野生大豆根系形态对局部磷供应的响应及其对磷吸收的贡献
2013-09-12林雅茹唐宏亮申建波
林雅茹,唐宏亮,2,申建波*
(1中国农业大学资源与环境学院资源环境与粮食安全研究中心,北京100193;2河北大学生命科学学院,河北保定071002)
在自然生态系统中,土壤中的养分资源相对比较贫瘠,并呈现出高度的时空异质性分布[1-2]。为了获取土壤中的异质性养分资源,植物根系在形态和生理上表现出高度的可塑性反应。根系形态可塑性为养分资源的捕获提供了载体,而根系生理可塑性为养分资源的吸收提供了保障。植物根系在养分富集区内的增生能够增强养分离子的捕获和吸收能力[2-3]。
磷是植物必需的大量营养元素之一,对植物生长发育具有至关重要的作用,其在土壤中生物有效性低。由于磷在土壤中的移动性差,扩散速率低,使植物根系对磷的获取更多依赖于根系的生长和根形态的改变[4]。土壤施肥、农业生产活动、土壤生物化学过程等因素导致磷在土壤中呈现异质性分布。为了从土壤中的磷富集区捕获足够的磷来满足自身生长发育需要,植物通常会增加根系在磷富集区内的生长和扩大根系表面积来增强对磷的吸收[5]。对大豆、玉米、小麦等作物的研究发现,根系在土壤磷富集区中的分布越多,越有利于对磷的吸收[6-8];在拟南芥中,局部氮富集区侧根密度增大,而磷富集区无显著变化[9];在玉米中,局部磷富集区的大小能够明显影响地上部磷吸收,说明较小的磷富集区根系无法触及时不能被根系利用[4]。当低磷土壤中生长的根系进入磷富集区时,根系能够快速地做出反应,增强对磷的吸收能力[7]。由此可见,局部磷供应在很大程度上能够改变根系形态和生理吸收能力,引起植物地上生物量的积累和磷吸收的增加。
尽管前期的研究表明,局部磷施用对植物产生诸多有益的作用。然而不同植物种类的根系对局部磷供应的响应程度有所不同[5,9-14],且大多研究集中在农作物种类,关于野生种类的研究很少有报道。因此,探讨野生植物种类根系对局部磷供应的响应以及根形态各参数对磷吸收的贡献,对于深入理解植物根系对局部异质性养分的响应及其功能,提高养分的利用效率具有重要的理论与实践意义。
野生大豆(Glycine soja Sieb.et Zucc)为一年生草本植物,在我国各地广泛分布,抗逆性强,可用作牧草、绿肥和水土保持植物,是一种具有重要开发利用价值的植物种质资源,广泛应用于栽培大豆育种研究。为此,本研究以野生大豆为研究对象,通过土柱试验模拟磷局部供应,定量评价磷局部供应对野生大豆根系形态的影响以及这些根系形态参数对磷吸收的贡献。
1 材料与方法
1.1 土壤准备
供试土壤为中国农业大学昌平长期定位试验低磷钙质壤土,属于潮湿雏形土(aquic cambisols)。土壤基础肥力如下:有机碳7.09 g/kg,全氮 0.51 g/kg,总磷 0.45 g/kg,速效磷(Olsen-P)1.29 mg/kg,碱解氮89.68 mg/kg,土壤 pH(去离子水)为8.35。风干后的土壤过2 mm筛,除去植物残体,与洗净的河沙按重量比3∶1充分混合均匀,土壤容重为1.44 g/cm3。