模拟酸雨对蒙古栎幼苗生长和根系伤流量的影响
2013-09-15梁晓琴丁文娟常瑞英王仁卿
梁晓琴,刘 建,丁文娟,常瑞英,王仁卿,,*
(1.山东大学环境研究院,济南 250100;2.山东大学山东省植被生态工程技术中心,济南 250100;3.山东大学生命科学学院生态学与生物多样性研究所,济南 250100)
酸雨是指pH值小于5.6的雨水,也包括雪、雾、雹等其它形式的酸性降水,科学上称作酸沉降[1]。我国已成为继欧洲、北美之后世界上的第三大酸雨片区,酸雨区的覆盖面积占我国国土面积的40% 左右,酸雨危害已相当严重[2]。自20世纪60年代以来,随着酸雨污染的日益严重及其生态影响的日益扩大,酸雨问题越发地受到各国政府和科研人员的重视[3]。
在陆地生态系统中,植物是酸雨污染的主要受体[3],因此,酸雨对植物影响的研究是生态学的热点。国外学者分别从植物细胞、植物个体和种群与群落组成的不同水平上分析了酸雨对植物生长的影响[4-6],国内学者对酸雨问题及酸雨对植物影响的研究也屡有报道,但多集中在西南地区,北方地区由于酸雨污染较轻,所以对这方面的研究较少。但是,近20年来,我国酸雨区范围明显向北持续扩展,且酸雨酸度增强趋势明显,局部地区如北京、山西、河南和山东等部分省、直辖市的酸雨和强酸雨频率在2008年达到自1993年来的最高值[7]。酸雨检测资料分析显示我国北方地区酸雨污染形势明显加重[1-2,7]。因此,研究酸雨对于北方地区典型树种的影响具有重要意义。
栎类是我国北方阔叶树种中非常重要的一个属,在温带的针阔混交林、暖温带的落叶阔叶林和亚热带的常绿阔叶林中,栎属的一些种都占有非常重要的地位[8]。其中,蒙古栎在栎林中面积最大,是温带针阔混交林区域地带性植被类型松林的重要伴生树种之一,同时是在松林被干扰后,形成次生阔叶林面积比例最大的建群树种,是我国北方阔叶林的典型树种之一[9]。
国内外在进行模拟酸雨对植物生长影响的实验研究中,多是在单因素水平上根据pH值设置不同的酸雨酸度水平,结合其他因素的多因素实验,如不同臭氧水平,添加不同钙、镉离子浓度等,虽有所涉及,但相对较少[10-12]。而结合降雨量因素,分析酸雨酸度和降雨量双因素对植物生长的影响还鲜有报道。因此,本研究通过人工模拟不同酸度和降雨量水平的酸雨,研究其对蒙古栎幼苗的影响,试图回答问题:(1)酸雨对蒙古栎幼苗的生长是否造成伤害;(2)酸雨的酸度和降雨量因素分别对蒙古栎幼苗的生长造成什么影响;(3)两因素间是否存在交互作用,以探讨我国北方日趋严重的酸雨是否会影响蒙古栎幼苗的生长,为酸雨区森林恢复植物的选择提供依据。
1 材料与方法
1.1 实验材料及研究地点
蒙古栎(Quercus mongolica Fisch.ex Ledeb.),壳斗科栎属,落叶乔木,主要分布于我国东北三省、内蒙古和河北省北部,为我国落叶阔叶林的主要组成树种和我国的主要用材树种。蒙古栎喜光、耐寒、喜凉爽气候;根系发达,耐干旱耐瘠薄、耐火烧、不耐盐碱、喜中性至酸性土壤。本实验选用从种子站购买成熟的蒙古栎种子作为实验材料。
