农业景观格局与麦蚜密度对其初寄生蜂与重寄生蜂种群及寄生率的影响
2013-09-15关晓庆刘军和赵紫华
关晓庆,刘军和,赵紫华,*
(1.宁夏大学农学院,银川 750021;2.黄淮学院生物工程学院,驻马店 463000;3.中国科学院动物研究所农业虫害鼠害综合治理研究国家重点实验室,北京 100101)
随着景观生态学的理论与方法在昆虫生态学中的广泛应用,景观结构影响昆虫的种间关系得到多次的研究证实[1-4],包括寄生蜂对寄主的寻找效应寄生率[5-7]。高度的异质性与多生境组成是农业景观格局的重要特点,其中非作物生境为寄生蜂提供食物、避难所、适宜的微气候环境、转移寄主或这些资源的结合[8-9]。寄生蜂在非作物环境中寻找并利用大多数资源(花粉与花蜜),在作物生境中仅仅是为寻找寄主[7,10]。因此与简单农业景观相比,复杂农业景观中提供的更大丰富度与多样性的资源可能会为寄生蜂提供更好的资源与生境[11],例如花粉、花蜜、避难所以及越冬环境等等。而且最近的研究证明农业景观中非作物生境的丰富度与多样性显著影响寄生蜂的多样性和丰富度[5,7,11-12]。
寄主密度与寄生蜂的寄生率也存在正相关关系,寄主密度的增加有利于寄生蜂对寄主的寻找和寄生,但通常存在一定的滞后效应。而且这些研究大多是特异性的寄主-初寄生蜂系统,也就是种群水平上的研究,寄主密度对初寄生蜂组成,尤其是对重寄生蜂组成的影响,从群落水平上的研究还很少[5,13]。初寄生蜂的寄主为麦蚜,重寄生蜂的寄主为初寄生蜂,形成了一个完整的麦蚜-初寄生蜂-重寄生蜂系统,因此在群落水平上研究寄生蜂的反应意义更大。景观的复杂性能够增加寄生蜂多样性[6],寄主密度也能够增加寄生蜂的多样性,但在不同的农业景观中寄主密度与寄生率关系并不清楚[6,13]。景观格局与寄主密度对寄生蜂的交互作用研究很多还是未知,尤其是对重寄生蜂的影响研究更为少见。Marino&Landis在密歇根的研究表明春季复杂农业景观下的寄生蜂对Pesudaletia unipuncta的寄生率更高[11],然而在其他季节初寄生率并无明显差异,可能因为初寄生蜂的优势种的变化与他们在不同农业景观下生活史特征的差异[14]。春季Meteorus communis为优势种,在复杂农业景观格局下表现出高的寄生率,秋季Glytapanteles militaris成为优势种,受农业景观格局的影响不显著。然而在不同农业景观格局下寄主密度对寄生率的影响却一直没有受到关注。本文研究了2种农业景观格局下3种不同的寄主密度对麦蚜初寄生蜂及重寄生蜂的影响,探讨农业景观格局与寄主密度对麦蚜初寄生蜂与重寄生蜂群落多样性的影响及交互作用。基于国内外文献,本文提出了以下假说:1)景观结构的复杂性能够影响麦蚜寄生蜂的群落结构和种群密度,寄主密度同样影响麦蚜寄生蜂的种群和群落结构;2)比起农业景观结构的复杂性,寄主密度可能对麦蚜寄生蜂的种群及群落组成影响更为重要,景观结构与寄主密度间存在着强烈交互作用。这些对揭示麦蚜种群活动规律的变化、蚜虫生物防治及预测预报具有重要意义。
1 材料与方法
1.1 研究区域
研究区域设在宁夏银川平原,银川平原处于温带干旱地区,日照充足,年均日照时数3000h左右,无霜期约160d。热量资源较丰富,10℃以上有效积温约3300℃。气温日较差大,平均达13℃,有利于小麦的生长发育和营养物质积累。年降水量200mm左右,黄河年均过境水量达300余亿m3,灌溉条件较好,小麦种植历史较长,是小麦的优质生产基地,选择了2种不同的农业景观,对麦田麦田-天敌群落进行调查。
1)复杂农业景观 本文选取了2个农场的复杂农业景观,分别为银川西夏区军马场与银川市兴庆区掌政。西夏区军马场试验地(38°32'35N,106°7'58E),作物种植类型多样,而且是温棚设施农业集中区域,面积为0.05万hm2,居民区较多,农田、道路、林地、杂粮与荒地交错纵横,为复杂的农业景观格局,主要种植小麦、玉米和蔬菜为主,小麦斑块面积较小,选择军马场一队棚间空地小麦斑块27块,平均面积85.19m2;兴庆区掌政五度桥(38°25'45N,106°22'21E)是典型的农耕区,但种植模式,作物种类多样,形成典型的城镇居民区、设施温棚区和零散麦田的农业景观镶嵌体,土地面积广,条田林网纵横,小麦,玉米,水稻,杂粮与枸杞形成特有的生态农业景观,选择五度桥小麦斑块21块,平均面积732.67 m2;
