不同光强对美国红栌光合特性的影响
2013-09-15张超
张 超
(太原大学外语师范学院,山西太原030012)
美国红栌(Cotinus coggygria‘Royal purple’)为漆树科黄栌属美国黄栌(C.coggygria)的变种,其叶色在春季为鲜红或紫红;入夏后,上部新生叶保持红紫色,下部叶逐渐转绿;入秋后叶片鲜红动人,一般作为园林建设中的彩叶植物。按前人对彩叶植物的定义进行分类[1-2],美国红栌属叶色多变、表现优秀的彩叶树木,也是北京等地的5大栽培树种之一[3]。关于美国红栌的快繁与栽培[4]、组织培养[5]方面的研究较多,而光照是影响彩叶植物叶色变化最重要的环境因素[6-7]。叶片是进行光合作用最主要的器官,其对生境条件的变化最为敏感[8]。目前,关于光照强度对彩叶植物叶片光合特性的研究报道还不多。
本试验是继美国红栌在不同光质下光合特性研究[9]之后,为进一步研究不同光强对其光合特性的影响所进行的,目的是为彩叶植物常规的园林养护工作和保证叶色正常表达提供一定的参考。
1 材料和方法
1.1 试验地概况
试验以山西农业大学林学院苗圃内2年生美国红栌为试材。试验地位于山西省中部,海拔790 m,属温带大陆性季风气候,春季多干旱风,夏季炎热,秋季凉爽,冬季寒冷,昼夜温差大,年均温9℃左右;1月份均温7℃左右,7月均温为23℃左右,年降水量540 mm,无霜期140~180 d。
1.2 试验设计
本试验采用单因素设计,3个处理(含对照),随机区组排列,3次重复。以市售黑色遮阳网分别进行覆盖处理:处理1.加盖1层遮阳网遮光,相对光强为60%;处理2.加盖2层遮阳网遮光,相对光强30%;对照(CK)不使用遮阳网,在无其他遮挡物的自然条件下受光,相对光强100%。每个处理选取3株,共27株,处理期为35 d。采用TES-1332A型数位式照度计测定光照强度。
1.3 光合特性的测定
采用美国CID公司生产的CI-301PS型便携式光合测定系统,使用开放气路进行美国红栌叶片光合特性的测定。分别于2007年6月10日,6月15日和6月20日进行。测定时间为7:00—18:00,选择生长于树冠东南方,由顶部向下第5个枝条上发育良好的功能叶3片,每隔1 h测定一次,每次测定读数3次,计算平均值。
光合测定系统直接输出的数据有:时间(t),叶面积(cm2),大气温度(Ta,℃),叶表面温度(T1,℃),光合有效辐射(PAR,μmol/(m2·s)),空气相对湿度(RH,%),细胞间隙CO2的体积分数(Ci,μL/L),净光合速率(Pn,μmol/(m2·s)),蒸腾速率(Tr1,mmol/(m2·s)),气孔导度(Gs,mmol/(m2·s))等生理指标。
2 结果与分析
2.1 不同光强下净光合速率的日变化
不同光强下处理35 d后,叶片的净光合速率测定结果如图1所示。

从图1可以看出,对照的净光合速率日变化曲线呈双峰曲线,出现了明显的“午休”现象,一天中的净光合速率高峰分别出现在11:00和15:00,峰值分别是4.07,3.88μmol/(m2·s);处理1叶片的净光合速率的日变化呈单峰曲线,高峰出现于13:00,峰值为4.9μmol/(m2·s),明显高于对照,且全天较长时间净光合速率都维持在较高的水平;处理2叶片的净光合速率日变化也呈单峰曲线,高峰出现在13:00,峰值为4.04μmol/(m2·s),低于处理1与对照的峰值。