模拟实验于2011年4月至9月在山东莱芜市房干村山东大学房干生态学教学科研基地(北纬36°26',东经117°27')进行。该地气候为暖温带季风型大陆气候,年平均气温约12.4℃,年平均降雨量为600—830 mm,主要分布在 7、8、9 月份,四季分明,雨热同期[13],地带性土壤为棕壤[14-15]。
1.2 实验设计
将购买的成熟蒙古栎种子于2011年4月放入人工气候箱中进行催芽萌发,后栽种于内径23 cm、深30 cm的塑料花盆中,每盆1株。取当地土壤(棕壤土)作为栽培土,土壤基本理化性质为:有机质含量27.86 g/kg,总氮1.29 g/kg,有效磷24.07 mg/kg,有效钾85.38 mg/kg,pH为6.54。实验前保证幼苗充足供水,并进行常规田间管理。
经过前期适应性生长后,选取长势基本一致、平均株高约15 cm的蒙古栎幼苗于人工气候大棚中开始进行控制实验。将其随机设置为4个酸雨酸度与3个酸雨降雨量处理的组合,每个处理6个重复,共72株,即4个酸雨处理 ×3个降雨量处理 ×6重复。根据山东地区酸雨检测分析资料[16-17],按SO24-∶NO-3体积分数 12∶1配置母液,用自来水(pH 6.5—7)稀释成pH值分别为2.5(重度)、3.5(中度)、4.5(轻度)和5.6(CK)的酸雨溶液。根据山东省自有气象记录以来的夏季均降雨量410 mm[18,19],计算并设置自然降雨量处理(R)的每盆植株实验期总喷淋量约为17.03 L,分别增减30% 设置为增雨处理(R+)和减雨处理(R-)。喷淋于6月底开始,进行3个月处理,每个月喷淋4次,每7d喷1次。
1.3 测定指标与方法
从实验处理开始,每15d测量1次生长指标,共6次,用钢性卷尺测量株高,并记录叶片数目。
蒙古栎幼苗根系伤流量的测定:在实验结束时,进行根系伤流夜的收集并测定伤流量[20-21]。在试管内塞入一定量的脱脂棉,用塑料薄膜封口,准确称出试管重量。每个处理组内选取4株蒙古栎幼苗,在临近傍晚时,将茎部距根约2 cm处剪断,下部茎段自小孔插入试管与脱脂棉轻轻接触,收集约15 h,到第2天清晨取下试管再次准确称重。两次质量之差即为植株根系伤流量。
实验结束后,将幼苗整棵挖出清洗干净,分为地下和地上两部分,在80℃下烘干至恒重。称量两部分生物量并计算生物量分配的相关参数,包括地下生物量比(BMR,地下生物量/总生物量)、地上生物量比(AMR,地上生物量/总生物量)、根冠比(R/S,地下生物量/地上生物量)。
1.4 统计分析
利用SPSS 13.0(SPSSInc.IL,USA)对数据进行正态检验和方差齐性检验后进行单因素与双因素的ANOVA分析,比较各个参数分别在不同酸雨酸度与酸雨降雨量水平下的差异,以及两因素间是否存在交互作用。存在显著差异的进行邓肯(Duncan)多重比较,并用字母进行标记。绘图由Origin 8.0(OriginLab Co.,Massachusetts,USA)完成。
2 实验结果