2)简单农业景观 兴庆区掌政(38°25'21N,106°22'21E)是银川平原的小麦主产区,种植模式单一,更低比例较大,非作物生境很少,面积为0.5万hm2,小麦种植广泛,面积较大,作物品种单一,仅有部分水稻种植,随逐年降水减少,水稻产区逐步衰退,小麦面积逐年扩大,选择小麦斑块22块,平均面积4356.88m2。调查区为春麦区,近年有小面积冬麦种植。
1.2 研究方法
研究对象包括麦长管蚜Macrosiphum avenae(F.)、麦二叉蚜Schizaphis graminum(Rond)、禾缢管蚜Rhopalosiphum padi(L.)、初寄生蜂群落以及重寄生蜂群落。以不同面积的小麦田块为样地,GPS定位,共研究调查了小麦斑块70块,分别在小麦的麦蚜迁入期、增长期与高峰期调查5次。
僵蚜调查与寄生蜂的饲养:调查方法为棋盘式五点取样法,根据田块特点分为东,南,西,北,中5个方位,每个方位随机选择100株小麦,采取目测和计数相结合的方法,每100株小麦观察并记录15—20min,分别记录100株小麦上麦蚜的种类和数量,每块样地采集到的所有僵蚜分别装入指形瓶,带回实验室饲养至成虫以待鉴定,每块样地的僵蚜放入一个培养皿,用采集日期与样地代号编号,放入光照培养箱中饲养,饲养条件(L∶D=16∶8),共饲养40d以上,直至僵蚜中无新寄生蜂羽化。没有羽化的僵蚜在实验室内进行解剖,观察没有羽化的原因,连同蚜尸放入酒精,以待制作玻片标本鉴定[4,15-26]。
农田边缘植被调查:在每种农业景观下的农田边缘选择3块10m×10m的样方,进行每木调查。调查时5m×5m为调查单位,记录木本植物胸高(以离地130cm为胸高标准)的周长,测量出以划红线作为标记;鉴定每株树木的种类并记录数量,投影法并估算每株树木的高度,草本植物不列入调查范围。
不同寄主密度的划分:对麦蚜防治指标研究较多,本文采用500头/百株为防治指标[24]。调查数据中0—200头/百株为低寄主密度,200—500头/百株为高寄主密度,麦田中500头/百株以上的麦蚜就已经进入了防治期,农药的干扰导致昆虫群落的破坏,因为不采用500头/百株以上的麦田数据。
1.3 分析方法
采用成对F检验比较2中不同的农业景观、麦蚜种群密度、寄生率与捕食性率等的差异,并进行Tukey显著性检验。本文采用的显著性没有注明的情况下均为0.05。
2因素2水平的方差分析(ANOVA):将农业景观因子与寄主种群密度作为两个处理因素,农业景观因子有2个水平(简单与复杂),寄主种群密度同样有2个水平(低寄主密度与高寄主密度),分别计算农业景观因子和寄主密度对寄生性天敌个捕食性天敌的影响以及二者间的交互作用。本文采用的显著性没有注明的情况下均为0.05。
以上数据处理分析及作图均采用Microsoft Office Excel与SAS 8.2(Statistics Analysis System 8.2,SAS Institute Inc.)数据处理系统进行。
2 结果与分析
2.1 农业景观结构与麦蚜在不同农业景观结构中的分布
2.1.1 不同农业景观结构中的植被分布
不同农业景观下麦田边缘的植被组成结构差异很大。复杂农业景观下麦田边缘比简单景观的非作物生境更宽,平均高度也更高(表1)。复杂农业景观中树种更为丰富,树木与灌木的比例也比简单景观中的要高。复杂景观种树木的优势种为杨树、臭椿与刺槐,分别占总树木的32.2%、35.6%与9.5%。简单景观中树木的优势种为刺槐与野枸杞,分别占总树木的43.2%与26.5%。

表1 不同农业景观下麦田边缘植被特点及数据分析Table 1 Analysis and Characteristics of wheatfields variables(mean±SE)in different agricultural landscapes
2.1.1 农业景观结构对麦蚜分布的影响
复杂农业景观中的麦长管蚜与禾溢管蚜种群数量大于简单农业景观;简单农业景观中麦二叉蚜的种群数量较高,复杂农业景观中与简单农业景观中的总蚜量分别为(234.5±45)与(254±59.2)头/百株(F1,30=1.36,P=0.370),3种蚜虫种群密度差异都不显著。麦二叉蚜迁入麦田的时间稍晚于麦长管蚜,麦二叉蚜有翅蚜在5月2日左右开始迁飞进入麦田,较麦长管蚜晚15d左右,简单农业景观的麦田有翅蚜迁入量显著高于复杂景观的麦田。景观结构对3种麦蚜以及总麦蚜的种群数量影响不显著,但简单农业景观中有翅蚜的迁入量高于复杂的农业景观。

