内蒙古羊草群落、功能群、物种变化及其与气候的关系
2013-09-11谭丽萍周广胜
谭丽萍,周广胜
(1.中国科学院植物研究所植被与环境变化国家重点实验室,北京 100093;2.中国气象科学研究院,北京100081;3.中国科学院研究生院,北京 100049)
全球气候变化及其影响已经引起了各国政府、科学界与公众的强烈关注。据预测,CO2浓度倍增后的全球平均地面气温可能上升1.9—5.2℃,且在中高纬度地区升温最为显著;全球年降水量将增加[1]。内蒙古草原是全球半干旱地区典型草原生态系统之一,其植被主要为真旱生与广旱生多年生丛生禾草[2],且该区域是全球气候(降水和温度)变化幅度较大的地区之一[2-3]。研究表明[4],1951—2009年我国及各地区增温显著,其中以内蒙古地区增温速率最快;冬季气温的变幅最大,平均气温上升了2.3℃,其中内蒙古地区以0.63℃/10 a的极显著速率位列第一;1951—2009年全国降水距平不如平均气温变化剧烈,但内蒙古地区四季降水量均呈明显的下降趋势。内蒙古锡林郭勒草原自然保护区占内蒙古天然草地总面积的1/3,生态系统非常脆弱[5],气候变化必将严重影响草原生态系统。研究表明,在不同生态系统中,气候变化对草地生态系统的影响尤为显著[6-9]。生物量作为生态系统功能的重要表现,是草地生态系统物质循环和能量流动不可缺少的环节。研究表明,降水对草地的影响存在明显的时间性和空间性,不同的研究地点、研究时段所得出的结论存在差异[10-12]。
群落变化过程中可能存在多个功能群、物种之间的补偿作用,功能相似而环境敏感性不同的物种稳定了生态系统的过程[13-15]。研究指出,种群多度的长期变异性依赖于物种对环境波动的敏感性和种间相互作用对环境波动的放大效应[16]。物种对环境波动响应的多样性增加可使群落的变异性降低,稳定性增强。为此,本研究试图基于1981—1994年内蒙古羊草草原的群落学调查数据和同期气象资料,分析羊草(Leymus chinensis)群落、功能群与主要物种变化及其生物量与气候的关系,揭示内蒙古典型草原对气候变化的响应机制及适应规律,为半干旱典型草原的可持续发展与管理提供依据。
1 研究区域概况和研究方法
1.1 研究区域概况
研究区域为内蒙古锡林郭勒草原自然保护区,位于内蒙古高原中部与大兴安岭南段西侧山地遥相接触的地带,在自然地理上属于温带内蒙古草原区典型草原栗钙土亚区。地理位置为43°26'—44°39'N,115°32'—117°12'E。研究区域地势东南高西北低,属大陆性温带气候,冬季寒冷干燥,夏季温暖湿润,年均温为-1.4—2.0℃,年降水量为240—360 mm,该地区生长季干燥指数在2以上,3—5月份常有大风。草原是以多年生根茎禾草羊草为主要优势种的温带典型草原,植物群落通常在每年的4月中旬开始萌发,4月下旬返青,5月初分蘖拔节,6月下旬抽穗,7月初开花,8月上旬种子成熟,其中6—8月份是其旺盛生长时期,9月下旬至10月日均温低于0℃以后,其地上部分停止生长,逐渐枯黄,进入休眠期,生长期长达150—160 d左右[17-18]。
1.2 研究方法
该样地[19-20]自1979年围封后未加利用,本研究资料为1981—1994年在中国科学院内蒙古草原生态系统定位研究站的羊草草原样地观测数据[21]。每年5月中旬到9月中旬,每半个月做1次植被调查,包括20个样方的植物种类、分种密度、频度、分种地上生物量。样方面积为1 m×1 m。同期气象资料(月尺度上的降水、温度)由锡林浩特市气象站提供[20]。
(1)变异系数与多样性指数

多样性指数计算如下:
Shannon-Wiener多样性指数
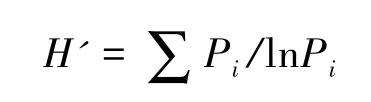
Simpson优势度指数
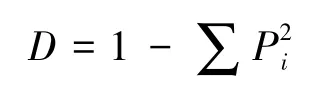
Margalef丰富度指数