2.1 模拟酸雨对蒙古栎幼苗生物量的影响
双因素方差分析的结果表明,酸雨降雨量对蒙古栎幼苗地上、地下和总生物量均有显著性影响(表1),而酸雨酸度的影响不显著,两因素之间没有明显的交互作用。从图1中可以看出,在低酸雨酸度下,蒙古栎幼苗生物量呈现出随着降雨量增加而增加的趋势,并且在酸雨酸度最低(pH 2.5)时表现的尤为显著。当酸雨降雨量相同时蒙古栎幼苗生物量指标的表现不甚一致,但是在增雨处理(R+)内,当酸雨酸度最低(pH 2.5)时蒙古栎幼苗的生物量指标表现出显著的增加。虽然模拟酸雨对蒙古栎幼苗生物量分配方面的影响不显著,但其基本的变化趋势与生物量的变化趋势相同。
2.2 模拟酸雨对蒙古栎幼苗生长指标的影响
降雨量的增多直接导致了土壤含水量的增加(表1),进而使得蒙古栎幼苗的株高和叶片数均有显著
增加。而酸雨酸度对蒙古栎幼苗生长的影响不显著,同时与降雨量没有显著的交互作用。但在减雨(R-)和自然雨量(R)处理内,蒙古栎幼苗的株高和叶片数基本表现出pH 2.5>pH 3.5<pH 4.5<pH 5.6的趋势;同样在增雨处理(R+)酸雨酸度最大(pH 2.5)时,蒙古栎幼苗株高的生长和叶片数明显的增加。
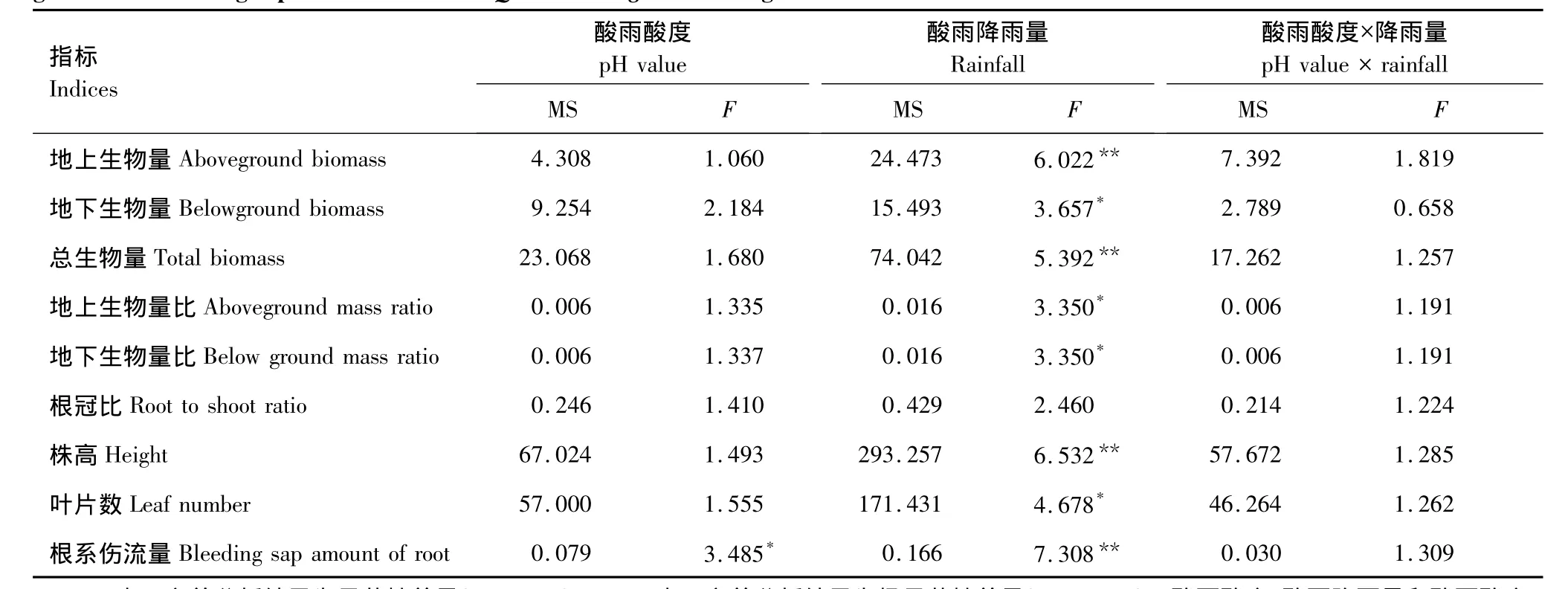
表1 不同酸雨酸度和降雨量处理对蒙古栎幼苗生物量及其分配、生长指标和根系伤流量影响的F值Table 1 F value of two-way ANOVA of the effects of different p H value and rainfall of simulated acid rain on biomass,biomass allocation,growth and bleeding sap amount of root of Quercus mongolica seedings
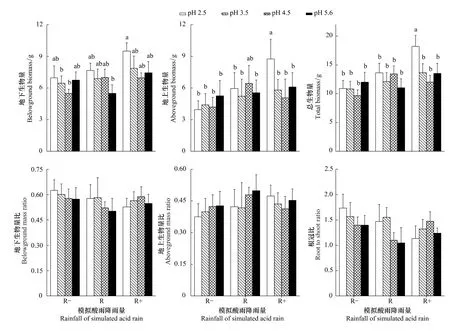
图1 模拟不同酸雨酸度和降雨量处理下蒙古栎幼苗生物量和生物量分配的变化(平均值±标准误)Fig.1 Effects of different p H value and rainfall of simulated acid rain on biomass and biomass allocation of Quercus mongolica seedlings(mean±SE)不同处理间字母不同表示存在显著差异(P<0.05)
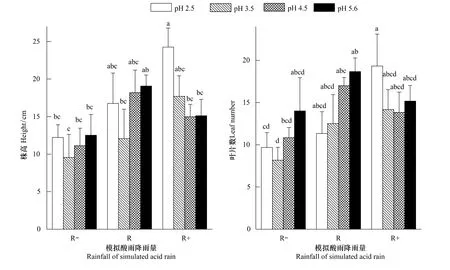
图2 模拟不同酸雨酸度和降雨量处理下蒙古栎幼苗生长指标的变化(平均值±标准误)Fig.2 Effects of different pH value and rainfall of simulated acid rain on growth of Quercus mongolica seedlings(mean ± SE)不同处理间字母不同表示存在显著差异(P<0.05)
