松嫩草地返青期芦苇形态及解剖结构对全球气候变化的响应
2013-09-15石连旋于文若郭继勋
石连旋,于文若,蒋 章,郭继勋
(1.东北师范大学草地科学研究所,吉林 长春 130024;2.吉林大学生命科学学院,吉林 长春 130012)
大气CO2浓度升高和其他温室气体排放导致全球气候变暖,预计21世纪末全球平均气温将增加1.8℃~4.0℃[1].同时,由于化石燃料燃烧和农业生产中氮肥的不合理使用,向大气排放的含氮化合物逐渐增加,引起大气氮沉降成比例增加.据估计,到2050年氮沉降量将增加到200亿t/a.全球气候变化的影响及适应研究不仅将成为今后一个时期内科学研究的重点,而且会成为国际社会关注的焦点[2-3].
植物外部形态和内部显微结构对气候变化的响应,是群落结构、群落生物量、生产力以及植物光合作用、呼吸作用、蒸腾作用等发生变化的原因和基础[4].目前,国内外对于气候变暖与氮素沉降的研究大部分集中在森林生态系统中,对草原生态系统的研究也多集中于对植物光合作用、呼吸作用、蒸腾作用等的影响,以及对群落结构、群落生物量及生产力、物种多样性、土壤及植物中矿质元素含量、土壤微生物量影响等方面[5-6],而全球气候变暖与氮素沉降对植物外部形态以及内部显微结构影响的研究鲜见报道.
本实验以松嫩草地的优势植物芦苇为研究材料,在野外自然状态下,人工模拟增温、施氮控制实验,观察、测定了植物外部形态以及内部显微结构的变化情况,探讨了返青期植物形态结构对于全球气温升高和氮素沉降的变化趋势,进而揭示了草原植物在形态结构方面对于全球气候变化做出的响应过程.
1 材料与方法
1.1 实验设计
在松嫩草地选取地势平坦、植被均匀、具有代表性的地段设置144m2样地,划分为12个4m×3m的小区,各小区四周间隔3m.其中6个小区进行增温模拟实验:采用红外线加热仪(infrared radiator,USA)实施增温处理,灯管为南北方向,距地面2.25m;地表增温幅度为(1.7±0.1)℃,温度处理保持终年实施;其他6个小区设为对照.同时,每个小区一半施氮(10g/m2),一半不施氮,氮肥采用分析纯硝酸氨,每年施肥一次.本研究共设置4实验处理,增温(W)、施氮(N)、增温+施氮(WN)、对照(CK),每个实验处理6次重复.实验设备于2004年安装,开始进行增温、施氮模拟处理实验.
1.2 取样方法及实验样品的前处理
于2010年和2011年的5月初芦苇返青期取样,每个相同处理的小区分别取芦苇5株,共取30株.为防止破坏样地土壤环境,只取芦苇地上部分即茎、叶.取样时,选择芦苇每个植株上发育成熟、位于中间节位的叶片,剪成长4~5mm、宽2~3mm的小片,放入标准FAA固定液(50%乙醇90mL+冰醋酸5mL+甲醛5mL)中固定保存[7].
1.3 植株外部形态测量
对芦苇外部形态特征(茎长、叶数、节数,叶长、叶宽)进行统计测量.其中茎长是芦苇最上端叶片的顶端到根茎分界处的长度;叶宽是植株成熟叶片最宽处的宽度.
1.4 显微切片制作
采用改进的植物石蜡切片方法制作显微切片[7-9].利用Motic BA400型(麦克奥迪公司生产)生物显微镜对叶的维管束大小(芦苇的维管束近似椭圆形,所以采用测量椭圆形的方法测量,指标包括椭圆形的长半轴、短半轴、面积、周长)以及气孔保卫细胞的长度、宽度、气孔下腔面积进行测量;周长的测量利用Motic Images 2000.
1.5 数据分析
采用Sigma Plot和SPSS 16.0(SPSS,Chicago,Illinois,USA)软件进行数据处理和图表制作.
2 实验结果
2.1 增温、施氮处理对芦苇高度和节数的影响
增温、施氮及增温+施氮处理均增加了芦苇返青期植株的高度(见图1),3种实验处理之间不存在显著差异,而与对照组间显著差异(P<0.05).3种实验处理均使芦苇植株节数平均值有所增加,与对照相比,施氮处理的增加效果显著(P<0.01),增温、增温+施氮的处理差异未达显著水平(P>0.05).
2.2 增温、施氮处理对芦苇叶长、叶宽和叶片数量的影响
松嫩草地芦苇叶片为长线形或披针形,叶鞘呈圆筒形,叶舌分布有茸毛,叶片排列成两行;叶片长6~12cm,宽1~2cm.3种实验处理对芦苇叶片和宽度有着一定的影响(见图1).增温、增温+施氮处理显著降低了芦苇的叶长(P<0.05);施氮处理与对照组相比较,也呈现降低的趋势,但是没有达到显著水平(P>0.05).3种处理都增加了芦苇叶片的宽度,尤其是施氮处理,叶片宽度显著高于其他实验组(P<0.05).3种处理与对照相比,均使芦苇叶片数量呈现增加的趋势,数据统计分析显示,增温处理与对照组之间并不存在显著性差异(P>0.05),施氮、增温+施氮处理与对照组间差异显著(P<0.05).