式中,Pi=Ni/N,Pi为第i个物种的个体(或生物量、重要值等)所占样方总个体数(或生物量、重要值等)比值,S为物种数目[22-23]。本研究依据个体数进行多样性计算,生物多样性指数计算运用Biodiversity Pro软件,相关数据分析运用Excel 2007,SPSS16.0软件,Sigmaplot10.0作图软件。
(2)植物功能群划分
根据羊草草原植物的生活型和水分生态类型组成进行功能群的划分。生活型功能群包括多年生草本,灌木,一二年生草本;生态型功能群主要包括旱生植物和中生植物。
(3)Mann-Kendall方法[24-25]可以用于变量的趋势检验及突变分析,具体方法如下。
假设所检验数据序列x1,x2,…,xnn≥10
( ),Mann-Kendall统计量(Sk)计算如下:

式中,sign xj-xi

趋势变化由式(2)计算的UFk检验值判断。时,变化趋势不显著;反之,变化趋势显著。若UFk>0,表明序列具有上升趋势;UFk<0,则表明序列具有下降趋势。
(4)多元逐步回归分析方法[26]是在多元线性回归分析基础上派生的一种算法,它从大量可供选择的变量中选择建立回归方程的重要变量。在逐个因子选入回归方程的过程中,如果发现先前被引入的因子在气候由于某些因子的引入而失去其重要性时,可以在回归方程中随时予以剔除,直到最后被选入的因子对因变量都有显著的影响为止。
2 结果与分析
2.1 羊草群落的结构特征
样方调查记录表明,羊草群落以多年生根茎禾草羊草((Leymus chinensis)和多年生丛生禾草大针茅(Stipa grandis)为建群种和优势种,1981—1994年间共出现了93种植物,分属于28科66属。其中,禾本科(10属12种),豆科(6属7种),菊科(8属12种),藜科(4属7种),百合科(3属9种),蔷薇科(3属9种)这6个科的物种数占群落物种数的60.2%,且这些科多年间一直存在于羊草群落,并是主要组成物种。
在93种植物中,多年生植物有79种,占总物种数的84.9%,包含多年生灌木半灌木4种;旱生植物占总种数的74.2%,而中生植物24个,占总种数的25.9%。在6大科56个物种中,仅有7个物种属于中生。这
式中,UF1=0,E(Sk)和Var(Sk)分别是Sk的均值和方差。
对计算的UFk值做图,将得到对应的变化曲线。给定显著水平α,在正态分布表查临界值UFα/2。若表明,羊草群落以多年生/旱生植物为主。
2.2 羊草群落结构与生物量变化
(1)1981—1994年间,8月中旬羊草群落的多样性(Shannon-wiener)指数、优势度指数、丰富度指数、平均高度、生物量均呈波动变化,没有显著的升高或降低的趋势,变异系数分别为9.6%、20.35%、14.95%、16.81%和26.1%。14a间羊草群落结构和功能变异性相对较小,主要变化体现在生物量方面。
进一步对生长季中其他月份的多样性、群落高度、群落生物量的年际间变异分析表明(表1):14a间生长季各月均以生物量变异最高,其次为优势度指数,变异性较低的有丰富度指数和多样性指数;各指数在5月的年际间变异明显高于其他月份,其中群落生物量的变异从5月到9月依次降低;不同月份的群落高度年际间变异相差不大,丰富度指数的变异在8月份明显高于6、7和9月。

表1 1981—1994年不同月份羊草群落的多样性、高度、生物量的年际间变异Table 1 Annual variation of diversity indices,height,aboveground biomass of Leymus Chinesis community during the growing season from 1981 to 1994
(2)植物生活型功能群组成中(以8月中旬数据进行分析),多年生草本,一二年生生草本,灌木生物量的变异系数分别为27.9%、64%和44.2%。一二年生草本生物量年际间变异系数最高,灌木生物量年际间变异数次之,分别为群落高度年际间变异系数的2.45倍和1.69倍。