表2 景观结构对麦蚜种群的影响Table 2 Effects of landscape suructure on wheat aphids
2.2 寄主密度与景观格局对寄生蜂种群分布的影响
2.2.1 寄主密度与景观格局对初寄生蜂种群分布的的影响
当低寄主密度(平均为135.0头/百株)时,复杂农业景观与简单农业景观中初寄生蜂寄生率无明显差异(F1,19=2.26,P=0.28),但复杂农业景观中初寄生蜂8种,分别为燕麦蚜茧蜂Aphidius avenae、烟蚜茧蜂A.gifuensis、四川蚜茧蜂A.sichuanensis、混合柄瘤蚜茧蜂Lysiphlebus confusus、翼蚜外茧蜂Praon volucre、东方蚜外茧蜂P.orientale、缢管蚜外茧蜂P.rhopalosiphum、亚洲三叉蚜茧蜂Trioxys asiaticus、三叉蚜茧蜂T.sp.、弓蚜茧蜂Toxares sp.与蚜小蜂Phelinus sp.,简单景观中只有5种,比复杂景观种缺少翼蚜外茧蜂Praon volucre、东方蚜外茧蜂P.orientale与缢管蚜外茧蜂P.rhopalosiphum,其余种类一致。当高寄主密度时,平均为286.0头/百株,景观结构对初寄生蜂寄生率也无明显差异(F1,14=2.96,P=0.34),复杂景观中初寄生蜂为11种,简单景观中9种(表3)。
在复杂农业景观下,寄主密度对初寄生率的影响显著(F1,17=36.96,P=0.01);在简单农业景观下,寄主密度对初寄生率的影响同样显著(F1,16=46.93,P=0.01)。高寄主密度下采集的初寄生蜂是低寄主密度下的1.5倍,而高密度寄主是低密度寄主的1.4倍。不同农业景观下两种寄主密度对初寄生蜂的影响是相似的。
在两种寄主密度下,燕麦蚜茧蜂与烟蚜茧蜂都为初寄生蜂的优势种,分别占总初寄生蜂的74.49%与19.94%(表3)。在以前的文献中,麦蚜初寄生蜂只分布于蚜茧蜂属、蚜外茧蜂属与柄瘤蚜茧蜂属,本文的研究鉴定表明还包括三叉蚜茧蜂属、弓蚜茧蜂属与蚜小蜂属的5种。尽管初寄生蜂的优势种优势度很高,但非优势种的数量也表现出了明显的寄主密度效应(F1,30=5.36,P=0.05)。景观结构虽对初寄生蜂的寄生率无显著影响,但对初寄生蜂的种类影响较大(F1,30=7.62,P=0.02),初寄生蜂多样性受寄主密度影响显著(F1,30=17.85,P=0.01)。
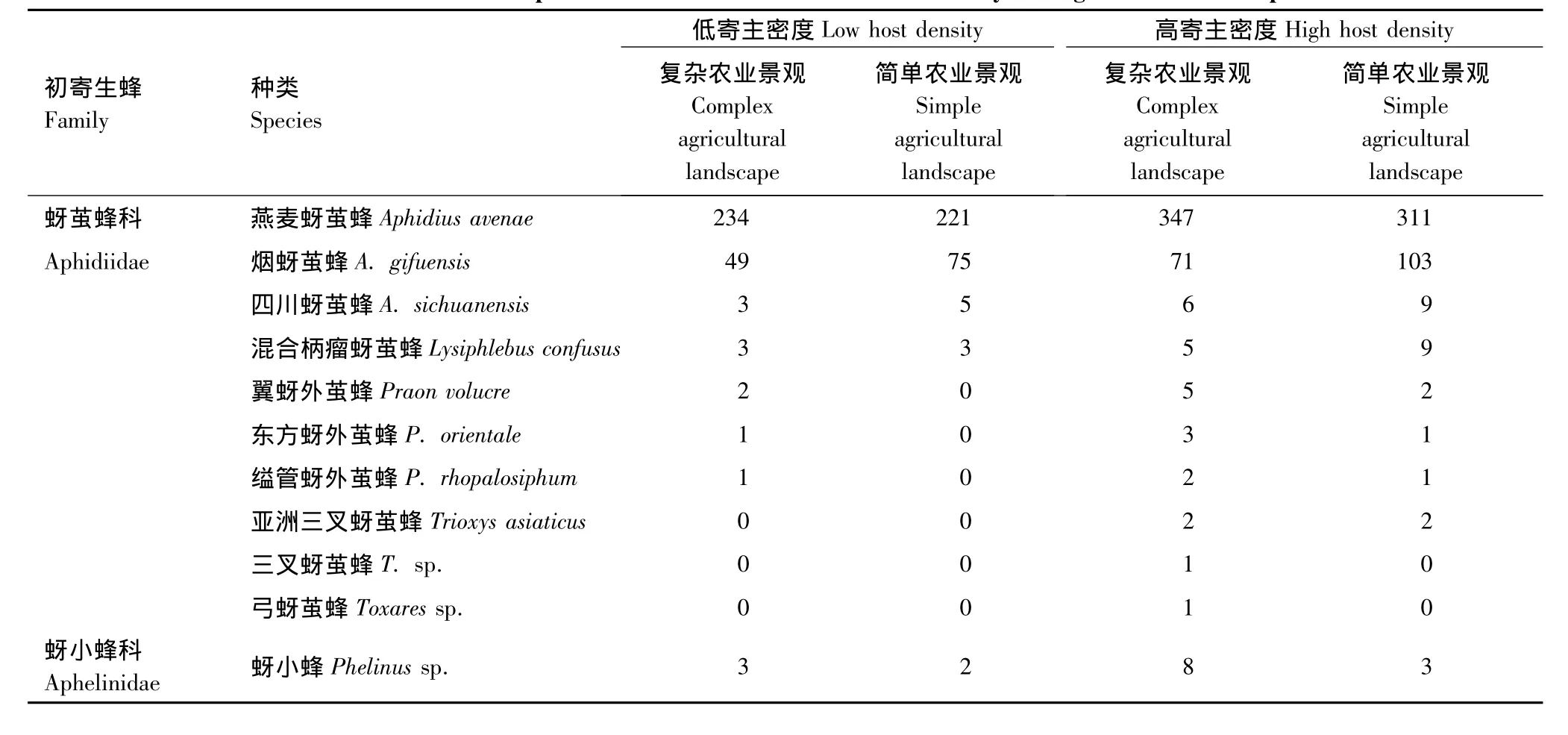
表3 不同寄主密度与农业景观下初寄生蜂的种类与数量Table 3 Numbers and Species of Parasitoids in different host density and agricultural landscapes
2.2.2 寄主密度与景观格局对重寄生蜂种群分布的影响
当低寄主密度(平均为135.0头/百株)时,复杂农业景观与简单农业景观中重寄生蜂寄生率无明显差异(F1,19=2.65,P=0.35),但复杂农业景观中初寄生蜂10种,蚜茧蜂长背瘿蜂Alloxysta sp.1、蚜茧蜂长背瘿蜂A.sp.2、蚜茧蜂长背瘿蜂A.sp.3、蚜虫宽缘金小蜂Pachyneuron aphidis、宽肩阿莎金小蜂Asaphes suspensus、蚜茧蜂金小蜂Asaphes vulgaris、金小蜂Pteromalidae sp.、蚜虫跳小蜂Aphidencyrtus aphidivorus、啮小蜂Tetrastichus sp.与合沟细蜂Dendrocerus carpenteri。简单景观中同样也有10种,种类一致。当高寄主密度时,平均为(286±76.69)头/百株,景观结构对重寄生蜂寄生率也无明显差异(F1,14=3.06,P=0.46),复杂景观中重寄生蜂为10种,简单景观中同样10种(表4)。
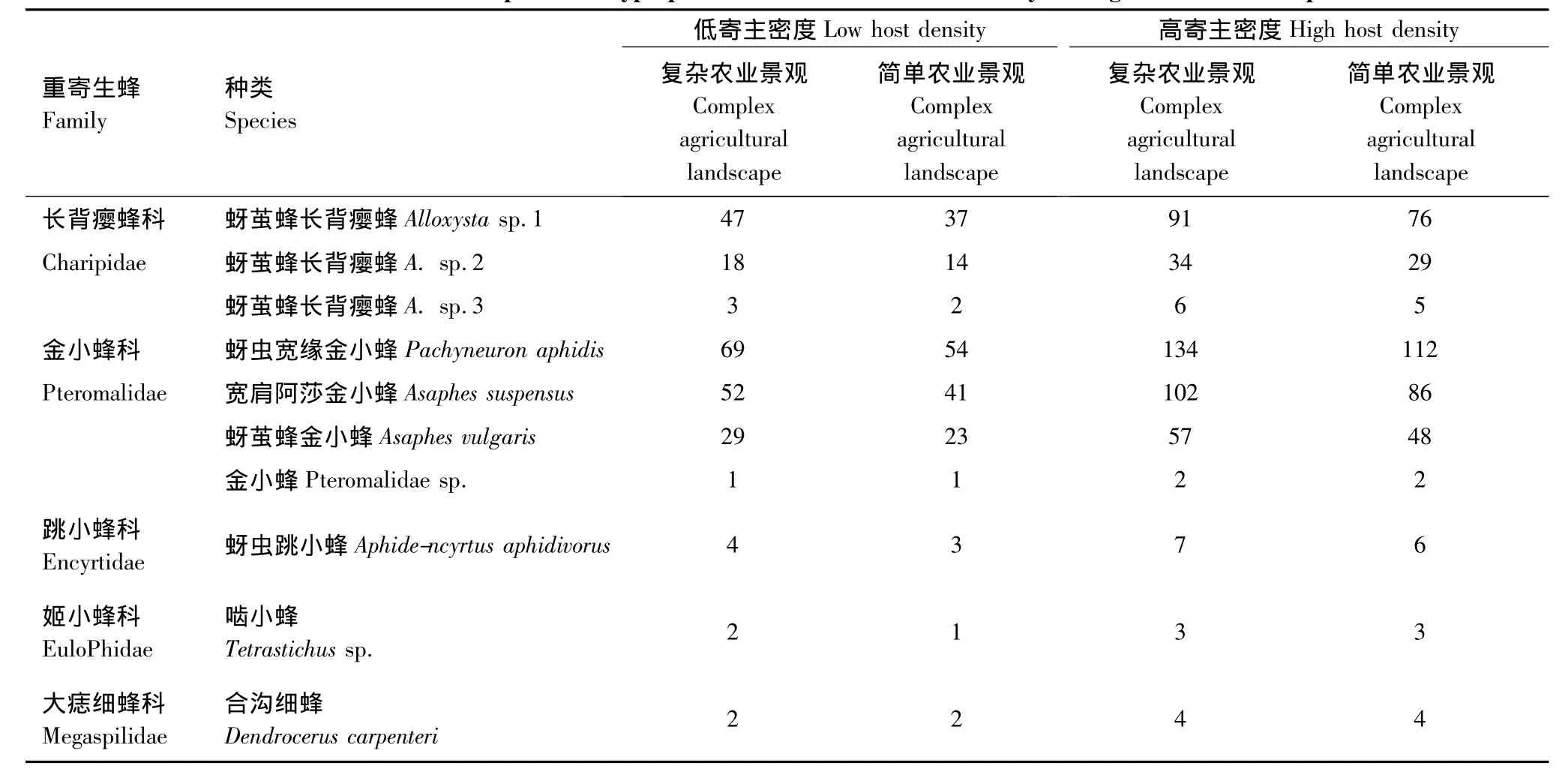
表4 不同寄主密度与农业景观下初寄生蜂的种类与数量Table 4 Numbers and Species of Hyperparasitoids in different host density and agricultural landscapes
在复杂农业景观下,寄主密度对重寄生率的影响显著(F1,17=75.62,P=0.01);在简单农业景观下,寄主密度对重寄生率的影响同样显著(F1,16=69.69,P=0.01)。高寄主密度下采集的重寄生蜂是低寄主密度下的2.1倍,而高密度寄主是低密度寄主的1.4倍。不同农业景观下两种寄主密度对重寄生蜂的影响是相似的。