2.3 模拟酸雨对蒙古栎幼苗根系伤流量的影响
根系伤流量是表征根系活力的指标之一,作为重要的根系生理活动指标,其大小直接影响植株对营养物质的吸收,进而影响地上部分的生长发育[22-23]。由于根压作用,从植株茎部的切口处溢出液滴的现象被称为根系伤流,伤流量的大小可以反映根系生理活动的强弱[24]。本实验结果显示,酸雨降雨量的增加极显著地增强了蒙古栎幼苗的根系伤流量。而酸雨酸度的增加使得根系伤流量显著减弱,蒙古栎幼苗根系受到伤害。酸度和降雨量间没有明显交互影响作用。
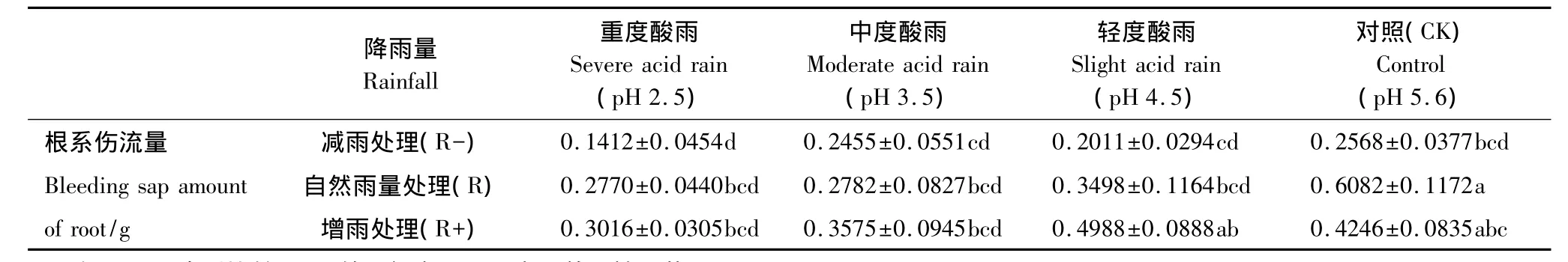
表2 不同酸度和降雨量酸雨对蒙古栎幼苗根系伤流量的影响(平均值±标准误)Table 2 Effects of different p H value and rainfall of simulated acid rain on bleeding sap amount of root of Quercus mongolica seedlings(mean±SE)
3 讨论
水分是限制植物生长的主要决定因子之一,降水变化可以从不同尺度影响植物生理及生态学过程,改变其水分利用策略[25-26]。本实验选择模拟酸雨降雨量和酸度,研究不同的酸雨降雨水平对蒙古栎幼苗生长的影响,结果显示在本实验的酸雨强度下,降雨量减少对幼苗总生物量的影响表现为减少其累积[26-27],而植株分配给根系的生物量却有所增加,蒙古栎幼苗根冠比增大,表明幼苗将更多的生物量分配给地上部分而减少了地下部分的投入[28];随降雨量的减少,幼苗的株高等形态指标也逐渐降低[29]。酸雨降雨量对根系伤流量的影响表现为随着降雨量的增大,蒙古栎幼苗的根系伤流量增强。而根系作为植物吸收水分和矿质影响的重要器官,根系伤流量的高低,对植物地上部分生长具有直接的影响[20],根系伤流量加大会促进植株地上部分的生长和生物量的累积[30],这又进一步验证了本研究中酸雨降雨量这一因素对蒙古栎幼苗的影响结果。
过去模拟酸雨对植物影响的研究大多为模拟高强度酸雨酸度,特别是pH值低至2—2.5的情况,在此条件下,酸雨对植物叶片会造成明显伤害、抑制植物幼苗生长、降低生物量累积等。但仍有部分研究证明,酸雨处理可能促进某些阔叶树种的生长,提高幼苗的生物量,其原因可能是酸雨中硝酸根离子的添加增加了土壤的氮含量,因此对植物产生了施肥作用,从而有利于植物幼苗的生长[31-33]。本研究根据中国北方的酸雨现状并预测若酸雨污染严峻后可能的高酸度酸雨情况,设置不同酸度水平的模拟酸雨,发现酸雨酸度增加对蒙古栎幼苗的株高生长、叶片数增加和生物量累积有一定的促进作用,这支持了酸雨处理具有施肥效应的假说[34],特别是在酸雨酸度和降雨量同时达到最大时,大量累积的硝酸根离子使得这一处理下的蒙古栎幼苗在生长和生物量指标的各个方面都达到最大值。