养分按下述两种方式加入纵向可拆卸的土柱中(直径10 cm,高50 cm,顶部留有5 cm空间用于浇水):1)磷均质供应(mg/kg,土)Ca(NO3)2·4H2O 1686、K2SO4133、MgSO4·7H2O 43.3、Fe-EDTA 5.5、CuSO4·5H2O 2.0、MnSO4·4H2O 6.6、CaCl2·6H2O 125、H3BO30.67、(NH4)6Mo7O24·4H2O 0.12 和ZnSO4·7H2O 10、KH2PO4878;2)磷局部供应 其他养分均质(mg/kg,土):磷(KH2PO4)按 2634 mg/kg,土施用量加入到中层,其他养分按上述养分含量均匀加入至土柱各层,即将磷均质供应处理土柱加入KH2PO4的量集中施入土柱的中层。每个土柱从上到下分为3层,每层高度15 cm,加土1.4 kg。试验共2个处理,每个处理4个重复,中层为磷局部供应区(磷浓度为600 mg/kg),相应磷均匀供应处理的中层为对照区(磷浓度200 mg/kg)(图1)。
1.2 植物培养
试验于2011年7~9月在中国农业大学日光玻璃温室进行。试验所用野生大豆种子采自北京市密云县石城镇白河岸边,后在中国农业大学上庄试验站繁殖备用。在培养之前,野生大豆种子先用浓硫酸(H2SO4,98%)浸泡30~40 min以除去种衣,处理之后的种子用蒸馏水反复冲洗,直至洗去残留的硫酸;将处理过的种子浸入饱和CaSO4溶液中吸胀12 h后,摆放于湿润的滤纸上,置于黑暗中萌发24 h。选取萌发一致的种子,于每个土柱播种5粒,待幼苗出土后,保留两株长势一致的幼苗直至收获,试验结果(生物量、磷含量和根形态)按两株植物的统计数据来表述。
1.3 根系形态参数的测定
播种45 d后,将土柱纵向拆卸,依刻度尺将土壤切分为上、中、下三层,分别过2 mm筛,筛分收集各土层的根系,冲洗干净后,装入自封袋,带回实验室在-20℃条件下冷冻保存。为了获得野生大豆不同根形态参数,将冻存的根系缓慢解冻后,在双面扫描仪(Epson Expression 1600 pro,Model EU -35,Japan)上进行根系扫描。扫描获得的图片直接通过WinRHIZO图像分析系统(WinRHIZO Pro2004b,version 5.0,Canada)获得总根长、根表面积、平均根直径等根形态参数。比根长通过总根长除以根干重来计算获得。

图1 磷局部供应对野生大豆根形态的影响Fig.1 Effect of phosphorus localized supply on the root morphology in wild soybean
1.4 生物量及植株磷含量的测定
收获植株后,将地上部在105℃下杀青30 min,在70℃烘干48 h至恒重后称重(g)。地下部分则在根系扫描结束后,烘干至恒重后称重(g)。将烘干的地上和地下部分研磨成粉末(<0.5 mm)后,用6∶1 HNO3-HClO4消煮,消煮液的磷浓度用钒钼黄比色法测定[15]。
1.5 数据分析
采用SPSS 13.0 for Windows(September 2004,SPSS Inc.,USA)对数据进行单变量和多变量统计分析。独立样本t检验单个指标在磷供应方式之间的差异显著性;主成分分析和多变量方差分析(MANOVA)用于比较根形态参数在磷供应方式之间的总体差异显著性;采用SigmaPlot 10.0对磷吸收和根长进行一元线性回归拟合;磷吸收和根形态指标之间的关系采用Simca-P 11.5软件进行偏最小二乘法回归分析。
2 结果与分析
2.1 局部磷供应对野生大豆根系形态、磷吸收及生物量的影响