2.3 增温、施氮处理对芦苇维管束结构的影响
芦苇为禾本科植物,叶为等面叶,即叶肉中没有栅栏组织和海绵组织的分化.芦苇叶的横切面是由上、下表皮、叶肉以及维管束组成.表皮则是由表皮细胞、表皮毛、气孔器和泡状细胞组成;表皮外切向壁可被番红-固绿联合染料染成红色,证明表皮有木质化的加厚,角质化程度很高.在芦苇叶的解剖结构中,维管束呈一大一小相间排列,大维管束形态结构完整,木质部、韧皮部以及维管束鞘等各组分清晰可见,其维管束鞘细胞为1~2层,为不完全封闭的“花环型”结构,表现出C4植物的结构特点.
增温、施氮以及增温+施氮处理对芦苇叶片维管束结构具有较大的影响(见图2).3个实验处理组均增加了芦苇维管束长半轴的长度,与对照组间存在显著差异(P<0.01),其中施氮处理增加幅度最为显著(P<0.01).对叶片维管束短半轴长度的测量表明,施氮和增温+施氮处理增加了维管束短半轴的长度,尤其是施氮处理,达到了极显著水平(P<0.01);而增温处理下,维管束短半轴的长度没有显著变化.增温、施氮以及增温+施氮处理下,芦苇叶片维管束面积及周长的变化趋势基本一致(见表1).3种处理都增大了芦苇维管束的面积与周长,其影响与维管束长半轴长度的变化趋势一致,4个实验组之间存在显著差异(P<0.05),其中,增温、增温+施氮对维管束周长的影响未达到显著水平(P>0.05);而施氮处理对维管束面积与周长的影响均达到了显著水平(P<0.01).

图1 增温和施氮条件下芦苇形态的变化

表1 增温、施氮条件下芦苇维管束面积与周长变化

图2 增温和施氮条件下芦苇维管束变化
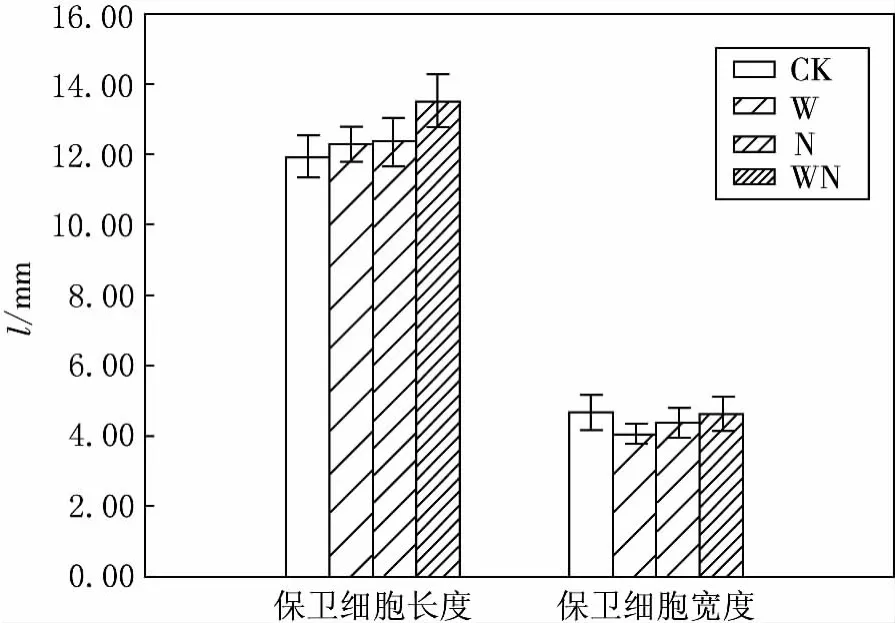
图3 增温和施氮条件下芦苇气孔保卫细胞的变化
2.4 增温、施氮处理对芦苇气孔保卫细胞特征的影响
芦苇叶片的上下表皮均有气孔分布,下表皮数量较多.气孔均由一对哑铃型保卫细胞和一对副卫细胞组成,与同样是松嫩草地优势植物的羊草叶片的气孔相比,芦苇的气孔下腔较小,面积及周长亦小于羊草.
增温与施氮处理对芦苇叶片气孔保卫细胞的长度与宽度均有不同程度影响(见图3),增温+施氮处理显著增加了芦苇气孔保卫细胞的长度,并达到了显著性水平(P<0.01);增温、施氮处理也显示出增加的趋势,但与对照相比差异并不显著(P>0.05).3种处理对芦苇叶片气孔保卫细胞宽度的影响,均表现出一定的降低现象,与对照相比较,差异同样没有达到显著水平(P>0.05).
