同一离体培养下梓树属不同种的反应差异1)
2013-09-18于永明
于永明
(甘肃省小陇山林业科学研究所,天水,741022)
王军辉 麻文俊
(国家林业局林木培育重点实验室(中国林业科学研究院林业研究所))
马建伟 张宋智 韩云花 李平英
(甘肃省小陇山林业科学研究所)
灰楸(Catalpa fargesii Bureau)、滇楸(Catalpa fargesii f.duclouxii Dode)、梓树(Catalpa ovata G.Don)、黄金树(Catalpa speciosa(Barney)Engelm)、楸树(Catalpa bungei C.A.Meyer)为紫葳科梓树属(Catalpa scop)较有代表的5个种,目前,国内外对楸树各方面已展开研究,并取得一定成果,关于梓树属方面做组培研究的文献还未见报道,尤其是属内不同种间的对比性试验[1-7]。“十一五”期间,珍贵阔叶树种种质创新课题组对梓树属不同种源和无性系选择、目标树选择和培育、立地选择、干形培育、混交林结构调整、密度控制和水肥调控等关键技术进行了深入研究,建立珍贵用材林定向单株培育的科技创新平台,提高了我国珍贵用材林培育的技术创新能力。在梓树属组培研究领域,国内对楸树研究报道较多,各研究配方与针对种源有所不同,普遍应用楸树嫩枝为外植体,有部分研究人员对楸树体细胞胚胎发生进行研究并取得一定的研究结论[8-9]。珍贵阔叶树种种质创新课题组对梓树属组培技术进行了深入研究,已成功对优良无性系大批量生产,为规范优良无性系组培技术并便于生产,制定了《楸树无性系组培快繁技术规程》地方标准[10],为良种的推广应用及开发提供了重要的理论价值和实践意义。为进一步选育特性良种,应用梓树属遗传转化体系,有必要对其组织培养特性进行研究。本试验对典型的5个种进行组织培养比较研究,以探讨梓树属不同种再生芽增殖和生根培养遗传因素及外界条件影响的大小,通过不同种间离体植株再生能力及性状表型间的差异,揭示其诱导表现规律和特点,为建立不同种快繁及遗传转化体系提供理论依据。
1 材料与方法
实验材料为珍贵阔叶树种种质创新课题组收集的5个种:灰楸(河南洛阳)、滇楸(贵州毕节)、黄金树(辽宁恒仁)、梓树(辽宁开原)、楸树(河南洛阳),在中国林业科学院林业研究所培育这些种的组培苗。
1.1 增殖与生根培养
选择生长状况一致的无菌组培苗,转接增殖分化培养基和生根培养基上。每个种接种10瓶,每瓶接种3个分生株,设置3个重复。增殖培养茎段为带1个腋芽的茎段,确保诱导分化在同一水平;生根茎段长度为1.5 cm,确保插入培养基茎段部位没有腋芽,以免影响根系的诱导。
继代增殖分化培养基为 DKW+1.0 mg·L-16-BA+0.2 mg·L-1IBA+25 g·L-1蔗糖+4.5 g·L-1琼脂(pH值5.8);生根培养基为 1/2 MS+0.1 mg·L-1IBA+0.01 mg·L-1NAA+10 g·L-1蔗糖+5.0 g·L-1琼脂(pH值5.8)。组培苗培养温度为22~26℃,光照强度为2000 lx,光照时间为14 h·d-1。
1.2 移栽炼苗