酸雨对植物的影响一方面在于酸雨淋溶对地上部分的直接影响,另一方面在于酸雨在导致土壤酸化的过程中对植物的间接影响[3]。而酸雨造成的土壤酸化对植物的直接影响主要表现在对根系的伤害。本研究结果表明,随着酸雨酸度的增加,蒙古栎幼苗的根系伤流量明显降低,说明重度酸雨对植物根系造成了伤害,虽然在对地下生物量累积方面没有显著影响,但是根冠比的增加显示蒙古栎幼苗通过增加植株对地下生物量的投入来抵抗这一伤害。
蒙古栎作为我国落叶阔叶林的主要组成树种,对环境具有广泛的适应性。目前对蒙古栎适应性的研究多集中在其对温度、CO2和光能等方面的适应,而研究其对酸雨的适应情况的文章相对较少,进一步将酸雨酸度和降雨量双因素的影响结合在一起综合分析其适应情况的研究则更具有意义。酸雨对植物生长的影响表现在酸雨酸度、酸雨降雨量和酸雨频率等方面,本研究仅模拟在其他条件相同的情况下酸雨酸度和降雨量对蒙古栎幼苗的影响。结果显示,酸雨降雨量对蒙古栎幼苗的生长在多方面均表现出显著性影响,而酸雨酸度仅影响了幼苗的根系伤流量,相对于酸雨酸度,降雨量对蒙古栎生长起到了主效作用。二者的交互作用不显著,但是酸雨酸度最大降雨量也最大的条件对蒙古栎幼苗的生长具有明显的促进作用,说明蒙古栎是对酸雨具有抗性的树种,而满足其适生条件的土壤为偏酸性土壤也显示出蒙古栎是一种耐酸甚至喜酸的植物,这与金清等的研究结果阔叶树幼苗对酸胁迫具有一定的适应和缓冲能力相符合[35]。
我国是煤炭资源比较丰富的国家,从能源消费结构来看,煤炭依然在我国能源消费中占主导地位。这就决定了在未来相当长的一个时期内,酸雨都将会作为我国一个严重的环境问题而存在[36]。所以,在我国已成为全球第三大酸雨区,且酸雨区范围明显向北持续扩展、酸雨酸度增强趋势明显的情况下,通过研究酸雨对蒙古栎幼苗生长的影响,有利于进一步开展酸雨区酸雨对陆地生态系统的影响研究。其次,通常酸雨对植物造成的伤害首先是使植物生长变得缓慢,随之,植物为生态系统固定的碳量减少[37],酸雨区内植物碳库含量的减少将导致这一区域碳循环的失衡,所以选择对酸雨抗性较好、缓冲能力较高、适应性较强的植物,对维持酸雨区生态系统的碳平衡具有重要意义。另外,选择既是我国北方地区的典型阔叶树种又具有酸雨抗性的植物作为酸雨区森林恢复重建的候选树种[35],对于在区域环境胁迫下生态系统结构和功能的恢复可能具有重要的意义。本研究的结果可为我国已知及潜在的酸雨区内筛选诸如蒙古栎这样适应性较强、抗酸雨能力较高的植物作为植被构建物种提供理论参考依据。
本研究中酸雨对蒙古栎幼苗的影响是在人工气候大棚的层次上进行的短期实验,而自然条件下酸雨对植物的影响是十分复杂的,而且具有长期的累积效应,其影响因素众多,需要进行相关因素的复合影响实验。同时,测定不同处理下根系、叶片氮含量随时间的变化,叶片光合作用相关指标等将有助于深入理解酸雨对北方植物的影响及其可能的氮施肥效应。
[1] Zhao Y X,Hou Q.Characteristics of the acid rain variation in China during 1993—2006 and associated causes.Acta Meteor Sinica,2010,24(2):239-250.