在群落植物的生态类功能群中,旱生植物占很大成分,其生物量的年际间变异系数为23.1%,而中生植物生物量年际间变异极其显著,变异系数达100%,为群落生物量年际间变异系数的3.83倍。这反映出气候变化导致的草原水分亏缺使得草原中生植物发生重大变化。
相关分析表明,羊草群落生物量及其年度动态与多年生草本的生物量、旱生植物生物量有着极显著的正相关关系,相关系数分别为0.980(P<0.005)和0.986(P<0.005)。这表明,羊草群落生物量的变异主要由多年生草本及旱生植物决定,反映出了羊草群落生物量的稳定性。
1981—1994年观测数据表明,围封样地中羊草平均相对地上生物量为27.9%,大针茅平均相对地上生物量为21.9%,两者绝对生物量之和与群落地上生物量的相关性为0.882(P<0.005),反映出羊草和大针茅的优势地位。进一步分析羊草与大针茅植物绝对生物量变化的相关性(图1)表明,相关系数达-0.767(P<0.001),反映出羊草与大针茅存在生物量补偿作用,即在羊草地上生物量降低时,大针茅地上生物量增加,两者之间存在显著的补偿性,这种优势种之间的补偿性很好地维持了羊草群落地上生物量的稳定性。

图1 羊草和大针茅植物的生物量变化Fig.1 Variation of aboveground biomass of Leymus chinensisand Stipa grandis plants
这种波动性不仅发生在羊草-大针茅草原的优势种与亚优势种之间,还存在于群落的其他组成物种之间。1981—1994年的8月中旬共出现77种植物,其中前37种植物的平均相对生物量为94%,变异系数从32%到202%,仅4个物种的变异系数在50%以下,物种生物量的变异远高于群落及功能群生物量的变异。对这37种植物的生物量进行pearson相关分析表明,植物种对之间没有相关关系的占90.4%,也就是大部分物种的生产力之间不存在显著的关系;存在正相关关系的物种有58种对,占总对数的8.7%;负相关关系仅仅在少数种之间存在,共6种对,占总对数的0.9%。这些种对间可能存在生物量升降的互补关系,共包含冰草(Agropyron)、黄囊苔草(Carex korshinskyi)、早熟禾(Poa sphondy lodes)等13个物种,且这种负相关关系在生活型上主要存在于多年生草本,生态型上主要存在于旱生植物中。这些非优势种植物虽然相对生物量较低,但是其地上生物量间的相互补充与优势种间的补偿作用共同减少群落生物量的强烈波动,使羊草群落更为稳定。
2.3 群落结构和生物量的关系
相关分析表明,群落生物量和多样性指数、优势度指数、丰富度指数之间pearson相关系数分别为-0.354、0.350、0.038,且都不显著,而与群落的优势种(羊草、大针茅)的生物量之和的变化有着密切的关系。因此群落的稳定性并非主要受多样性的影响,而群落优势度和羊草大针茅的生物量之和有很高的正相关关系,相关系数为 0.644(P<0.01)。
2.4 羊草群落生物量与气候的关系
研究表明[27],1982—1998年中国科学院内蒙古草原生态系统定位研究站所在地区的年均温、>0℃积温均呈显著的增加趋势,且主要表现为冬季增温,冬季增温使该地区的春季干旱进一步加剧;降水虽有增加,但波动较大,除1986、1990、1992年水分略有盈余外,其余年份的水分亏缺都较严重。利用1981—1994年内蒙古锡林浩特市气象站的气象数据,结合Mann-Kendall方法[24-25]分析表明[28],除1986、1987年外,上年10月至当年8月的均温呈不显著的增加趋势,而期间的年累积降水在1986年前呈不显著的减少趋势,1986年开始呈不显著的增加趋势(图2);1月至8月的降水和生长季(4—8月)降水与上年10月至当年8月的累积降水变化趋势基本一致,但1986—1988年的1月至8月降水和生长季(4—8月)降水呈下降趋势,而上年10月至当年8月的累积降水呈增长趋势,表明上年10月至当年1月降水在1986—1988年呈增长趋势;1—8月均温的变化趋势和上年10月至当年8月均温变化趋势一致,但前者相对剧烈,生长季(4—8月)均温在1983—1989年先呈不显著的上升趋势,而后开始下降。