基质配比为V(泥炭土)∶V(珍珠岩)=4∶1,基质应于炼苗前提前配好并装入营养钵,营养钵一般采用规格为8 cm×8 cm,配基质时适量加入多菌灵。炼苗时将生根瓶苗置于温室,适应环境7 d左右松开封瓶绳,隔天将瓶膜松开,10 d左右移栽,移栽后及时覆膜,遮阴,每天喷水3次。
1.3 数据统计
增殖培养10、25、35 d观测统计,统计指标:增殖芽数、芽长、叶数、接入茎段基部愈伤组织横向及纵向膨大情况。生根培养30 d后观测统计,统计指标:接入茎段发根数、根长、发芽数、芽长、叶数。依据芽数与芽长计算增殖系数。数据统计分析采用SPSS14.0。
2 结果与分析
2.1 梓树属不同种不定芽增殖差异
芽的继代增殖,种的不同增殖效果有着明显差异,这说明不同种在芽增殖生长上对培养基具有选择性。方差分析表明,增殖分化指标增殖系数(P=0.055)、增殖芽数(P=0.0005)、芽长(P=0)、叶数(P=0)、接入茎段基部愈伤组织横向膨大(P=0.043)和纵向膨大(P=0)在不同种间达到显著水平。从方差分量看,种的差异是影响各指标分量的主要因素,各指标种的方差分量在78.491% ~97.647%。增殖系数种方差分量为77.014%,这说明梓树属不同种瓶苗生长受较强的遗传因素制约。
2.2 继代增殖培养多重比较及方差分量动态变化
各指标不同培养时间方差分量(表1)显示,增殖系数随着培养时间的递增,种间方差分量在逐渐减小;培养7~15 d,增殖分化芽数、增殖芽芽长随培养时间递增,种间方差分量在逐渐增大,叶数的分量在逐渐减小;15~25 d,芽数的分化与增殖芽的生长、茎段基部愈伤组织纵向膨大种间方差分量在逐渐减小,叶数、茎段基部愈伤组织横向膨大却在增大;培养25~35 d,芽的生长、茎段基部愈伤组织横向膨大种间方差分量在逐渐减小,增殖芽的分化、叶数、愈伤组织纵向膨大却在增大。结果说明,在培养过程中,种的差异是各指标生长变化的主要因素,在不同的培养时间段影响程度不同。

表1 不同培养时间继代培养各指标分量变化
表2显示,在同一培养条件下,培养35 d后,滇楸增殖分化芽数最多为4.35个,种间由多到少为滇楸>灰楸>楸树>梓树>黄金树;黄金树增殖分化芽生长最快,为3.03 cm,由快到慢为黄金树>梓树>滇楸>灰楸>楸树;增殖系数滇楸与灰楸、楸树差异显著,种间由高到低为滇楸>梓树>黄金树>灰楸>楸树;叶数滇楸与其余4个种差异显著,相对滇楸分化叶数最少为5.29个,梓树最多为7.33;茎段基部愈伤组织横向与纵向膨大的种间差异显著,以灰楸膨大面积最大,为0.80 cm×1.01 cm。

表2 梓树属不同种继代培养性状比较
2.3 生根培养性状及移栽成活率比较
由表3可知,培养30 d后,生根率黄金树最高,为73.33%,滇楸最低,为25%,由高到低为黄金树>梓树>楸树>灰楸>滇楸;发根数梓树最高,为3.54个,滇楸最低,为 0.64;根长楸树最长,为1.82 cm,灰楸最短,为0.86 cm;瓶苗生长以黄金树最快,30 d后新发芽芽长为1.23 cm,楸树最慢,为0.22 cm,由快到慢为梓树>黄金树>灰楸>滇楸>楸树;移栽成活率黄金树(85.5%)>梓树(82.4%)>楸树(70.4%)>灰楸(56.3%)>滇楸(43.3%)。
