜湖市镜湖轮虫群落结构分析及水质的生态学评价.水生生物学报,2006,30(2):153-158.
[7]周凤霞,陈剑虹.淡水微型生物图谱.北京:化学工业出版社,2005.
[8]黄祥飞.湖泊生态调查观测与分析.北京:中国标准出版社,2000.
[9]章宗涉,黄祥飞.淡水浮游生物研究方法.北京:科学出版社,1991.
[10]王家楫.中国淡水轮虫志.北京:科学出版社,1961:21-283.
[11]国家环境保护总局《水和废水监测分析方法》编委会.水和废水监测分析方法:第4版.北京:中国环境科学出版社,2002.
[12]林少君,贺立静,黄沛生等.浮游植物中叶绿素a提取方法的比较与改进.生态科学,2005,24(1):9-11.
[13]潘继征,熊 飞,李文朝等.抚仙湖浮游甲壳动物群落结构与空间分布.湖泊科学,2009,21(3):408-414.
[14]任晶晶,林秋奇,韩博平.热带水库敞水区轮虫种类组成与数量结构——基于海南7座典型水库的分析.湖泊科学,2010,22(2):272-280.
[15]李慧明,陈 花,韩小玉.海南热带水库敞水区浮游甲壳动物群落结构特征.湖泊科学,2011,23(2):251-256.
[16]杨桂军,秦伯强,高 光等.太湖不同湖区轮虫群落结构季节变化的比较研究.环境科学,2008,29(10):2963-2969.
[17]李秋华,陈丽丽,夏品华等.贵州百花湖麦西河河口浮游植物群落结构及其与环境因子之间关系.湖泊科学,2011,23(4):612-618.
[18]刘丛强.生物地球化学过程与地表物质循环——西南喀斯特流域侵蚀与生源要素循环.北京:科学出版社,2007.
[19]李秋华,夏品华,吴 红等.用物理-生态集成技术局部控制富营养化.环境科学,2012,33(7):2303-2308.
[20]Reynolds CS.The ecology of freshwater phytoplankton.Cambridge:Cambridge University Press,2006.
