Cry1Ab 蛋白在蜘蛛体内的富集及对蜘蛛相关酶活性的影响
2013-09-25魏宝阳李超民罗翔宇肖安琪
周 颖,魏宝阳,王 智,李超民,罗翔宇,肖安琪
(湖南农业大学生物科学技术学院,湖南 长沙410128)
自1996年,全球第一例转基因作物在美国开始商品化种植生产以来,转基因作物就开始大面积推广,到2011年全世界转基因作物种植面积由2010年的14 800 万hm2增加8.1%达到16 000 万hm2[1]。2009年我国农业部批准了2 个抗虫转基因水稻品系的安全证书后[2-4],Bt 作物更是得到广泛的关注。Harwood 等[5]研究表明,转Bt 基因玉米中的Cry1Ab 蛋白可通过食物链的传递转移到玉米地中的相关天敌玉米跳甲、日本金龟子等动物的体内。姜永厚等[6]研究表明,取食转Bt 水稻的褐飞虱(Nilaparvata lugens)、稻蚜(Sitobion avenae)以及用取食转Bt 水稻的二化螟或稻眼蝶幼虫喂食的拟水狼蛛(Pirata subpiraticus)体内都能检测出Cry1Ab 蛋白,这些研究都表明Bt 蛋白能够随着食物链进行传递。但是,Bt 蛋白随着食物链进行传递过程中在蜘蛛体内的富集情况、时间效应以及对蜘蛛体内的相关的酶活性的影响还有待研究。以拟环纹豹蛛为主要试验材料,研究了Cry1Ab 蛋白在蜘蛛体内的富集及对蜘蛛相关酶活性的影响,以期为转Bt 水稻安全性评价提供参考依据。
1 材料和方法
1.1 材 料
1.1.1 供试水稻 转Cry1Ab 基因水稻汕优63(简称Bt 水稻)与对照品种亲本汕优63 均由湖南师范大学提供,Bt 水稻和对照品种在湖南农业大学实验室内盆栽进行培养。水稻在生长期内不喷洒任何农药,其他管理均为常规管理。
1.1.2 供试稻飞虱 以湖南省农科院水稻所试验田捕捉的褐飞虱为材料,在实验室盆栽水稻中进行培养,对照组取食对照水稻,试验组取食Bt 水稻。
1.1.3 供试蜘蛛 拟环纹豹蛛采集于湖南农业科学院水稻所试验田,采回后分装于试管内,饥饿48 h 后分别以取食10~15 d 的对照组(即普通汕优63水稻)和试验组(即Bt 水稻)盆栽水稻的褐飞虱进行饲养,提供充足的稻飞虱和水,在温度25℃、湿度70%的人工气候箱内进行喂养。喂食褐飞虱5、10、15 d 后的蜘蛛在冰水中用匀浆器进行匀浆后,4 000 r/min 离心,取上清液作为酶源。
1.2 方 法
1.2.1 试验设计 试验设2 个处理,蜘蛛饥饿48 h 后,试验组以取食10~15 d Bt 水稻的褐飞虱饲养,对照组以取食10~15 d 普通汕优63 水稻的褐飞虱饲养。试验期间提供充足的稻飞虱和水,在温度25℃、湿度70%的人工气候箱内进行喂养。喂食褐飞虱5、10、15 d 后的蜘蛛在冰水中用匀浆器进行匀浆后,4 000 r/min 离心,取上清液作为酶源。
1.2.2 数据测定 (1)蜘蛛体内可溶性蛋白的测定:可溶性蛋白采用考马斯亮G-250 法[7],以牛血清蛋白做蛋白制备标准曲线。(2)酶活力的测定:乙酰胆碱酯酶(AchE)活力测定参照高希武[8]的方法,超氧化物歧化酶(SOD)活力和谷胱甘肽过氧化物酶(GSH-Px)活力用南京建成公司生产的相关试剂盒进行测定,测定严格按照试剂盒说明操作。(3)Bt蛋白含量的测定:Bt蛋白含量测定采用由美国Agdia 公司提供的Bt-Cry1Ab/Ac ELISA 平板试剂盒测定。
1.2.3 数据分析 所有数据均用SPSS 17.0 软件进行统计分析,用Excel 2003 作图。
2 结果与分析
2.1 稻飞虱和蜘蛛体内Cry1Ab 蛋白含量的变化