图2 不同时段均温和降水的M-K法趋势分析Fig.2 Mann-Kendall measure curve of precipitation and average temperature in different periods Ce0.05为 α=0.05的显著性水平
围栏样地的羊草草地地上生物量变化主要由气候决定。多元逐步回归分析表明(表2):上年10月至当年8月均温对群落总生物量和旱生植物生物量有显著的负效应,中生植物生物量与4月至8月的降水呈显著正相关;多年生草本生物量与上年10月至当年8月降水呈显著正相关,而灌木和一二年生植物与气候因子不存在显著关系。研究表明[12],群落地上初级生产力与上年10月至当年8月累积降水呈显著的二次曲线关系。本研究表明,群落生物量与上年10月至当年8月均温呈显著的负相关,说明羊草群落的生物量由水热条件共同决定。

表2 羊草群落及功能群的生物量与气候的关系Table 2 Relationships between climate factors and the biomass of Leymus chinensis community and functional groups
3 讨论
3.1 羊草群落变化
1981—1994年羊草群落结构和生物量变异性相对较小,与群落多样性及高度的变化相比,生物量变化较大。因此,气候影响更多地反映在羊草生长特性的变化方面,而非物种数量的变化。当温度与降水格局发生变化时,物种的分布范围会发生变化[29],但1981—1994年14a的气候变异还不足以解释物种组成的变化,因而可能使得羊草群落多样性的年际变异较群落生物量的变异低。羊草群落的多样性指数、优势度指数、丰富度指数、群落高度、群落生物量在5月的年际间变异性整体高于其他月份,且8、9月份的变异性相对较低,说明气候、种间竞争等干扰因子最易影响植物的萌发及返青,而当植物进入成熟期时,羊草群落已经适应了各种干扰,从而使得群落相对稳定[18]。
不同生活型中,且多年生草本相对较为稳定,一二年生草本,灌木的变异系数相对很高,因为一二年生植物通常在遇到适宜条件时能凭借高的结种量迅速占领生境并繁衍后代[30]。羊草草原已经处于高多样性、相对稳定、以多年生草本为主的群落状态,对干扰有较高的抵抗力。不同生态型中,中生植物变异系数相对很高,旱生植物相对稳定,这是由于内蒙古典型草原属于干旱半干旱地区,多数旱生物种已经适应该地区的气候条件,且多年生草本和旱生植物中存在的补偿效应,增强了群落的稳定性[31]。
生物量的相关分析表明,相对生物量之和接近群落一半的优势种(羊草和大针茅),两者的生物量变化呈显著负相关,这些有补偿关系的物种对应着不同的敏感因子,且对干扰的敏感性及响应的趋势不同,部分物种生物量减少,因为种间关系的作用,其他物种个体数或生物量可能会有不同的变化,这种均衡效应使得群落变异性明显降低。
研究指出,羊草群落多样性的变化并不明显。因此,不是所有多样性的变化都能很好地代表群落的结构变化,而且这种由物种丰富度或多样性、优势度指数所衡量的方法在一定程度上忽略了物种组成等更具体更复杂的因素,可能不能全面反映群落的变化。同时,任何生态系统的组成物种,其功能特征在维持具有决定性的生态系统过程和生态系统服务功能上,至少和物种的量有同样重要的作用,特定的功能群及其成分较多样性本身更能决定生物产量。多样性本身既是生态系统中某些物种存在可能性的测度,又是生态系统中物种特征变异的度量。因此,植物多样性研究必须与生产力、物种组成、干扰、养分供应动态和气候综合考虑才能反映生态系统的结构和动态[32]。
3.2 羊草群落结构和生物量的关系