在两种寄主密度下,蚜虫宽缘金小蜂与蚜茧蜂长背瘿蜂都为寄生蜂的优势种,分别占总重寄生蜂的30.23%与20.26%(表4)。在以前的文献中,麦蚜重寄生蜂只分布于蚜茧蜂长背瘿蜂属、阿金小蜂属、白木细蜂属与宽缘金小蜂属,本文的研究鉴定表明还包括啮小蜂属、蚜虫跳小蜂属的2种。重寄生蜂的优势种不是非常明显,但非优势种的数量也表现出了明显的寄主密度效应(F1,30=81.26,P=0.01)。景观结构对初寄生蜂的寄生率无显著影响,对初寄生蜂的种类影响也不大(F1,30=1.62,P=0.62),重寄生蜂种类受寄主密度影响也不显著(F1,30=2.02,P=0.41)。
2.2.3 寄主密度与景观格局对总寄生蜂丰富度与多样性的影响
在低寄主密度下,景观结构对总寄生蜂丰富度的影响显著(F1,19=25.03,P=0.01),复杂景观中总寄生蜂为19种,简单景观中仅为15种,但景观结构对总寄生蜂的多样性影响并不显著(F1,19=3.09,P=0.21),复杂景观与简单景观中多样性指数分别为3.24与2.87;高寄主密度下,景观结构对总寄生蜂丰富度的影响也不显著(F1,14=4.01,P=0.13),复杂景观中总寄生蜂为21种,简单景观中仅为19种,景观结构对总寄生蜂的多样性影响也不显著(F1,14=3.82,P=0.15),复杂景观与简单景观中多样性指数分别为5.48与4.39。
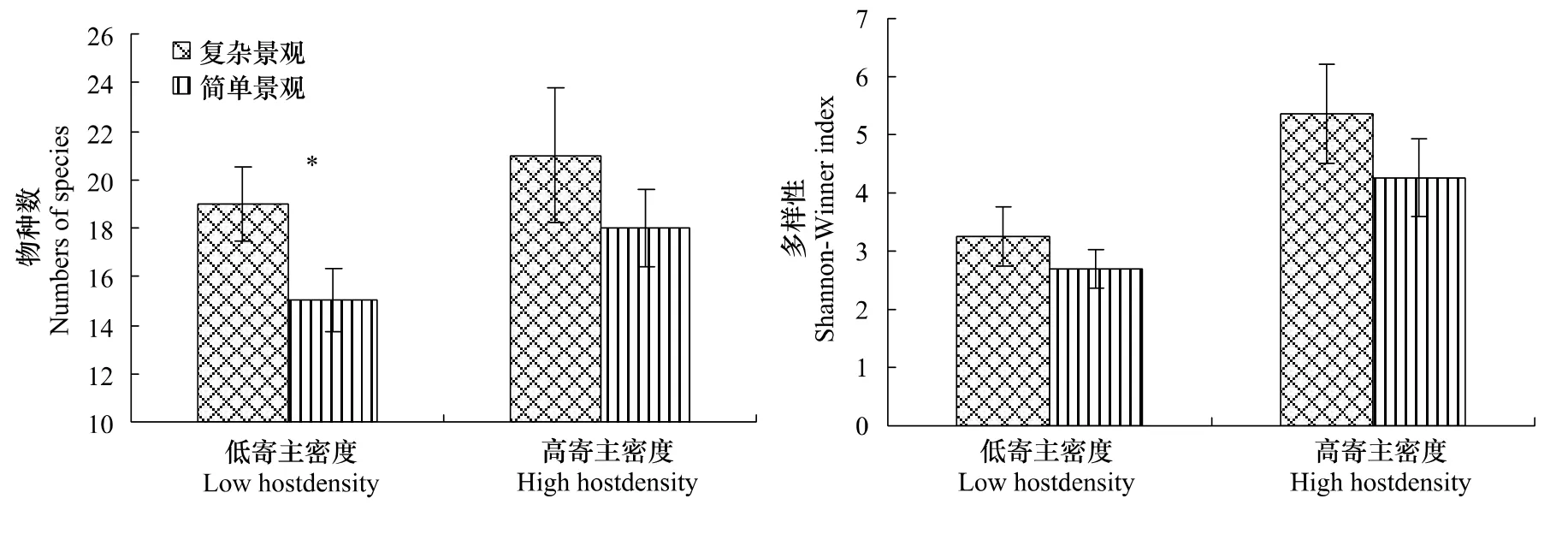
图1 不同农业景观下两种不同寄主密度的寄生蜂的丰富度与多样性Fig.1 Mean ±SD parasitoid species richness and diversity obtained wheat aphids at low and high densities in simple and complex agricultural landscapes
在复杂农业景观下,寄主密度对总寄生蜂丰富度的影响不显著,高寄主密素中总寄生蜂为21种,简单景观中为19种,但寄主密度对总寄生蜂的多样性影响显著(F1,17=35.16,P=0.01),低寄主密度与高寄主密度中多样性指数分别为3.24与5.48(图1);简单农业景观下,寄主密度同样对总寄生蜂丰富度的影响显著(F1,16=5.32,P=0.05),复杂景观中总寄生蜂为19种,简单景观中仅为15种,寄主密度对总寄生蜂的多样性影响也及显著(F1,16=11.01,P=0.01),复杂景观与简单景观中多样性指数分别为2.87与4.39(图1)。
2.3 寄主密度与景观格局对寄生蜂的交互作用
寄主密度、景观结构及他们间的交互作用对麦蚜初寄生蜂与重寄生蜂的影响(表5),寄主密度对初寄生蜂的影响显著(P<0.05),对重寄生蜂的影响极显著(P<=0.05),但景观结构对初寄生蜂及重寄生蜂的影响均不显著,寄主密度与景观结构间也无明显的交互作用。
2.4 寄主密度与景观结构对麦蚜寄生蜂寄生率的影响
在低寄主密度下,复杂景观与简单景观的初寄生蜂寄生率无明显差异(F1,19=3.36,P=0.14),初寄生率分别为26.2%与21.8%,复杂景观与简单景观重寄生蜂寄生率也无显著差异,与初寄生蜂一致,在高寄主密度下也表现出同样的特征,复杂景观与简单景观中初寄生蜂与重寄生蜂的寄生率均无显著差异;在简单农业景观中,低寄主密度与高寄主密度的初寄生蜂寄生率无明显差异,初寄生率分别为26.2%与31.52%,但重寄生蜂寄生率差异显著(F1,16=31.35,P=0.01),重寄生率分别为35.26%与59.62%(图2)。在复杂农业景观中,低寄主密度与高寄主密度的初寄生蜂寄生率差异显著(F1,17=6.36,P=0.05),初寄生率分别为21.8%与28.62%,重寄生蜂也差异显著(F1,4=1.02,P=0.05),重寄生率分别为38.26%与52.32%(图2)。
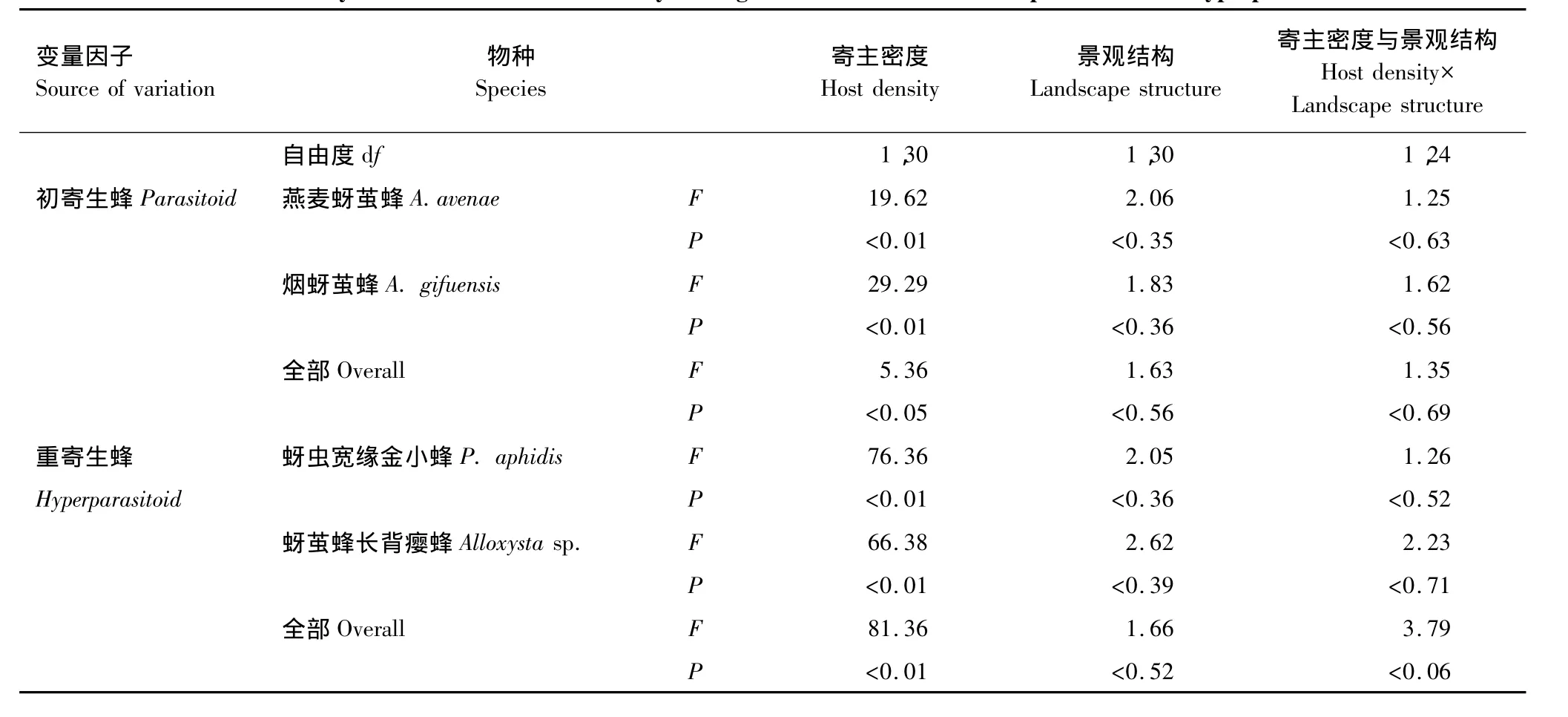
表5 寄主密度及农业景观结构对初寄生蜂与重寄生蜂优势种寄生率地影响分析Table 5 Analysis of the effect of host density and agricultural structure on the parasitism and hyperparasitism