由表1 可知,取食Bt 水稻后,在0~5 d 内,褐飞虱体内的Bt 蛋白含量很低,几乎没有检测到,在5~15 d 内,褐飞虱体内的Bt蛋白含量逐渐上升,在第15 天时到达最高。故研究采用取食Bt 水稻10~15 d 内的褐飞虱喂食蜘蛛。
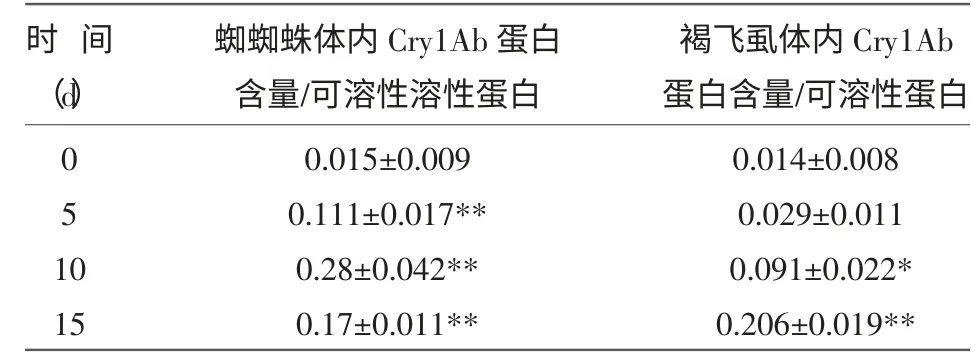
表1 褐飞虱、拟环纹豹蛛体内Cry1Ab 蛋白含量
喂食取食Bt蛋白10 d 的稻飞虱后,在5~15 d内,蜘蛛体内均检测到Bt蛋白含量,在第10 天Bt蛋白含量达到最高,10 d 后呈逐渐下降趋势,但在第15 天时,蜘蛛体内的Bt 蛋白含量仍高于第5 天时的水平。蜘蛛体内Bt蛋白的最高含量高于稻飞虱体内Bt蛋白的最高含量,说明Bt蛋白沿“Bt 水稻—稻飞虱—蜘蛛”这一条食物链传递时具有生物富集效应。
2.2 蜘蛛体内相关酶活力的变化
2.2.1 AchE 活力的变化 如表2 所示,在喂食含有Bt蛋白的稻飞虱5 d 后,试验组拟环纹豹蛛体内的AchE 活力较对照组的出现了略微升高;在喂食10 d 后,当拟环纹豹蛛体内的Bt蛋白含量达到最高值时,试验组拟环纹豹蛛体内的AchE 低于对照组;在喂食15 d 时,试验组拟环纹豹蛛体内的AchE 活力高于对照组,但没有达到显著性差异(P>0.05)。由此推测,AchE 活力的变化可能与Bt蛋白的含量存在负相关性。
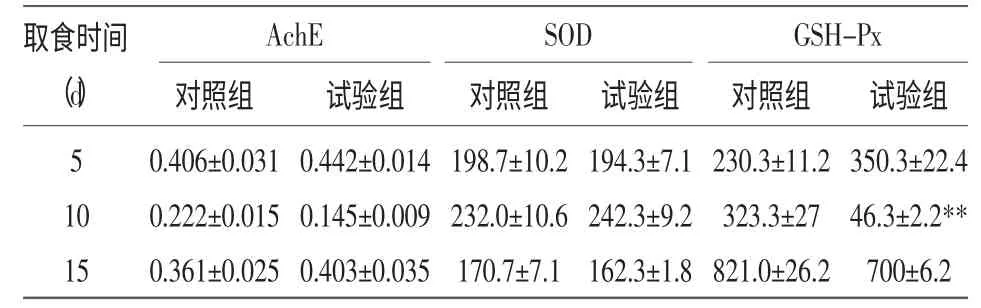
表2 Cry1Ab 蛋白富集作用对拟环纹豹蛛体内酶活力的影响 (U/mg)
2.2.2 SOD 活力的变化 如表2 所示,在喂食含有Bt 蛋白的褐飞虱5 d 后,试验组拟环纹豹蛛体内的SOD 活力较对照组的略低,可能是Bt蛋白的富集对蜘蛛体内的酶活力出现了抑制作用;在喂食10 d 后,拟环纹豹蛛体内的Bt蛋白含量处于最高值,试验组拟环纹豹蛛体内的SOD 活力高于对照组,由于Bt蛋白的富集,造成蜘蛛体内自由基含量较多,SOD 对其进行消除;在喂食15 d 时,试验组拟环纹豹蛛体内的SOD 活力均低于对照组,可能是由于Bt蛋白的富集造成整个拟环纹豹蛛体内的SOD 活性无法调控,从而抑制酶活力。但是,在整个试验期间试验组的拟环纹豹蛛体内的SOD 活力与对照组相比,均没有达到显著性差异(P>0.05)。
2.2.3 GSH-Px 活力的变化 如表2 所示,在喂食含有Bt蛋白的稻飞虱5 d 后,试验组拟环纹豹蛛体内的GSH-Px 活力略高于对照组;但在喂食10 d后,拟环纹豹蛛体内的Bt蛋白含量达到最高值时,试验组拟环纹豹蛛体内的GSH-Px 活力低于对照组,且存在极显著性差异(P<0.01)。这可能是由于蜘蛛体内的保护机制开始能够消除Bt蛋白对拟环纹豹蛛体内酶活力的影响,但由于Bt蛋白的大量富集,超出了保护机制的范围,导致酶活力受到抑制。在喂食15 d 后,试验组拟环纹豹蛛体内的GSH-Px活力仍然低于对照组,说明GSH-Px 活力的恢复需要一定的时间。由此推测,GSH-Px 活力的变化可能与Bt 蛋白的含量存在负相关性。
3 讨 论
该研究中Bt 蛋白能够随着“Bt 水稻—稻飞虱—蜘蛛(拟环纹豹蛛)”食物链进行传递,并具有生物放大效应;拟环纹豹蛛体内的Bt 蛋白含量随着时间变化而变化,当Bt 蛋白增长到一定的程度后,逐渐出现下降的趋势,这和陈茂等[9]研究结果一致,可能与蜘蛛体内存在相关保护机制有关,其具体机理还有待进一步研究。