方差分析表明,再生芽生根指标生根率(P=0)、生根数(P=0)、根长(P=0)、发芽数(P=0)、叶数(P=0.002)及生根苗生长速度(芽长P=0)在不同种间差异达到极显著水平;观测指标方差分量生根率(96.381%)、生根数(97.647%)、根长(95.113%)、发芽数(95.669%)、芽长(96.274%),除叶数量(89.966)外,都达到95%以上;这说明梓树属生根培养瓶苗生长指标受较强的遗传因素制约,各种在生长性状上存在着丰富的遗传变异,种的不同是影响生长性状的主要因素。
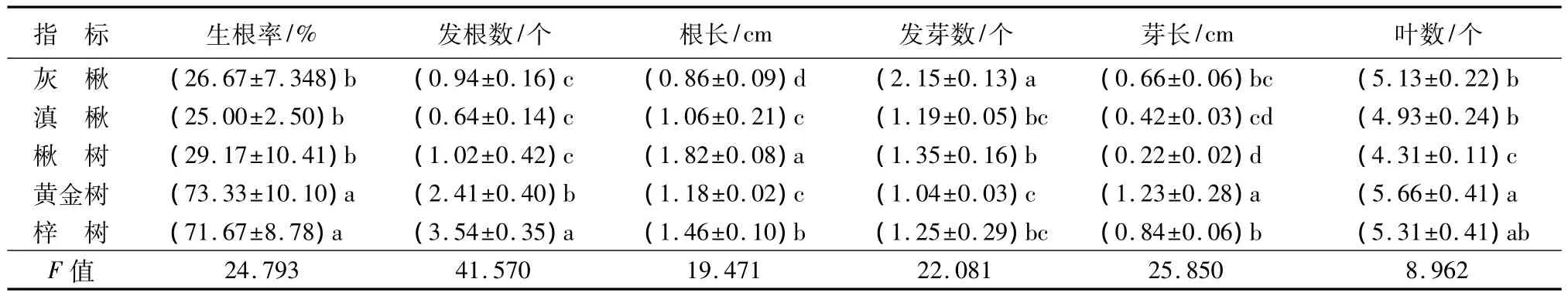
表3 梓树属生根培养性状比较
3 结论与讨论
植物组织培养是利用植物细胞的全能性,通过一定的营养物质、激素和其他外界条件的作用,诱导发育成完整的植株个体,已广泛的应用于苗木生产、种质资源保存、转基因体系的应用等。珍贵阔叶树种种质创新课题组研究表明,在同等培养环境下,影响楸树[11]无性系组培繁殖的主要因素是遗传基因,外界条件在不同的培养阶段亦有不同的影响,本试验研究表明,5个种的继代培养与生根培养生长指标分量均达到90%左右,说明梓树属瓶苗生长指标受较强的遗传因素制约,各种在瓶苗生长性状上存在着丰富的遗传变异,种的不同是影响梓树属不同种组培繁殖生长性状的主要因素;通过对组培繁殖瓶苗生长情况、生根率与移栽成活率比较,综合表现以黄金树较好。对同种不同无性系间对比研究的较多,北美悬铃木[12]、杉木、桉树[13]、楸树已有相关研究报道,得出的结论基本类似,在同一培养环境下,无性系繁殖指标受较强的遗传因素制约。在相同的环境下,不同群体间的表型差异就反映了它们基因型的差异[14],这说明种组培繁殖表型性状的差异在一定的程度上表现出种间基因的远近,关于组培表型性状与基因远近的研究少有报道,这还需进一步深入研究。
“十一五”期间,珍贵阔叶树种种质创新课题组对灰楸、滇楸、楸树、黄金树、梓树进行了资源分布、田间表型性状调查,并进一步进行了无性系选择、目标树选择和培育等研究,研究结果表明,梓树属分为3个组(灰楸、滇楸与楸树为一组,黄金树与梓树为一组,紫葳楸为一组)。黄金树、梓树与紫薇楸主要以种子繁殖为主,种子结实率、发芽率较高,黄金树、梓树实生苗主要用于楸树、灰楸优良无性系的嫁接砧木;灰楸、楸树种子结实率、发芽率都较低,主要以嫁接、组织培养繁殖,其生长迅速、材质优良是珍贵优质用材及庭院观赏、道路绿化树种。
林木组织培养分化主要指标在于继代增殖分化芽数、芽的生长、生根率与移栽成活率,本试验结论表明,这些指标灰楸、滇楸、楸树无性系性状表现相近,梓树与黄金树相近,这与田间表现基本相同。大多研究表明,不同植物组织培养需要不同的配方与培养环境,很少有将同属植物做组织培养对比分析研究,本试验研究表明,影响组培分化诱导与各生长性状的主要因素是植物本身遗传因素。根据细胞全能性原理,植物组培的遗传因素完全与母本相同,林木植物组培所表现的表型性状,一般都作为供试配方的筛选依据,从初代外植体诱导分化、继代培养、生根诱导及移栽炼苗性状的表现,很少有研究将此列入育种选育的依据,只是辅助的扩繁育种材料或为转基因做遗传载体,组培性状的表现能否作为林木遗传育种选择的依据,这还需大量的不同植物相关研究来做基础,倘若能够成立,这无疑能够缩短育种进程,推进林木产业的发展。同时,为拓宽组培应用领域,在育种选择过程中能否将植物组培与田间表型性状的表现统一追踪观测,根据两者数据的统计与分析,以得出相应的基础理论,为进一步深入研究与发展做基奠,这还需相关研究领域的人员的涉及与深入。