从表1可以看出,磷局部供应对野生大豆的根系形态各参数(总根长、比根长、根表面积和根直径)和地上磷吸收有显著的影响(P<0.05),对苗干重的影响极显著(P<0.01),但没有显著影响根干重和根冠比。在磷供应总量相同的情况下,与磷均匀供应相比,磷局部供应使总根长增加了80.51%,比根长增加了32.62%,根表面积扩大了70.65%,根直径减小了27.56%,说明磷局部供应能够在促进野生大豆总根长增加的同时,刺激了许多细根的产生,使根系表面积扩大,促进了对磷的吸收(43.15%),最终增加了地上生物量(72.00%)。这些结果表明,野生大豆根系形态对磷的局部供应具有较强的可塑性反应,这种根的可塑性很可能源于磷局部供应区根系形态参数的改变以及对磷吸收能力的增强。
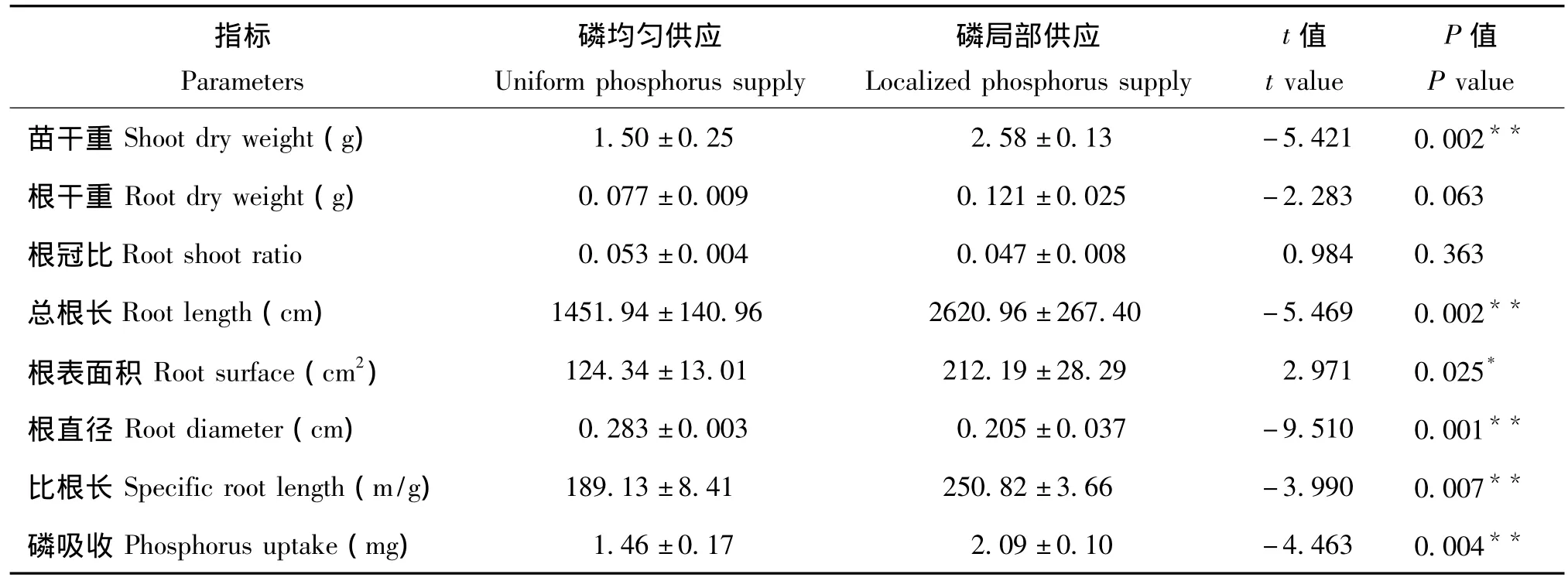
表1 局部磷供应对野生大豆根系形态、磷吸收及生物量的影响Table 1 Effect of localized phosphorus supply on root morphology,P uptake and biomass in wild soybean
为此,本研究比较了磷局部供应区和对照区根形态参数的差异。由表2可以看出,除了根干重外,磷局部供应区的总根长、比根长、根表面积显著高于对照区,分别增加了 123.93%、22.45%、115.64%,而平均根直径则明显低于对照区(P<0.05),相应减少了5.34%。通过比较表1和表2中的数据,可以计算出局部磷供应区域根形态参数占整个根系的比例。根干重、总根长、根表面积分别占总根系的37.19%、68.10%、66.78%,充分说明局部磷供应区内野生大豆根系大量增生。

表2 局部磷供应区和对照区野生大豆根系形态参数的比较Table 2 Comparison of root morphological parameters between localized and uniform phosphorus supply in wild soybean
为了研究局部磷供应对野生大豆根系形态的总效应,采用主成分分析方法对根干重、总根长、比根长、根表面积和根直径5个根形态参数进行了主成分提取。从图2的排序图可以看出,提取的前两个主成分累积贡献率为97.39%,其中主成分1(PC1)贡献率为 76.02%,主成分 2(PC2)贡献率为21.37%。
由于各根形态参数权重不同,其在所提取主成分中的载荷量也是不同的。由表3可以看出,在第一主成分中,根干重、总根长、比根长和根表面积有较高的载荷(权重在0.815~0.989之间),根直径则有较低的载荷量(权重为0.647),主成分1可以明显将磷均匀供应与磷局部供应两个处理分开;在第二主成分中,根直径有较高载荷量(权重为0.749),根干重、总根长、比根长和根表面积有较低的载荷量(权重在0.126~0.568之间),由于根直径在主成分1和主成分2中的载荷量相当,因此两个处理之间在主成分2方向没有显著差异。采用多变量方差分析进一步对5个根形态参数的分析结果表明,野生大豆根形态在局部磷供应和磷均匀供应之间有明显的差异(Wilk’s Lambda=0.004,F=179.932,P< 0.001)。

图2 磷局部供应对野生大豆根形态影响的主成分分析Fig.2 The principal component analysis of root morphological parameters as affected by the localized phosphorus supply in wild soybean