群落生物量与多样性的关系一直存在争议。本研究表明,羊草群落生物量和多样性没有明显的相关关系,而与优势种(羊草、大针茅)的生物量相关性显著,这两个优势物种在群落中长期占有充足有利的资源,维持着优势地位,且生物量的变化存在显著的负相关,因此优势种之间的补偿效应维持了群落的稳定性[33-34]。这并非表明多样性不重要,或许在更长的时间尺度上,非优势种及多样性的变化可能会对群落生物量及其变化产生影响。有研究表明,凋落物的累积也会影响草地生产力[35],而气候、土壤环境的变化都会影响凋落物作用,因此羊草群落的变化是多个因子综合作用的结果。
3.3 气候对羊草群落生物量的影响
在空间尺度上,降水被认为是影响生物量变化的主要因素[36-37]。但在时间尺度上,有关植物群落生物量的年际变化与降水的相关性研究仍存在争论[38-40],降水量变化对干旱半干旱区草原群落地上生物量的影响机制十分复杂。本研究表明,羊草群落生物量只与上年10月至当年8月的均温呈负相关。也有研究表明[12],群落生物量和上年10月至当8月的降水呈显著的二次曲线关系。这说明,羊草群落生物量的变化是由该地区不同时段水热因子的累积影响所致。回归分析还表明:不仅是生长季降水,包括非生长季在内的累积降水和均温对植物的生物量均有显著作用,对中生植物,生长季的降水较其他气候因子作用更明显。实验表明,温度升高引起的N释放的增加,虽然在短期内刺激了植物生长和植物净第一性生产力(NPP)的积累,长期来看却对NPP不利,因为它将引起低产的杂草和非本地物种的入侵和繁茂,这些物种固碳能力低,分解速率快(因组织中含N多),对土壤有机物的贡献小,导致土壤贫瘠和养分的流失,从而导致生物量降低[41]。也有研究认为,温度升高可以使草原趋于干旱,从而对其群落生物量产生负效应[42-43],当然并非所有的变暖效应都是负面的[44-45]。
对影响羊草群落功能群的气候因子分析表明,降水波动主要影响中生植物的变化,旱生植物对其抗性较高,其生物量变化并不明显。温度,尤其是上年10月至当年8月均温的波动主要影响群落及旱生植物生物量的变化。一二年生植物及灌木生物量的变异系数相对很高,但回归分析表明它们的生物量变化与气候因子无直接的相关性,且由于两者相对生物量较低,更易受到干扰,因此其变化可能是水热条件变化所引起的土壤条件、物种选择、资源竞争、种间作用及其他生境因子变化引起,这些变化将影响物种多样性和生产力[46],其机理有待进一步研究。
研究表明,群落多样性和生物量间不存在显著的相关性,因为气候对群落生物量有着显著的影响,且对多样性也有不同程度的影响。自然群落中,多样性与生物量之间的关系将会受到各种干扰(生物因素和非生物因素)的综合影响[47]。
4 结论
对1981—1994年间内蒙古羊草群落的调查数据和同期气象资料的分析表明,羊草群落的多样性指数、优势度指数、丰富度指数、群落高度、群落生物量在5月份的年际间变异明显高于其他月份,其中羊草群落生物量的变异从5月到9月逐渐降低,生物量和多样性之间没有显著的相关关系。羊草群落、功能群、物种的生物量年际变异依次增加。不同功能群中一二年生草本和中生植物的年际波动最大,物种间尤其是优势物种间的均衡效应降低了群落的变异性,使得羊草群落能更好地适应气候。
羊草群落生物量具有气候累积效应,主要受上年10月至当年8月的均温、1—8月的降水、上年10月至当年8月的累积降水影响,表明羊草群落生物量的变化由该地区不同时间段水热因子的综合作用决定。
[1] Parry M L.Climate Change 2007:Impacts,Adaptation and Vulnerability//Canziani O F,Palutikof J P,van der Linden P J,Hanson C E,eds.Contribution of Working GroupⅡto the Fourth Assessment Report of the Intergovernmental Panel on Climate Change.Cambridge:Cambridge University Press,2007:146-160.