[1]赵曦阳,王军辉,张金凤,等.梓树属4个种种子表型性状和发芽特性的研究[J].西北农林科技大学学报:自然科学版,2008,36(12):149-154.
[2]黄金生,樊汝波,姜力.梓树属植物花及叶细胞中核内含体的研究[J].植物学报,1989,31(4)323-324.
[3]杨玉珍,王顺财,彭方仁.我国楸树研究现状及开发利用策略[J].林业科技开发,2006,20(3):4-7.
[4]王念,乇巧玲.梓树属植物基因组DNA提取及RAPD体系优化[J].河南林业科技,2012,32(1):1-3.
[5]王改萍,鲁丹,彭方仁,等.不同梓属树种花器官蛋白电泳分析[J].林业科技开发,2012,26(3):54-57.
[6]于永明,王军辉,马建伟,等.LaCl3对楸树无性系试管苗生长的影响[J].东北林业大学学报,2011,39(1):31-33.
[7]于永明,王军辉,张宋智,等.二次回归正交设计在楸树离体生根培养中的应用[J].南京林业大学学报:自然科学版,2011,35(4):47-50.
[8]江荣翠,彭方仁,谭鹏鹏,等.楸树体细胞胚胎发生的研究[J].南京林业大学学报:自然科学版,2010,34(2):15-18.
[9]任莺,彭方仁,杨燕,等.楸树不同类型胚性和非胚性愈伤组织的细胞学观察[J].林业科技开发,2011,25(5):43-45.
[10]甘肃省小陇山林业实验局,甘肃民勤连古城国家自然保护区管理局.DB62/T 2013—2011楸树无性系组培快繁技术规程[S].兰州:甘肃省质量技术监督局.
[11]于永明,王军辉,马建伟,等.楸树无性系离体培养特性差异研究[J].西北植物学报,2012,32(1):199-204.
[12]蔡晓明,杨立伟,施季森.北美悬铃木无性系间组培特性差异研究[J].江苏农业科学,2008(4):89-93.
[13]吴大忠.不同杉木优良无性系组培诱导特性的研究[J].福建林业科技,1999,26(1):26-29.
[14]续九如.林木数量遗传学[M].北京:高等教育出版社,2006.
[15]曹孜义,刘国民.实用植物组织培养技术教程[M].兰州:甘肃科学技术出版社,1996.
[16]Feng Daling,Zhang Jie,Liu Xia,et al.In vitro culture of immature embryos from Koelreuteria bipinnata var.integrifoliola[J].Forestry Studies in China,2009,11(3):179-184.
[17]Zhong Deyi,Zhu Youyin,Liu Qian,et al.Production of embryogenic callus and plant regeneration from elite Guizhou waxy maize inbred lines[J].Agricultural Sciences in China,2011,10(4):490-498.