表3 主成分载荷矩阵Table 3 Principal component loading matrix
2.2 野生大豆根系形态与植株磷吸收的关系
上述研究结果表明,野生大豆根系在局部磷供应区大量增生(图1和表2),这也许解释了地上磷吸收的增加。为了研究根形态参数与磷吸收的关系,对总根长、比根长、根表面积与磷吸收之间进行了回归分析。由图3可以看出,总根长、比根长、根表面积分别解释了81.07%、78.81%、76.79%的地上磷吸收,与磷吸收呈现明显的正相关关系(P<0.05)。随着根系的伸长,根系对磷的吸收也增加。由此可见,局部磷供应区根形态的改变对野生大豆磷吸收有重要影响。然而,由于根形态参数之间存在明显的互相关,存在多重共线性问题,从而掩盖了根形态参数(解释变量)对因变量(磷吸收)的解释。
因此,本研究进一步对根形态参数[根干重(x1)、总根长(x2)、根表面积(x3)、根直径(x4)、比根长(x5)]与磷吸收(y)的关系进行偏最小二乘(PLS)回归分析,并对各个指标的相对重要性进行了评价。结果表明,偏最小二乘回归自变量和因变量之间的交叉有效性为0.816(Q2),由根形态参数提取的主成分(t1)在解释了自身75.9%信息的同时,也解释了磷吸收(u1)85.9%的信息。两者之间表现出良好的相关性(R2=0.859、P <0.01),说明由偏最小二乘法建立的根形态参数与磷吸收关系的模型拟合度较高,可进行相关预测,回归方程为:Y=0.771+1.943X1+1.336 ×10-4X2+1.616 ×10-3X3-1.274X4+2.649 ×10-3X5。偏最小二乘回归在预测磷吸收的同时,也给出了根形态各参数对磷吸收作用力大小的变量投影重要性(VIP)评价。

图3 总根长、比根长和根表面积与磷吸收的一元线性回归Fig.3 The linear regression between total root length,specific root length,root surface area with P uptake
由图4可以看出,5个根形态参数在偏最小二乘回归模型中变量投影重要性(VIP)值均超过0.8,说明这5个指标均对地上磷吸收具有重要的影响。其中总根长、根表面积和比根长的VIP值超过1,对地上磷吸收起着明显重要的作用。为了进一步检验根形态参数对磷吸收的定量贡献,根据所建立的回归方程,对其中四个自变量取定值,将剩余的1个自变量值改变(提高或降低)10%,计算相应的磷吸收量比原始数值的增加量。从表4的计算结果中可以看出,根干重、总根长、根表面积、比根长10%的增加量分别引起地上磷吸收1.08%、1.53%、1.53%、3.28%的增加,而平均根直径10%的减小可以导致地上磷吸收1.75%的增加。相比之下,比根长对植株磷吸收的贡献最大,这也暗示根长增加的同时,根直径的减小可以明显增加野生大豆根系对磷的吸收。
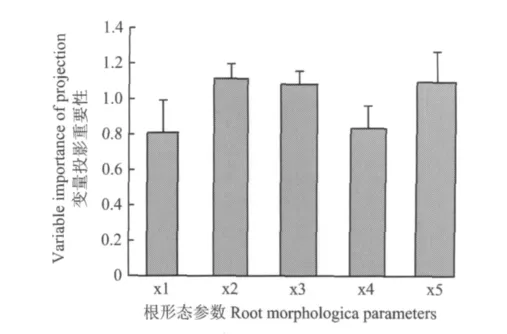
图4 野生大豆根形态参数对磷吸收影响的重要性评价Fig.4 Variable importance of projection(VIP)of root morphological parameters for P uptake in wild soybean

表4 野生大豆根形态参数改变对磷吸收的相对贡献Table 4 Relative contribution of root morphological parameters to P uptake