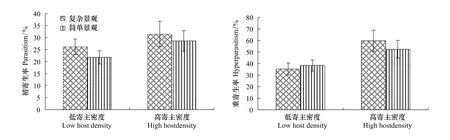
图2 不同农业景观结构下不同寄主密度对初寄生率与重寄生率的影响Fig.2 Percent parasitism and hyperparasitism(mean±SD)of wheat aphids and parasitoids at low and high densities in simple and complex agricultural landscapes
3 讨论
麦蚜初寄生蜂在不同农业景观中无明显差异,但寄主密度对两种初寄生蜂寄生率影响显著,寄主密度与景观格局无明显交互作用;麦蚜重寄生蜂在不同农业景观中也无明显区别,但寄主密度同样对两种初寄生蜂寄生率影响显著,寄主密度与景观格局对重寄生蜂有弱的交互作用。本文首次研究证实不同寄主密度与农业景观格局对初寄生蜂优势种燕麦蚜茧蜂A.avenae与烟蚜茧蜂A.gifuensis对麦蚜寄生率、重寄生蜂优势种蚜虫宽缘金小蜂P.aphidis与蚜茧蜂长背瘿蜂Alloxysta sp.1对麦蚜的重寄生率,以及对麦蚜寄生蜂群落多样性与丰富度的影响。由于对初寄生蜂非优势种四川蚜茧蜂A.sichuanensis、混合柄瘤蚜茧蜂Lysiphlebus confusus、翼蚜外茧蜂Praon volucre、东方蚜外茧蜂P.orientale、缢管蚜外茧蜂P.rhopalosiphum、亚洲三叉蚜茧蜂Trioxys asiaticus、三叉蚜茧蜂T.sp.、弓蚜茧蜂Toxares sp.、蚜小蜂Phelinus sp.与重寄生蜂非优势种蚜茧蜂长背瘿蜂A.sp.2、宽肩阿莎金小蜂Asaphes suspensus、蚜茧蜂金小蜂Asaphes vulgaris、金小蜂Pteromalidae sp.、蚜虫跳小蜂Aphidencyrtus aphidivorus、啮小蜂Tetrastichus sp.、合沟细蜂Dendrocerus carpenteri生物学特征与生态学特性的缺乏,因此主要集中在2类寄生蜂优势种的讨论。
本论文鉴定的共21种寄生蜂全部直接或间接以麦蚜作为寄主,形成了一个完整的寄主-初寄生蜂-重寄生蜂群落。群落中不同寄生蜂对寄主密度及景观结构的复杂性具有不同的反应[1,6,27],同一种寄生蜂在不同时空尺度内对景观格局的反应也存在变异。寄主密度与寄生蜂多样性形成一种正相关关系,寄生蜂对景观格局和寄主密度的不同反应可能受产卵时间、行为、体型和多种与寄主有关的环境因素影响[27]。燕麦蚜茧蜂与烟蚜茧蜂对寄主密度的不同反应可能由于不同的生活史特征和体型大小。由于群居产卵特征与较短的雌性产卵时间,烟蚜茧蜂在麦蚜发生初期种群密度较小时种群数量较大,随寄主密度的上升,由于温度的升高和种间竞争,体型较大的燕麦蚜茧蜂由于具有较长的生活史,在种间竞争中逐步获胜,成为优势种。对烟蚜茧蜂与燕麦蚜茧蜂的更细致的行为学特征,包括田间对寄主种群的搜寻效应,对寄主及植物特征的识别,能够进一步的揭示寄生蜂对寄主密度反应的潜在机制。重寄生蜂也表现出相似的特征,蚜虫宽缘金小蜂在麦蚜发生初期种群数量较大,随时间的推移与重寄生蜂的种间竞争,蚜茧蜂长背瘿蜂逐渐在竞争中获胜,成为麦蚜发生后期的优势种。
初寄生蜂与重寄生蜂的总寄生率受景观结构的复杂性影响不显著[28],这与 Costamagna对粘虫Pseudaletia unipunctas寄生蜂研究的结论一致[29],但也与很多研究景观结构与生物过程间相互作用的结论相反[3,5,14,30-31]。对不同寄主密度对初寄生蜂及重寄生蜂群落寄生率与多样性影响显著,但有关寄主密度的研究较少,仅发现Costamagna对粘虫的研究,结果也是一致的[29]。在麦蚜的发生期寄生蜂优势种的转变可能与寄主丰富度的波动、成虫的食物资源、自然种群循环、分布型、竞争作用或气候因子有关[1,14,29]。