[2] Wu D,Wang SG,Shang K Z.Progress in research of acid rain in China.Arid Meteorology,2006,24(2):70-77.
[3] Qi Z M,Wang X D,Song G Y.The research progress of the effect of acid rain on plant.World Sci-tech Research& Develepment,2004,26(2):36-41.
[4] Ferenbaugh R W.Effects of simulated acid rain on Phaseolus vulgaris L.(Fabaceae).American Journal of Botany,1976,63(3):283-288.
[5] Lee JJ,Weber D E.The effect of simulated acid rain on seedling emergence and growth of eleven woody species.Forest Science,1979,25(3):393-398.
[6] Ulrich B,Mayer R,Khanna P K.Chemical changes due to acid precipitation in a loess-derived soil in central Europe.Soil Science,1980,130(4):193-199.
[7] Zhang X M,Chai F H,Wang S L,Sun X Z,Han M.Research progress of acid precipitation in China.Research of Environmental Sciences,2010,23(5):527-532.
[8] Feng Q H,Shi Z M,Dong L L,Liu SR.The response of functional traits of Quercus species to meteorological factors in temperate zone of NSTEC.Acta Ecologica Sinica,2010,30(21):5781-5789.
[9] Yu SL,Ma K P,Xu C B,Jin SF,Song X B,Chen L Z.The species diversity characteristics comparison of Quercus mongolica community along environmental gradient factors.Acta Ecologica sinica,2004,24(12):2932-2939.
[10] Chappelka III A H,Chevone B I.White ash seedling growth response to ozone and simulated acid rain.Canadian Journal of Forest Research,1986,16(4):786-790.
[11] Liu T W,Wu F H,Wang W H,Chen J,Li Z J,Dong X J,Patton J,Pei Z M,Zheng H L.Effects of calcium on seed germination,seedling growth and photosynthesis of six forest tree species under simulated acid rain.Tree Physiology,2011,31(4):402-413.
[12] Liao B H,Liu H Y,Lu SQ,Wang K F,Probst A,Probst J L.Combined toxic effects of cadmium and acid rain on Vicia faba L.Bulletin of Environmental Contamination and Toxicology.2003,71(5):998-1004.
[13] Zhang X Q,Liu J,Welham C V J,Liu C C,Li D N,Chen L,Wang R Q.The effects of clonal integration on morphological plasticity and placement of daughter ramets in black locust(Robinia pseudoacacia).Flora,2006,201(7):547-554.
[14] Wang R Q,Zhou G Y.Shandong Vegetation.Jinan:Shandong Science and Technology Press,2000:259-266.
[15] Fang Y,Meng Z N,Zhu X Z.Shandong forest plant resources and application.Beijing:China International Radio Press,2006:41-42.
[16] Liu H B,Wang X T.The city's atmospheric precipitation pH observation of Shandong and initial analysis.Environmental Monitoring in China,2006,22(6):92-95.
[17] Zhen W D.Study on conditions and genesis of acid precipitation in cities in Shandong Province[D].Jinan:Shandong University,2005.