2.2 影响光合作用的主要生理生态因子的日变化
2.2.1 光合有效辐射的日变化 不同光强下处理35 d后,光合有效辐射的测定结果如图2所示。
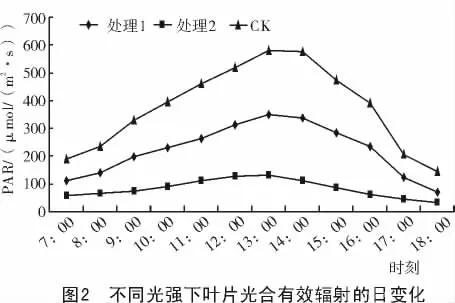
由图2可知,不同光强下的光合有效辐射日变化基本都呈抛物线型,但处理与对照的光合有效辐射强度差别极大。其中,处理与对照的峰值都出现在13:00左右,对照的峰值为580μmol/(m2·s),处理1的峰值为348μmol/(m2·s),而处理2的峰值为131.5μmol/(m2·s)。
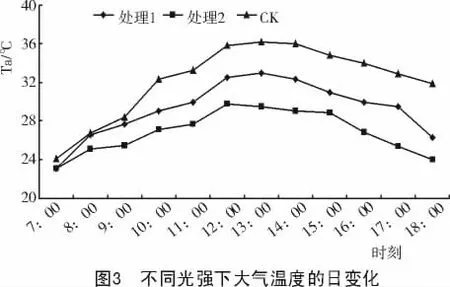
2.2.2 大气温度日变化 3种不同光强下的大气温度日变化测定结果如图3所示。
2.2.3 胞间CO2浓度的日变化 不同光强下处理35 d后,对美国红栌叶片胞间CO2浓度的日变化进行测定,结果如图4所示。
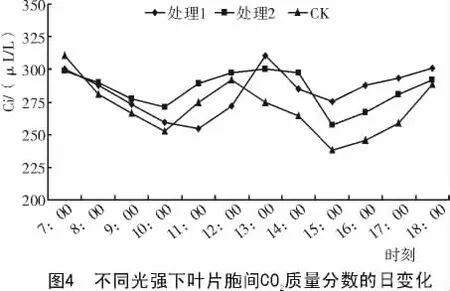
由图4可知,各处理与对照的美国红栌叶片的胞间CO2质量分数的日变化基本与其叶片的净光合速率日变化进程相反,这主要是因为净光合速率较大的时候,固定CO2较多,导致胞间CO2质量分数的降低;当中午光合降低的时候,胞间CO2质量分数的上升。
2.2.4 蒸腾速率的日变化 不同光强下处理35 d后,对美国红栌叶片蒸腾速率的日变化进行测定,结果如图5所示。
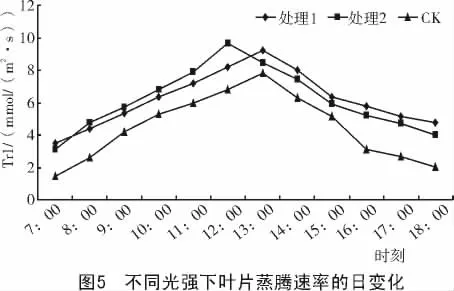
从图5可以看出,不同光强处理下的美国红栌叶片蒸腾速率均呈单峰曲线,处理与对照的蒸腾速率均从7:00开始随光照强度的增强和大气温度的上升呈逐渐上升趋势,12:00—13:00时达到最高峰,随后下降。日蒸腾速率表现为处理2>处理1>对照。
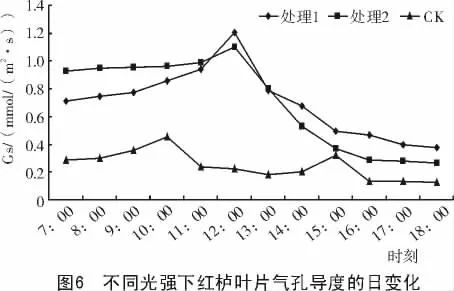
2.2.5 气孔导度的日变化 由图6可知,不同光强处理下美国红栌叶片的气孔导度日变化为单峰曲线,而对照的日变化为双峰曲线。在7:00—12:00,处理1和处理2的气孔导度绝对值均大大高于对照,有利于气孔张开,但是12:00以后,气孔导度出现明显的下降。一天中,对照的气孔导度始终最低,这是由于在自然条件下,光合有效辐射大,引起气孔部分关闭所造成的。
3 结论与建议
(1)在不同光强处理下,美国红栌叶片的净光合速率日变化不同。相对光强60%和30%处理的净光合速率的日变化呈单峰曲线。最高峰均出现在13:00左右,分别为4.9,4.04μmol/(m2·s);而对照的净光合速率日变化呈双峰曲线,有明显的“午休”现象,一天中的高峰分别出现在11:00和15:00,峰值分别是4.07,3.88μmol/(m2·s)。