尽管有文献表明景观结构的复杂性与寄生率呈正相关,农业景观中增加非作物生境能够提高寄生蜂的寄生率,而提高自然天敌的生物控害功能。但在试验研究中,我们没有得到景观结构复杂性与寄生率的相关关系,只发现了寄主密度与寄生率的正相关关系[11,14,32-33],不同寄生蜂对寄主密度的变化反应也有差异,这种不同的反应可能与寄生蜂群落在相同的时空范围内竞争同一资源长期性形成的进化机制有关。本文认为这种初寄生蜂与重寄生蜂群落的演化受景观结构的复杂性影响不大,而可能受景观格局中的植物种类有关,尤其是1年生或多年生的开花植物,这些植物可以为寄生蜂提供花粉、花蜜与适宜的微气候,甚至成为农田生境受干扰后的避难所或越冬场所,这些植物与寄生蜂间的关系很多是特异性的,寻找这种初寄生蜂与特异性植物间的关系,有些植物能够为初寄生蜂提供转移寄主和食物资源,有些植物能够为重寄生蜂提供寄主和食物资源,因此寻找为初寄生蜂提供资源的植物,排除为重寄生蜂提供资源的特异性植物可能是将来保护性生物防治的重要研究方向[8,34-36],农业景观中应用开花植物与植物群落来增加天敌是生境管理的重要理论之一,目前仍然是保护性生物防治研究中较新的一方面。生境管理的研究主要集中在探求植物提供的资源如何给加强天敌的生物、生态学功能与抑制害虫爆发的能力,核心技术是在连续的时空范围内农业景观格局的构建与设计,从景观格局上提高天敌生存活动的连续性,切断害虫存活的时空连续性,最大程度的抑制害虫种群。因此农业景观格局中的植物种类、非作物植物的密度与物候期的研究,以及非作物生境的比例和格局对寄生蜂群落的多样性与功能具有更重要的意义[37]。
[1] He D H.Landscape structure and control of insect pests in agro-ecosystems.Plant Protection,2009,35(3):12-15.
[2] Zhen Y K,You M S.Biological diversity in support of ecologically-based pest management at landscape level.Acta Ecologica Sinica,2009,29(3):1508-1517.
[3] Roland J,Taylor P D.Insect parasitoid species respond to forest structure at different spatial scales.Nature,1997,386(6626):531-538.
[4] Stary P.Parasitoids and biocontrol of Russian wheat aphid,Diuraphis noxia(Kurdj.)expanding in central Europe.Journal of Applied Entomology,1999,123(5):273-279.
[5] Kruss A.Effects of landscape structure and habitat type on a plant-herbivore-parasitoid community.Ecography,2003,26(3):283-290.
[6] Doak P.The effects of plant dispersion and prey density on parasitism rates in a naturally patchy habitat.Oecologia,2000,122(4):556-567.
[7] Thies C,Steffan-Dewenter I,Tscharntke T.Effects of landscape context on herbivory and parasitism at different spatial scales.Oikos,2003,101(1):18-25.
[8] Landis D,Menalled F.Ecological considerations in conservation of parasitoids in agricultural landscapes//Barbosa P,ed.Conservation Biological Control.San Diego:Academic Press,1998.
[9] Gurr G M,Wratten SD,Luna J M.Multi-function agricultural biodiversity:pest management and other benefits.Basic and Applied Ecology,2003,4(2):107-116.
[10] Tscharntke T.Parasitoid populations in the agricultural landscape//Hochberg M E,Ives A R,eds.Parasitoid Population Biology.Princeton NJ:Princeton University Press,2000.