[18] Chi Z P.Spatial and temporal distributions and climatic change of summer precipitation in Shandong Province.Plateau Meteorology,2009,28(1):220-226.
[19] Xu Z X,Meng C L,Zhao F F.Long-term trend analysis for temperature and precipitation in Shandong Province.Scientia Meteorologica Sinica,2007,27(4):387-393.
[20] Llugany M,Lombini A,Poschenrieder C,Dinelli E,BarcelóJ.Different mechanisms account for enhanced copper resistance in Silene armeria ecotypes from mine spoil and serpentine sites.Plant and Soil,2003,251(1):55-63.
[21] Yin Y D,Qiu L Q,Wei M,Yang F J,Wang X F.Effects of CO2enrichment in greenhouse on root growth as well as root exudates and bleaching sap of cucumber seedlings.Acta Ecologica Sinica,2010,30(7):1860-1867.
[22] Shen B,Wang X.Physiological activities of root system in two inter-subspecific hybrid rice combinations.Chinese Journal of Rice Science,2002,16(2):146-150.
[23] Cao SQ,Deng Z R,Zhai H Q,Tang Y L,Han GB,Zhang R X,Sheng SL,Gong H B,Yang T N.Analysis on heterosis and combining ability for bleeding sap amount of root and its declined properties in indica hybrid rice.Chinese Journal of Rice Science,2002,16(1):19-23.
[24] Shi X D,Liu Y F,Wen Z Q,Wang WW.Research progress in plant root bleeding.Journal of Anhui Agriculture Science,2006,34(10):2043-2045.
[25] Lindner M,Maroschek M,Netherer S,Kremer A,Barbati Anna,Garcia-Gonzalo J,Seidl R,Delzon S,Corona P,Kolstrom M,Lexer M J,Marchetti M.Climate change impacts,adaptive capacity,and vulnerability of European forest ecosystems.Forest Ecology and Management,2010,259(4):698-709.
[26] Wu JL,Wang M,Lin F,Hao Z Q,Ji L Z,Liu Y Q.Effects of precipitation and interspecific competition on Quercus mongolica and Pinus koraiensis seedlings growth.Chinese Journal of Applied Ecology,2009,20(2):235-240.
[27] Wang M,Li QR,Hao ZQ,Dong B L.Effects of soil water regimes on the growth of Quercusmongolica seedlings in Changbai Mountains.Chinese Journal of Applied Ecology,2004,15(10):1765-1770.
[28] Wang X P,Fang J Y,Zhu B.Forest biomass and root-shoot allocation in northeast China.Forest Ecology and Management,2008,255(12):4007-4020.
[29] Xu F,Guo W H,Xu W H,Wang R Q.Effects of water stress on morphology,biomass allocation and photosynthesis in Robinia pseudoacacia seedlings.Journal of Beijing Forestry University,2010,32(1):24-30.
[30] Song X,Zeng D H,Lin H M,Yamaguchi T,Nishizaki Y,Yamada P.Effects of peat and weathered coal on activity of rice root system and its nutritional absorption.Chinese Journal of applied ecology,2001,12(6):867-870.
[31] Fan H B,Wang Y H.Effects of simulated acid rain on germination,foliar damage,chlorophyll contents and seedling growth of five hardwood species growing in China.Forest Ecology and Management,2000,126(3):321-329.
[32] Dixon M J,Kuja A L.Effects of simulated acid rain on the growth,nutrition,foliar pigments and photosynthetic rates of sugar maple and white spruce seedlings.Water,Air and Soil Pollution,1995,83(3/4):219-236.
[33] Shan Y F.Acid rain,air pollution and vegetation.Beijing:China Environmental Science Press,1993:153-155.
[34] Shelburne V B,Reardon JC,Paynter V A.The effects of acid rain and ozone on biomass and leaf area parameters of shortleaf pine(Pinusechinate Mill.).Tree Pysiology,1993,12(2):163-172.
[35] Jin Q,Jiang H,Yu SQ,Jin J,Wang Y H.Research on the growth and photosynthesis of typical seedlings in subtropical regions under acid rain stress.Acta Ecologica Sinica,2009,29(6):3322-3327.