(2)从不同光强下的净光合速率的日变化可以看出,处理1,2的净光合速率峰值均高于对照,这说明自然条件下的光照强度过大,可能会使处于光合诱导期的光合机构受到伤害,进而会影响植物的净光合速率,也会导致叶片蒸腾速率加快,使体内含水量降低,抑制植物生长[10],而适当的减弱光照强度会在一定程度上提高植物的净光合速率;同时也表明,光合作用可以在弱光下正常进行,而在光照最强时光合速率下降,说明美国红栌有较为明显的光抑制效应,是具有一定耐阴性的彩叶植物。
(3)美国红栌的气孔导度日变化趋势与其光合速率的日变化趋势类似,证明气孔导度与光合速率间存在明显的相关性,气孔作为植物叶片水汽和CO2进出的门户,控制着植物的光合作用和蒸腾作用[11-13],是影响光合速率的直接因素之一。
美国红栌在北方强日照地区栽培时,可适当地进行遮光处理,以促进植物的生长、保护植物免受日灼伤害。但是光照强度改变,也会导致叶片内叶绿素、花色素苷的含量和比例发生改变,如花色素苷含量下降、叶绿素b的含量上升等现象,均会导致叶色变化,降低其观赏性,故在彩叶植物上应用还需慎重。建议针对应用愈加广泛的彩叶植物[14-15],可开展不同品种、不同生长期的观测,使彩叶植物的生长发育和叶色表达能够达到平衡,为观叶苗木生产提供更详实的依据。
[1]杨潇怡,张力,冯岳东,等.彩叶植物资源及其园林应用的研究进展[J].天津农业科学,2011,17(2):138-141.
[2]崔向东,张海江,汪林华.彩叶植物与园林景观[J].河北林业科技,2005(4):173-175.
[3]吴晓星,李成,王翠香,等.美国红栌的栽植技术[J].山东林业科技,2005(2):51.
[4]肖芳.美国红栌组培快繁研究 [J].内蒙古林业调查设计,2012(6):31-32.
[5]田成利.美国红栌组培快繁体系的研究[D].合肥:安徽农业大学,2008.
[6]李红秋,刘石军.光强度和光照时间对色叶树种叶色变化的影响[J].植物研究,1998,18(2):194-205.
[7]于晓南,张启翔.彩叶植物多彩形成的研究进展[J].园艺学报,2000(S1):533-538.
[8]王耀芝,王勋陵,李蔚.荒漠化草原常见植物叶内部结构的观察[J].兰州大学学报,1983(3):87-96.
[9]张超.不同光质对美国红栌光合特性的影响[J].山西农业科学,2012,40(4):348-352.
[10]张锦春,赵明,张应昌,等.灌溉植被梭梭、白刺光合蒸腾特性及影响因素研究[J].西北植物学报,2005(1):70-76.
[11]于强,谢贤群,孙菽芬,等.植物光合生产力与冠层蒸散模拟研究进展[J].生态学报,1999,19(5):744-753.
[12]宋庆安,童方平,易霭琴,等.虎杖光合生理生态特性研究[J].中国农学通报,2006,22(12):71-76.
[13]许大全.光合作用效率 [M].上海:上海科学技术出版社,2002.
[14]申浩.彩叶植物在园林景观设计中的应用 [J].天津农业科学,2012,18(2):166-168.
[15]杨潇怡,张力,冯岳东,等.彩叶植物资源及其园林应用的研究进展[J].天津农业科学,2011,17(2):138-141.