[11] Marino P C,Landis D A.Effect of landscape structure on parasitoid diversity and parasitism in agroecosystems.Ecological Applications,1996,6(1):276-284.
[12] Kruss A,Tscharntke T.Species richness and parasitism in a fragmented landscape:experiments and field studies with insects on Vicia sepium.Oecologia,2000,122(1):129-137.
[13] Sugiura S,Osawa N.Temporal responseof parasitoids to the density of the leafroller Eudemisgyrotis(Lepidoptera:Tortricidae)on bayberry Myrica rubra(Myricaceae).Environmental Entomology,2002,31(6):988-994.
[14] Menalled F D,Costamagna A C,Marino P C,Landis D A.Temporal variation in the response of parasitoids to agricultural landscape structure.Agriculture,Ecosystems and Environment,2003,96(1/3):29-35.
[15] Chen J H,Shi Q X.Systematic Studies on Aphidiidae of China(Hymenoptera:Aphidiidae).Fuzhou:Fujian Science and Technology Press,2001.
[16] Liao D J,Li X L,Pang X F.Economic Insect Fauna of China Fasc.34.Hymenoptera Chalcidoidea.Beijing:Science Press,1987.
[17] Chai Z Q,Deng JH,Wu W.Biology of pachyneuron aphidis-the hyperparasitoid of Myzus persicae.Journal of Southwest Forestry College,2005,25(2):56-58.
[18] Chai Z Q,Liao Q P,Zhu J Q,Wu W.Research situation on aphid hyperparasitoids.Tropical Agricultural Science and Technology,2008,31(3):47-51.
[19] Zong L B,Lei C L,Liu Z F.Aphid parasites(Hymenoptera:Aphididae)in wheat fields of Wuchang.Chinese Journal of Biological Control,1986,(2):75-79.
[20] Zhu Z L,Xi X,Song T H.Biological characteristics of aphid parasites(Hymenoptera:Aphididae)in wheat fields.Journal of Environmental Entomology,1982,4(2):18-21.
[21] Ji Z D,Zhen M Y,Yang SS,Zhao JZ.Liu J J,Song J.The studies of parasites on wheat aphids in the south-central area of Hebei Province.Journal of Agricultural University of Hebei,1985,8(3):61-65.