[36] Liu L Q,Liu C X,Sun Z Y.A survey of China's low-carbon application practice-Opportunity goes with challenge.Renewable and Sustainable Energy Reviews,2011,15(6):2895-2903.
[37] Fan H B,Huang Y Z,Yuan Y H,Li Y Y,Huang R Z,Fan H Y.Carbon cycling of forest ecosystems in response to global nitrogen deposition:a review.Acta Ecologica Sinica,2007,27(7):2997-3009.
参考文献:
[2] 吴丹,王式功,尚可政.中国酸雨研究综述.干旱气象,2006,24(2):70-77.
[3] 齐泽民,王玄德,宋光煜.酸雨对植物影响的研究进展.世界科技研究与发展,2004,26(2):36-41.
[7] 张新民,柴发合,王淑兰,孙新章,韩梅.中国酸雨研究现状.环境科学研究,2010,23(5):527-532.
[8] 冯秋红,史作民,董莉莉,刘世荣.南北样带温带区栎属树种功能性状对气象因子的响应.生态学报,2010,30(21):5781-5789.
[9] 于顺利,马克平,徐存宝,金淑芳,宋晓兵,陈灵芝.环境水平下蒙古栎群落的物种多样性特征.生态学报,2004,24(12):2932-2939.
[14] 王仁卿,周光裕.山东植被.济南:山东科学技术出版社,2000:259-266.
[15] 房用,孟振农,朱宪珍.山东森林植物资源及应用.北京:中国国际广播出版社,2006:41-42.
[16] 刘焕彬,王新堂.山东省城市大气降水pH值观测结果及其初步分析.中国环境监测,2006,22(6):92-95.
[17] 甄文栋.山东省城市酸沉降状况及成因研究.济南:山东大学,2005.
[18] 迟竹萍.近45年山东夏季降水时空分布及变化趋势分析.高原气象,2009,28(1):220-226.
[19] 徐宗学,孟翠玲,赵芳芳.山东省近40 a来的气温和降水变化趋势分析.气象科学,2007,27(4):387-393.
[21] 尹燕东,裘立群,魏珉,杨凤娟,王秀峰.温室CO2施肥对黄瓜幼苗根系生长及分泌物和伤流液组成的影响.生态学报,2010,30(7):1860-1867.
[22] 沈波,王熹.两个亚种间杂交稻组合的根系生理活性.中国水稻科学,2002,16(2):146-150.
[23] 曹树青,邓志瑞,翟虎渠,唐运来,韩国标,张荣铣,盛生兰,龚红兵,杨图南.籼型杂交水稻根系伤流量及其衰退特性的配合力及杂种优势分析.中国水稻科学,2002,16(1):19-23.
[24] 时向东,刘艳芳,文志强,王卫武.植物根系伤流研究进展.安徽农业科学,2006,34(10):2043-2045.
[26] 武静莲,王淼,蔺菲,郝占庆,姬兰柱,刘亚琴.降水变化和种间竞争对红松和蒙古栎幼苗生长的影响.应用生态学报,2009,20(2):235-240.
[27] 王淼,李秋荣,郝占庆,董百丽.土壤水分变化对长白山主要树种蒙古栎幼树生长的影响.应用生态学报,2004,15(10):1765-1770.
[29] 徐飞,郭卫华,徐伟红,王仁卿.刺槐幼苗形态、生物量分配和光合特性对水分胁迫的响应.北京林业大学学报,2010,32(1):24-30.
[30] 宋轩,曾德慧,林鹤鸣,山口·达明,西崎·泰,山田·帕丽达.草炭和风化煤对水稻根系伤流量和养分吸收的影响.应用生态学报,2001,12(6):867-870.
[33] 单运峰.酸雨、大气污染与植物.北京:中国环境科学出版社,1993:153-155.
[35] 金清,江洪,余树全,金静,王艳红.酸雨胁迫对亚热带典型树种幼苗生长与光合作用的影响.生态学报,2009,29(6):3322-3327.
[37] 樊后保,黄玉梓,袁颖红,李燕燕,黄荣珍,樊海燕.森林生态系统碳循环对全球氮沉降的响应.生态学报,2007,27(7):2997-3009.