Bt 蛋白在蜘蛛体内的富集造成蜘蛛体内相关酶活力的变化。喂食5 d 时,Bt 蛋白含量开始在体内积累,蜘蛛体内相关的保护酶,主要是GSH-Px活力升高,来抵御Bt 蛋白的影响,同时蜘蛛体内的AchE 活力也升高,通过在神经方面的某些调控作用来对Bt 蛋白活力起到防御作用;而SOD 活力下降,可能是由于Bt蛋白对其产生抑制作用所致。随着Bt 蛋白的持续积累,在喂食10 d 后,蜘蛛体内的AchE 和GSH-Px 活力出现最低值,其中GSHPx 活力与对照组差异达极显著水平,可能是因为Bt 蛋白的持续作用超出酶活力能够调控的范围,使酶活力反而受到抑制;而SOD 活力在此时升高,可能是由于大量Bt 蛋白积累,激活了其体内防御机制所致。之后,蜘蛛体内的防御机制导致Bt 蛋白含量下降,GSH-Px 和AchE 活力开始不断恢复,在喂食15 d 时均还略高于对照组。
在该研究中,酶活力的变化情况和张志罡等[10]报道的Cry1Ab 蛋白对稻纵卷叶螟幼虫体内AchE活力先迅速上升后有剧烈下降趋势,以及SOD 先稳定平缓后急剧升高又迅速下降趋势不一致;和郭文娟等[11]报道的基因水稻对大螟幼虫体内3 种保护酶的变化情况以及张雁等[12]报道的转Bt 基因南林895 杨对杨扇舟蛾体内酶的变化情况均不相同,这可能是由于Bt 蛋白对靶标生物和非靶标生物的作用机理不一致所造成的。
[1]孙雷心.国际农业生物技术应用服务组织(ISAAA)发布转基因作物年度发展报告[J].中国农业科学,2012,45(4):616.
[2]李黎红,叶卫军,郭龙彪.我国转基因水稻研究进展和商业化前景分析[J].中国稻米,2012,18(6):1-4.
[3]蒋建科. 转基因水稻能不能放心吃[N].人民日报,2010-02-01.
[4]王 佳.200 亿元财政搅动转基因水稻商业“神经”[N].中国经营报,2010-01-17.
[5]Harwood J K,Wallin W G.,Obrycki J J.Uptake of Bt endotoxins by nontarget herbivores and highter arthropod predators:molecular evidence from a transgenic corn agroecosystem[J].Mol.Ecol,2005,14(9):2815-2823.
[6]姜永厚,付 强,程家安.转Bt 基因水稻表达的毒蛋白Cry1Ab在害虫及其捕食者体内的积累动态[J].昆虫学报,2004,47(4):454-460.
[7]Bradford M M.A rapid and sensitive method for the quantitation of microgam quantities of protein utilizing the principle proteindye binding[J].Anal.Biochem,1976,72:248-254.
[8]高希武.Gorun 等改进的Ellman 胆碱酯酶活性测定方法介绍[J].昆虫知识,1987,24(4):425-246.
[9]陈 茂,叶恭银,卢新民.Cry1Ab 杀虫蛋白在水稻—褐飞虱—拟环纹豹蛛食物链中转移与富集[J].昆虫学报,2005,48(2):208-213.
[10]张志罡,孙继英,李 勇.Cry1Ab 蛋白对稻纵卷叶螟幼虫体内三种保护酶活性的影响[J].生物技术通报,2011,(1):148-152.
[11]郭文娟,陆驰宇,熊应强.转cry1Ab/cry1Ac 基因水稻对大螟幼虫体内三种保护酶活性的影响[J].昆虫学报,2012,55(8):958-963.
[12]张 雁,郭同斌,诸葛强.转Bt 基因南林895 杨对杨扇舟蛾体内酶的影响[J].林业科学,2012,48(6):95-99.