在不同处理条件下,芦苇气孔下腔面积与周长的变化均呈现相似的变化趋势(见表2).施氮与增温+施氮处理,均增大了芦苇气孔下腔面积与周长,而且达到了显著水平(P<0.01);增温处理后,气孔下腔面积小于对照组,气孔下腔周长大于对照组,但均未达到显著水平(P>0.05).

表2 增温施氮条件下芦苇气孔下腔面积与周长变化
3 讨论
植物的外部形态对于气候变化的响应是一个相对较慢的过程,然而持续不断的气候变化也会使植物的外部形态产生一些变化.关于施氮对植物形态结构的影响,前人的研究表明,氮沉降量过大才会使氮本身促进植物生长的效果降低,而且氮沉降多与淹水偶联[10].我们的实验结果表明,施氮处理明显改变了植物的外部形态,显示出氮肥增大植株高度、增加植株节数、增多植株叶片数量、增大叶片宽度等促进植物生长作用的效果;同样,在维管束、气孔两个方面也表现出一定的促进生长的作用.这与实验区属温带半干旱季风气候,且样品均取自较为干旱和贫瘠的盐碱地带有着密切的关系.研究表明,对于某些土壤干旱贫瘠的地区,一定量的施氮处理,能够增加土壤肥力、促进植物生长,而不显现氮沉降抑制植物生长的现象.
温度和氮元素对于植物的生长和分布来说具有重要而复杂的作用.全球气候变化条件下,温度升高和氮素沉降严重影响着植物的新陈代谢和生长发育.许多模拟温度升高的实验研究显示:随着温度的升高,植物比叶面积(SLA)显著增加,栅栏组织和海绵组织细胞的层数及厚度、叶片厚度等则减少;而气孔器面积和气孔长宽指数减小,气孔导度降低[11-13].窦晶鑫等的研究表明,随着氮沉降量的增加,小叶章植株叶面积、分蘖数、株高以及生物量的积累逐渐增大[14];氮沉降对杉木人工林生长影响的研究表明,氮沉降初期,促进了林木胸径的生长,且氮沉降水平越高,林木胸径生长得越快,但随着氮沉降时间的延长,这种促进作用会慢慢减弱[15].松嫩草地植物在增温实验处理下,叶片长度降低、气孔下腔面积减少,而对维管束性状没有明显的改变;同时,增温处理还削弱了施氮处理的作用效果.增温施氮同时处理下,保卫细胞长度增加,这将使得水分蒸腾加快,不利于水分的保持.这些实验研究表明,长时间的全球气温的升高对草地植物生长将产生一定的限制作用.
[1]温玉璞,汤洁,邵志清.瓦里关山大气二氧化碳浓度变化及地表排放影响的研究[J].应用气象学报,1997(8):129-136.
[2]葛全胜,陈泮勤,方修琦,等.全球变化的区域适应研究:挑战与研究对策[J].地球科学进展,2004,19:516-524.
[3]陈宜瑜,陈泮勤,葛全胜,等.全球变化研究进展与展望[J].地学前缘,2002(9):11-18.
[4]涂利华,胡庭兴.模拟氮沉降对华西雨屏区苦竹林细根特性和土壤呼吸的影响[J].应用生态学报,2010,21:2472-2478.
[5]WU D X,WANG G X.Interaction of CO2enrichment and drought on growth,water use,and yield of broad bean(Vicia faba)[J].Environmental and Experimental Botany,1998,43:131-139.
[6]齐红,刘洋,郭继勋.松嫩草优势植物羊草和芦苇凋落物的分解对模拟增温及氮沉降的响应[J].东北师大学报:自然科学版,2012,44(4):114-119.
[7]穆春生.イネ品種の穂形質に関する発育形態学的研究[D].东京:东京大学,2001.
[8]迟丽华,宋凤斌.松嫩平原西部盐碱地区10种植物叶片结构特征及其生态适应性[J].生态环境,2006,15:1269-1273.
[9]洪德艳,张丽娟,王立军.东北地区芦苇营养器官比较解剖学研究[J].吉林农业大学学报,2008,30:161-165.
[10]谢迎新,张淑利,冯伟,等.大气氮素沉降研究进展[J].中国生态农业学报,2010(4):897-904.
[11]PYANKOY V I,GUNIN P D,TSOOG S.C4plant in the vegetation of Mongolia their natural occurrence and geographical distribution in relation to climate[J].Oecologia,2000,123:15-31.
[12]ALWARD R D,DETLING J K.Grassland vegetation changes and nocturnal global warming[J].Science,1999(1):229-231.
[13]李芳兰.包维楷.植物叶片形态解剖结构对环境变化的响应与适应[J].植物学通报,2005,22:118-127.
[14]窦晶鑫,刘景双,王洋,等.小叶章对氮沉降的生理生态响应[J].湿地科学,2009(7):40-46.
[15]KURZ W A,APPS M J,BEUKEMA S J.20th century carbon budget of Canadian forests[J].Tellus,1995,47:170-177.