[2] Cui X Y,Chen Z Z,Du Z C.Study on light and water use characteristics of main plants in semiarid Steppe.Acta Pratacultural Science,2001,10(2):14-21.
[3] Li J H,Li Z Q,Ren J Z.The effects of grazing on grassland plants.Acta Prataculturae Sinica,2002,11(1):4-11.
[4] Yu H Y,Liu S H,Zhao N,Li D,Yu Y T.Characteristics of air temperature and precipitation in different regions of China from 1951 to 2009.Journal of Meteorology and Environment,2011,27(4):1-11.
[5] Peters D P C.Plant species dominance at a grassland-shrubland ecotone:an individual-based gap dynamics model of herbaceous and woody species.Ecological Modelling,2002,152(1):5-32.
[6] Li D F,Fu T,Hu Z Z,Wang C Z,Yan X B,Jiang Y B,Guo Y X.Response of relationship between biodiversity and productivity to precipitation change.Acta Agrestia Sinica,2010,18(9):651-655.
[7] Qian S,Fu Y,Pan F F.Climate change tendency and grassland vegetation response during the growth season in Three-River Source Region.Science China:Earth Sciences,2010,53(10):1506-1512.
[8] Felix S,Schmid B.Ecosystem effects of biodiversity:a classification of hypotheses and exploration of empirical results.Ecological Applications,1999,9(3):893-912.
[9] Nippert J B,Knapp A K,Briggs J M.Intra-annual rainfall variability and grassland productivity:can the past predict the future.Plant Ecology,2006,184(1):65-74.
[10] Robinson T M P,Gross K L.The impact of altered precipitation variability on annual weed species.American Journal of Botany,2010,97(10):1625-1629.
[11] Bai Y F.Influence of seasonal distribution of precipitation on primary productivity of Stipa Krylovii community.Acta Phytoecologica Sinica,1999,23(2):155-160.
[12] Wang Y H,Zhou G S.Responses of temporal dynamics of aboveground net primary productivity of Leymus chinensis community to precipitation fluctuation in Inner Mongolia.Acta Ecologica Sinica,2004,24(6):1140-1145.
[13] Chapin F S,Walker B H,Hobbs R J,Hooper D U,Lawton J H,Sala O E,Tilman D.Biotic control over the functioning of ecosystems.Science,1997,277(5235):500-504.
[14] Perner V W,Davis A J,Eggers T,Schumacher J,Bährmann R,Fabian B,Heinrich W,Köhler G,Lichter D,Marstaller R,Sander F W.Trophic levels are differentially sensitive to climate.Ecology,2003,84(9):2444-2453.
[15] Li S Y,Li X B,Liu Z L,Wang D D,Long H L,Liang C Z,Wang W.Stability and compensation of the aboveground biomass in the Leymus Chinensis and Stipa Grandis grassland of Inner Mongolia.Resources Science,2007,29(3):152-157.
[16] Ives A R,Gross K,Klug J L.Stability and variability in competitive communities.Science,1999,286(5439):542-544.
[17] Wang H,Li X B,Yu H J.Monitoring growing season of typical steppe in northern China based on NOAA/AVHRR NDVI data.Journal of Plant Ecology,2006,30(3):365-374.
[18] Zhang C Q,Yang C.Simulation of growth dynamics of four plants in the typical steppe of Inner Mongolia in growing season.Acta Ecologica Sinica,2007,27(9):3618-3629.
[19] Li B,Yong S P,Li Z H.The vegetation of the Xilin river basin and its utilization//Inner Mongolia Grassland Ecosystem Research Station,eds.Research on Grassland Ecosystem(Vol.3).Beijing:Science Press,1988:84-183.
[20] Wang Y H,Zhou G S.Response of leymus chinensis grassland vegetation in Inner Mongolia to temperature change.Acta Phytoecologica Sinica,2004,28(4):507-514.