3 讨论
土壤中的养分呈现高度的时空异质性分布[2,16],植物根系的模块化生长和可塑性促进了植物根系对土壤中这些异质性养分资源的获取[2,17]。当异质性养分资源为磷酸盐时,一些植物(如大麦,玉米,白羽扇豆,水稻,鹰嘴豆,大豆)表现出较强的可塑性反应[4,14,18-21],然而另一些植物种类(如小麦,蚕豆,紫羊茅,香黄花茅,甘松茅)表现出较弱的可塑性反应[3,10,22]。与浅根型的栽培大豆相比,野生大豆为深根型的植物[20,23],其对养分异质性的响应很可能更为显著。在本研究中,野生大豆在局部磷供应区的总根长、比根长和根表面积比对照分别提高了123.93%、22.45%和115.64%(图1和表2),表明野生大豆具有很强的根系形态可塑性反应,其对土壤异质性磷素养分资源的获取很可能是根形态主导的适应策略。需要指出的是,本研究中磷均匀供应和局部供应两个处理的养分总量相同,仅仅改变了养分的分布模式;在对照区,磷的供应强度为200 mg/kg,在局部磷供应区,磷的浓度达到600 mg/kg,因此局部供应区较高的磷浓度以及野生大豆表现出高度的可塑性和觅食特性是导致野生大豆根系在局部磷供应区大量增生的原因。这同时也提出了一个问题,局部磷供应区磷浓度究竟达到多大时才能诱导野生大豆根系的增生?这需要进一步研究来确定。通过本研究和前期的大量研究可以看出,植物根系对异质性养分资源的可塑性反应依赖于植物种类、养分资源组成和供应强度[2-3,10,18,22]。
本研究中,尽管磷局部供应没有显著影响野生大豆根系总干重和根冠比(表1),但明显提高了总根长,比根长,根表面积和减小了根系的平均直径(表1和图2)。这些结果说明野生大豆在养分供应模式改变的条件下,通过优化根系空间分布和改变根系形态,最小化地上部分碳向地下的分配,以此保持相对稳定的根冠比例,高效获得养分资源,暗示野生大豆能够很好地协调地上部分和地下部分的关系。
植物根系对养分的吸收主要依赖于质流和扩散,而通过根系截获获得的养分量往往很少。磷在土壤中的移动性差,且易被吸附和固定,因此植物对磷的吸收主要靠扩散和根系截获[24]。由于土壤施肥、农业生产活动、土壤生物化学过程等因素导致磷在土壤中呈现异质性分布,植物往往通过增加根系在磷富集区的分配比例、根系长度和分枝数量来占据更多的土壤体积,提高对磷的获取能力[2,4,14,18,20,25-26]。Robison[11]发现,土壤中养分离子移动性低的情况下,根系快速生长并在养分富集区域增生的能力才能显著发挥。对磷素这种在土壤中有效性低、移动性很小的营养元素,其获取主要取决于形态学特性(如根长、根表面积等),而并不依赖于根系生理学特性[4]。在本研究中,野生大豆增加了根系在磷局部供应区的分配(表1和表2),占据了更多的土壤体积,增加了与移动性差的土壤磷素的接触面积,补偿了其余根系由于无法获取磷对植物生长消耗的影响,最终增加了根系对磷的吸收,说明野生大豆在很大程度上依赖于根系形态特性的改变来增加对磷的获取。由此可以推测,野生大豆的根形态参数与地上磷吸收之间存在一定的相关性。对野生大豆总根长、比根长和根表面积与地上磷吸收量进行的一元线性回归分析结果表明,总根长、比根长和根表面积与地上磷含量存在明显的正相关关系,其分别解释了81.07%、78.81%、76.79%的地上磷吸收(图3),说明野生大豆根系在局部磷供应区的大量增生对地上磷吸收具有重要的作用。