[22] Zhen Y S,Dong D Z,Yang H F.The species of aphid parasites(Hymenoptera:Aphididae)in wheat fields.Journal of Environmental Entomology,1987,9(2):94-97.
[23] Ren X J,Gao Z L,Dang Z H,Li Y F,Pan W L.The species and distribution of Aphidiidae in wheat fields in Hebei Province.Journal of Hebei Agricultural Sciences,2007,11(1):37-40.
[24] Li SG,Liu A Z,Liu S M.Study on the relationships among wheat aphids,natural enemy and yield loss of wheat and the action threshold of aphids.Acta Phytophylacia Sinica,1994,21(1):15-19.
[25] French B W.Seasonal occurrence of aphids and natural enemies in wheat and associated crops.Southwestern Entomologist,2001,26(1):49-57.
[26] Kavallieratos N G,TomanovicˇŽ,Stary P,Athanassiou C G,Sarlis G P,PetrovicˇO,NiketicˇM,Veroniki M A.A survey of aphid parasitoids(Hymenoptera:Braconidae:Aphidiinae)of Southeastern Europe and their aphid-plant associations.Applied Entomology and Zoology,2004,39(3):527-563.
[27] Teder T,TanhuanpääM,Ruohomäki K,Kaitaniemi P,Henriksson J.Temporal and spatial variation of larval parasitism in non-outbreaking populations of a folivorous moth.Oecologia,2000,123(4):516-524.
[28] Vollhardt I M G,Tscharntke T,Wäckers F L,Bianchi F JJA,Thies C.Diversity of cereal aphid parasitoids in simple and complex landscapes.Agriculture Ecosystems and Environment,2008,126(3/4):289-292.
[29] Costamagna A C,Menalled F D,Landis D A.Host density influences parasitism of the armyworm Pseudaletia unipuncta in agricultural landscapes.Basic and Applied Ecology,2004,5(4):347-355.
[30] Östman Ö,Ekborn B,Bengtsson J.Landscapeheterogeneity and farmingpractice influence biological control.Basic and Applied Ecology,2001,2(4):365-371.
[31] Zhao Z H,Guan X Q,He D H.Community composition of parasitoids and hyperparasitoids of wheat aphids in different agricultural landscapes.Chinese Journal of Applied Entomology,2012,49(1):220-228.
[32] Menalled F D,Marino P C,Gage S H,Landis D A.Does agricultural landscape structure affect parasitism and parasitoid diversity?Ecological Applications,1999,9(2):634-641.
[33] Thies C,Roschewitz I,Tscharntke T.The landscape context of cereal aphid-parasitoid interactions.Proceedings of the Royal Society B:Biological Sciences,2005,272(1559):203-210.
[34] Zhao Z H,He D H,Hang J,Shi Y,Zhao Y S,Wang Y.Minimum amounts of suitable habitat for wheat aphid,parasitoid,and hyperparasitoid in facility-based agricultural landscapes.Chinese Journal of Applied Ecology,2011,22(1):206-214.
[35] Zhao Z H,Shi Y,He D H,Hang J,Zhao Y S,Wang Y.Population dynamics of wheat aphids in different agricultural landscapes.Acta Ecologica Sinica,2010,30(23):453-462.
[36] Zhao Z H,He D H,Hui C.From the inverse density-area relationship to the minimum patch size of a host-parasitoid system.Ecological Research,2012,27(2):303-308.
[37] Zhao Z H,Wang Y,He D H,Guan X Q,Xin M.Effects of landscape structure and key landscape factors on aphids-parasitoids-hyper parasitoids populations in wheat fields.Acta Ecologica Sinica,2012,32(2):472-482.
参考文献:
[1] 贺达汉.农业景观与害虫种群控制.植物保护,2009,35(3):12-15.
[2] 郑云开,尤民生.农业景观生物多样性与害虫生态控制.生态学报,2009,29(3):1508-1517.
[15] 陈家骅,石全秀.中国蚜茧蜂膜翅目:蚜茧蜂科.福州:福建科学技术出版社,2001
[16] 廖定熹,李学骝,庞雄飞.中国经济昆虫志第三十四册膜翅目:小蜂总科.北京:科学出版社,1987.
[17] 柴正群,邓建华,吴伟.烟蚜重寄生蜂-蚜虫宽缘金小蜂生物学特性研究.西南林学院学报,2005,25(2):56-58.
[18] 柴正群,廖启平,朱建青,吴伟.蚜虫重寄生蜂研究概况.热带农业科技,2008,31(3):47-51.
[19] 宗良炳,雷朝亮,刘作甫.麦田蚜茧蜂的初步研究.生物防治通报,1986,(2):75-79.
[20] 朱子龙,习学,宋亭华.麦田蚜茧蜂的生物学特性.昆虫天敌,1982,4(2):18-21.
[21] 季正端,郑明义,杨树森,赵建洲,刘建军,宋健.河北省中南部地区的小麦蚜虫寄生蜂.河北农业大学学报,1985,8(3):61-65.
[22] 郑永善,董大志,杨海峰.麦蚜蚜茧蜂种类.昆虫天敌,1987,9(2):94-97.
[23] 任雪娟,高占林,党志红,李耀发,潘文亮.河北省麦田蚜茧蜂的种类与分布.河北农业科学,2007,11(1):37-40.
[24] 李世功,刘爱芝,刘素梅.麦蚜与天敌相互关系研究及麦蚜防治指标初报.植物保护学报,1994,21(1):15-19.
[31] 赵紫华,关晓庆,贺达汉.农业景观结构对麦蚜寄生蜂群落组成的影响.应用昆虫学报,2012,49(1):220-228.
[34] 赵紫华,贺达汉,杭佳,石云,赵映书,王颖.设施农业景观下破碎化麦田麦蚜及寄生蜂种群的最小适生面积.应用生态学报,2011,22(1):206-214.
[35] 赵紫华,石云,贺达汉,杭佳,赵映书,王颖.不同农业景观结构对麦蚜种群动态的影响.生态学报,2010,30(23):453-462.
[37] 赵紫华,王颖,贺达汉,关晓庆,辛明.麦蚜和寄生蜂对农业景观格局的响应及其关键景观因子分析.生态学报,2012,32(2):472-482.