[21] Ren T,Hao D Y,Shi X,Liu Z L,Wang W,Liang C Z.The analysis of regression on the typical steppe in Inner Mongolia.Journal of Arid Land Resources and Environment,2001,15(3):80-84.
[22] Veech J A,Summerville K S,Crist T O,Gering J C.The additive partitioning of species diversity:recent revival of an old idea.Oikos,2002,99(1):3-9.
[23] Lou J.Partitioning diversity into independent alpha and beta components.Ecology,2007,88(10):2427-2439.
[24] Mann H B.Non-parametric tests against trend.Econometrica,1945,13(3):245-259.
[25] Li C L,Li W J.Tendency analysis and spatial pattern of aboveground vegetation biomass based on NDVI in Sunite Left Banner,Xilingole.Journal of A rid Land Resources and Environment,2010,3(3):147-152.
[26] Johnson R A,Wichern D W.Applied Multivariate Statistical Analysis.London:Prentice Hall,1989,9:308-328.
[27] Li Z Q,Liu Z G,Chen Z Z,Yang Z G.The effects of climate changes on the productivity in the Inner Mongolia steppe of China.Acta Pratacultural Science,2003,12(1):4-10.
[28] Bai Y F,Han X G,Wu J G,Chen Z Z,Li L H.Ecosystem stability and compensatory effects in the Inner Mongolia grassland.Nature,2004,431(7005):181-184.
[29] Peters R L.Effects of global warming on forests.Forest Ecology and Management,1990,35(1/2):13-33.
[30] Yan Q L,Liu Z M,Li R P,Luo Y M,Wang H M.Relationship of seed production,seed morphology and life form of plant species.Acta Pratacultural Science,2005,14(4):21-28.
[31] Wu J G,Bai Y F,Han X G,Li L H,Chen Z Z.Ecosystem stability in Inner Mongolia(reply).Nature,2005,435(7045):E6-E7.
[32] Lauenroth W.Grassland primary production:North American grasslands in perspective//French N.Perspectives in Grassland Ecology.New York:Springer-Verlag,1979,22(11):3-24.
[33] Polley H W,Wilsey B J,Derner J D.Dominant species constrain effects of species diversity on temporal variability in biomass production of tallgrass prairie.Oikos,2007,116(12):2044-2052.
[34] Lepš J.Variability in population and community biomass in a grassland community affected by environmental productivity and diversity.Oikos,2004,107(1):64-71.
[35] Wang J,Zhao M L,Walter W,Wang Z W,Han G D.Productivity responses of different functional groups to litter addition in typical grassland of Inner Mongolia.Chinese Journal of Plant Ecology,2010,34(8):907-914.
[36] McNaughton S J.Ecology of a grazing ecosystem:the Serengeti.Ecological Monographs,1985,55(3):259-294.
[37] Lauenroth W K,Sala O E.Long-term forage production of North American shortgrass Steppe.Ecological Applications,1992,2(4):397-403.
[38] Sims P L,Singh J S.The structure and function of ten Western North American grasslands.III.Net primary production,turnover and efficiencies of energy capture and water use.Journal of Ecology,1976,66(2):573-597.
[39] Abrams M D,Knapp A K,Hulbert L C.A ten year record of aboveground biomass in a Kansas tallgrass prairie:effects of fire and topographic position.American Journal Botany,1986,73(10):1509-1515.
[40] Esteban H M,Moore D I,Collins S L,Wetherill K R,Lightfoot D C.Aboveground net primary production dynamics in a northern Chihuahuan Desert ecosystem.Oecologia,2008,155(1):123-132.
[41] Kaiser J.Environent:green grass cool climate.Science,1996,274(5293):1610-1611.
[42] Xiao X M,Wang Y F,Chen Z Z.Dynamics of primary productivity and soil organic matter of typical steppe in the Xilin river basion of Inner Monglia and their response to climate change.Acta Botanica Sinica,1996,38(1):45-52.
[43] Eagles C F.The effect of temperature on vegetative growth in climatic races of Dactylis glomerata in controlled environments.Annals of Botany,1967,31(1):31-39.