然而由于根形态变量之间高度的相关关系,存在多重共线性问题,从而掩盖了根形态参数对磷吸收的解释能力。为了解释根形态参数对野生大豆地上磷吸收的贡献,本研究也采用了偏最小二乘回归分析研究了野生大豆5个根形态参数与地上磷的关系及其对磷吸收的贡献,结果表明,总根长,比根长和根表面积对磷吸收有重要的贡献(图4),其中比根长的贡献相对要大,10%的总根长、比根长、根表面积的增加可以分别引起1.53%、3.28%、1.53%磷吸收的增加(表4),这与一元线性回归的结果稍有所不同,表明野生大豆对土壤异质性磷的获取采取以根形态改变为主导的适应策略。以上研究进展为通过优化养分的施用方式定向调控根系的生长,减少磷肥的大量施用,提高作物对养分的利用效率提供了重要的科学依据。
[1] 王庆成,程云环.土壤养分空间异质性与植物根系的觅食关系[J].应用生态学报,2004,15(6):1063-1068.Wang Q C,Cheng Y H.Response of fine roots to soil nutrient spatial heterogeneity[J].Chin.J.Appl.Ecol.,2004,15(6):1063-1068.
[2] Hodge A.The plastic plant:root responses to heterogeneous supplies of nutrients[J].New Phytol.,2004,162:9-24.
[3] 荆晶莹.局部养分供应对植物根系生长和养分吸收的影响与根际调控机制[D].北京:中国农业大学博士论文,2011.Jing J Y.Effect of localized supply of nutrients on plant root growth and nutrient uptake and the mechanisms of rhizosphere management[D].Beijing:PhD dissertation,China Agricultural University,2011.
[4] Katsuya Y, TakashiK. Rootmorphologicalplasticity for heterogeneous phosphorus supply in Zea mays L[J].Plant Prod.Sci.,2005,8(4):427-432.
[5] Sun H G,Zhang F S,Li L et al.The morphological changes of wheat genotypes as affected by the levels of localized phosphate supply[J].Plant Soil,2002,245:233-238.
[6] Yao J P,Barber S A.Effect of one phosphorus rate placed in different soil volumes on P uptake and growth of wheat[J].Soil Sci.Soc.Am.J.,1985,49:152-155.
[7] Jackson R B,Manwaring J H,Caldwell M M.Rapid physiological adjustment of roots to localized soil enrichment[J].Nature,1990,344:58-60.
[8] 严小龙,廖红,戈振扬,等.植物根构型特性与磷吸收效率[J].植物学通报.2000,17(6):511-519.Yan X L,Liao H,Ge Z Y et al.Root architectural characteristics and phosphorus acquisition efficiency in plants[J].Chin.Bull.Bot.,2000,17(6):511-519.
[9] Birgit I L,Lisa C W,Alastair H F et al.Nitrate and phosphate availability and distribution have different effects on root system architecture of arabidopsis[J].Plant J.,2002,29(6),751-760.
[10] Robinson D.The response of plants to non-uniform supplies of nutrients[J].New Phytol.,1994,127:635-674.
[11] Robinson D.Resource capture by localized root proliferation:why do plants bother?[J].Ann.Bot.,1996,77:179-185.
[12] Brouder S M,Cassman K G.Cotton root and shoot response to localized supply of nitrate,phosphate and potassium:Split- pot studies with nutrient solution and vermiculitic soil[J].Plant Soil,1994,161:179-193.
[13] Terry J R,Zed R,Ma Q F et al.Crop species differ in root plasticity response to localized P supply[J].J.Plant Nutr.Soil Sci.,2009,172:360-368.
[14] Li H G,Shen J B,Zhang F S et al.Localized application of soil organic matter shifts distribution of cluster roots of white lupin in the soil profile due to localized release of phosphorus[J].Ann.Bot.,2010,105:585-593.
[15] 鲍士旦.土壤农化分析(第三版)[M].北京:中国农业出版社,2000.Bao S D.Soil and agro-chemistry analysis(3rd Ed.)[M].Beijing:China Agriculture Press,2000.
[16] Hodge A.Roots:the acquisition of water and nutrients from the heterogeneous soil environment[J].Prog.Bot.,2010,71:307-337.
[17] Hutchings M J,de Kroon H.Foraging in plants:the role of morphological plasticity in resource acquisition[J].Adv.Ecol.Res.,1994,25:159-238.
[18] Drew M C.Comparison of the effects of a localized supply of phosphate,nitrate,ammonium and potassium on the growth of the seminal root system,and the shoot,in barley[J].New Phytol.,1975,75:479-490.
[19] He Y,Liao H,Yan X L.Localized supply of phosphorus induces root morphological and architectural changes of rice in split and stratified soil cultures[J].Plant Soil,2003,248:247-256.
[20] 赵静,付家兵,廖红,等.大豆磷效率应用核心种质的根构型性状评价[J].科学通报,2004,49(13):1249-1257.Zhao J,Fu J B,Liao H et al.Application of phosphorus efficiency of soybean core collection of root architecture traits evaluation[J].Chin.Sci.Bull.,2004,49(13):1249-1257.
[21] Jing J Y,Rui Y K,Zhang F S et al.Localized application of phosphorus and ammonium improves growth of maize seedlings by stimulating root proliferation and rhizosphere acidification[J].Field Crop Res.,2010,119:355-364.
[22] Bart F,de Kroon H,Frank B.Root morphological plasticity and nutrient acquisition of perennial grass species from habitats of different nutrient availability[J].Oecologia,1998,115:351-358.
[23] 庄炳昌.中国野生大豆生物学研究[M].北京:科学出版社.1999.Zhuang B C.Biological research on wild soybean in China[M].Beijing:Science Press,1999.
[24] Barber S A.Soil nutrient bioavailability.A mechanistic approach[M].New York,USA:John Wiley.1984.
[25] Jackson R B,Caldwell M M.Integrating resource heterogeneity and plant plasticity:modeling nitrate and phosphate uptake in a patchy soil environment[J].J.Ecol.,1996,84:891-903.
[26] 金剑,王光华.不同施磷量对大豆苗期根系形态性状的影响[J].大豆科学,2006,25(4):360-364.Jin J,Wang G H.Effect of different phosphorus regimes on root morphological characteristics of soybean seedling[J].Soybean Sci.,2006,25(4):360-364.