[44] Hollister R D,Flaherty K J.Above-and below-ground plant biomass response to experimental warming in northern Alaska.Applied Vegetation Science,2010,13(3):378-387.
[45] Kudo G,Suzuki S.Warming effects on growth,production,and vegetation structure of alpine shrubs:a five-year experiment in northern Japan.Ecosystem Ecology,2003,135(2):280-287.
[46] Song L L,Fan J W,Zhong H P,Wang N.Changes of biomass and species richness of grassland community along an altitude gradient in Hongchiba,Chongqing.Acta Agrestia Sinica,2010,18(2):160-166.
[47] Tilman D,Reich P B,Knops J M H.Biodiversity and ecosystem stability in a decade-long grassland experiment.Nature,2006,441(1):629-632.
参考文献:
[2] 崔骁勇,陈佐忠,杜占池.半干旱草原主要植物光能和水分利用特征的研究.草业学报,2001,10(2):14-21.
[3] 李金花,李镇清,任继周.放牧对草原植物的影响.草业学报,2002,11(1):4-11.
[4] 虞海燕,刘树华,赵娜,李栋,于永涛.1951—2009年中国不同区域气温和降水量变化特征.气象与环境学报,2011,27(4):1-11.
[6] 李德峰,傅彤,胡自治,王成章,严学冰,姜义宝,郭玉霞.生物多样性与生产力间的关系及对降水变化的响应.草地学报,2010,18(5):651-655.
[11] 白永飞.降水量季节分配对克氏针茅草原群落初级生产力的影响.植物生态学报,1999,23(2):155-160.
[12] 王玉辉,周广胜.内蒙古羊草草原植物群落地上初级生产力时间动态对降水变化的响应.生态学报,2004,24(6):1140-1145.
[15] 李素英,李晓兵,刘忠龄,王丹丹,龙惠灵,梁存柱,王炜.内蒙古羊草大针茅草原的地上生物量稳定性与补偿性研究.资源科学,2007,29(3):152-157.
[17] 王宏,李晓兵,余弘婧.基于NOAA/AVHRR NDVI监测中国北方典型草原的生长季及变化.植物生态学报,2006,30(3):365-374.
[18] 张彩琴,杨持.内蒙古典型草原生长季内不同植物生长动态的模拟.生态学报,2007,27(9):3618-3629.
[19] 李博,雍世鹏.锡林河流域植被及其利用 //草原生态系统研究(第三集).北京:科学出版社,1988:84-183.
[20] 王玉辉,周广胜.内蒙古地区羊草草原植被对温度变化的动态响应.植物生态学报,2004,28(4):507-514.
[21] 任涛,郝敦元,石霞,刘钟龄,王炜,梁存柱.内蒙古典型草原原生群落植物种群动态趋势分析.干旱区资源与环境,2001,15(3):80-84.
[25] 李昌凌,李文军.基于NDVI的锡盟苏尼特左旗地表植被生物量的趋势分析和空间格局.干旱区资源与环境,2010,24(3):147-152.
[27] 李振清,刘振国,陈佐忠,杨宗贵.中国典型草原区气候变化及其对生产力的影响.草业学报,2003,12(1):4-10.
[30] 闫巧玲,刘志民,李荣平,骆永明,王红梅.科尔沁沙地75种植物结种量种子形态和植物生活型关系研究.草业学报,2005,14(4):21-28.
[35] 王静,赵萌莉,Walter W王忠武,韩国栋.内蒙古典型草原不同功能群生产力对凋落物添加的响应.植物生态学报,2010,34(8):907-914.
[42] 肖向明,王义凤,陈佐忠.内蒙古锡林河流域典型草原初级生产力和土壤有机质的动态及其对气候变化的反应.植物学报,1996,38(1):45-52.
[46] 宋璐璐,樊江文,钟华平,王宁.重庆红池坝地区草地群落生物量及物种丰富度沿海拔梯度格局变化.草地学报,2010,18(2):160-166.
