葡萄溃疡病菌转化子致病力快速评价体系的建立
2013-09-11刘爱芬燕继晔刘瑞琪庞倩茹李兴红黄金光
刘爱芬, 张 鑫, 张 玮, 燕继晔,刘瑞琪, 庞倩茹, 李兴红, 黄金光*
(1.青岛农业大学农学与植物保护学院,青岛 266109;2.山东省植物病虫害综合防控重点实验室,青岛 266109;3.北京市农林科学院植物保护环境保护研究所,北京 100097;4.河北农业大学生命科学学院,保定 071001)
葡萄是世界上重要的经济果树之一,据国家葡萄产业体系统计,随着葡萄所创造的经济效益的增加,截至2010年底我国葡萄种植面积已增至53万hm2。长期以来,葡萄病害是影响葡萄产业发展的重要因素,近几年研究发现葡萄溃疡病在我国葡萄主产区造成了严重危害,且有加重趋势。葡萄溃疡病主要由葡萄座腔菌科(Botryosphaeriaceae)真菌引起,病原菌侵染后可导致维管束变褐,进而使枝干或主干坏死,严重时会造成整株死亡[1]。目前,葡萄溃疡病已在美国、法国、西班牙等17个国家发现[2],在不同国家均造成了一定程度的危害。据统计,由葡萄溃疡病在美国加利福尼亚州造成的危害仅2001年就损失2.6亿美元,约占加州地区葡萄总产值的10%[3]。在我国,李兴红研究组2009年首次报道了Botryosphaeria dothidea[4]和B.rhodina[1]引起的葡萄溃疡病。目前,该病害在山东、广西、辽宁、湖北、湖南、四川、甘肃、河南等葡萄主产区均已有发现。
目前,关于葡萄溃疡病的报道集中于病原菌的鉴定和种群结构方面,对病原菌致病机理的研究尚未见报道[2]。随着分子生物学和分子遗传学技术的发展,人们开始用分子生物学的方法去获得插入转化子,为挖掘病菌的致病基因进而明确其致病机理做准备。然而对于一个庞大的转化子库,能否快速、准确地筛选出致病力突变体尤为重要。有关葡萄座腔菌科真菌致病力的室内评价方法,通常是通过接种离体的绿枝条,Brown-Rytlewski和 McManus已成功运用绿枝条接种方法比较了B.dothidea和B.obtusa的致病力强弱[5],Urbez-Torres 通过接种葡萄绿枝条将葡萄座腔菌科9个种的致病力分为高、中、低3个等级[6]。但是利用绿枝条对转化子库进行筛选会遇到很多限制因素。如:葡萄绿枝条生长受季节的影响,并且同时采集大量同一品种的粗细、木质化程度等较为一致的葡萄绿枝条非常困难,致使转化子库的评价工作会受到季节的影响。为了建立一个快速高效的葡萄溃疡病菌转化子致病力评价体系,本研究从REMI转化子库中随机挑选50株转化子,探讨了以‘富士’苹果果实为接种替代物,3种接种方法、4个不同菌龄对葡萄溃疡病菌致病力的影响,并与野生型菌株CSS-01s的致病力进行比较,同时采用了接种葡萄绿枝条的方法[7]对评价结果进行验证。
1 材料与方法
1.1 材料
供试菌株:野生型葡萄溃疡病菌(B.rhodina)菌株CSS-01s,由本实验室保存;50株葡萄溃疡病菌REMI转化子,由本实验室转化子库中随机选取。
供试接种材料:直径为(20±5)cm的烟台‘富士’苹果果实(一级果),购于超市发超市,一年生‘夏黑’葡萄绿枝条由北京市农林科学院林业果树研究所提供。
1.2 在苹果果实上的接种
1.2.1 不同接种方法对致病力测定的影响
分别用无伤、昆虫针针刺(1、4、8针)、5mm 的打孔器去掉果皮5种方法处理同一苹果。将直径为4mm的CSS-01s菌饼接种到相应的处理处,重复6次。将接种后的苹果放入保鲜盒中28℃保湿培养,每12h观察并测量病斑大小。
1.2.2 不同菌龄对致病力测定的影响
将菌龄为1、2、3、4d的野生型菌株和随机挑选的5个转化子采用打孔器法接种苹果,每个果实7处伤口,分别将直径为4mm的菌饼和无菌培养基接种到同一果实上,每个菌龄重复5次,并将其放于28℃培养箱中保湿培养,每12h记录其发病状况。
1.2.3 不同转化子的致病力测定
分别将随机挑选的50株转化子、野生型菌株CSS-01s的4mm菌饼接种到用打孔器处理过的伤口上,在同一个苹果最大纬线上均匀分布4处伤口,分别将2个转化子及野生型菌饼和无菌培养基接种到相应的伤口上,每个处理3次重复,将其放于28℃培养箱中保湿培养,3d后测量并记录病斑大小。
1.3 在葡萄绿枝条上不同转化子的致病力测定
参考 Urbez-Torres[6]等和燕继晔[7]等测定葡萄溃疡病菌(B.dothidea)致病力的方法,将在苹果上做过筛选的50株转化子和野生型CSS-01s接种一年生葡萄绿枝条(‘夏黑’),每个处理重复5次,5d后观察并测量接种结果。
1.4 数据分析
所得数据均采用SAS 9.1数据分析软件处理。不同菌龄处理间致病力的显著性差异采用LSD法,转化子在两种不同寄主植物上致病力的显著性差异采用Duncan氏新复极差法。检验水平均为P≤0.05。
2 结果与分析
2.1 在苹果果实上不同接种方法对菌株发病速度的影响
不同接种方法对野生型菌株CSS-01s在苹果果实上是否发病及发病快慢均有影响。无伤接种不能引起‘富士’苹果果实发病,针刺和打孔法均能使苹果果实在接种点处出现水渍状棕黄色病斑。在接种12h后,用打孔器打孔接种的6个苹果果实均出现症状,但用针刺接种的6个苹果果实在接种29h后才表现症状。在接种36h后,用打孔器打孔法接种的苹果果实病斑直径为4.33cm,而用针刺1针接种的苹果果实病斑直径为2.74cm,针刺4针和8针的苹果果实病斑直径分别为2.92cm和3.04cm。在接种46h后,苹果果实上不同处理间的病斑大小已无法区分。用打孔器打孔的方法比用针刺接种发病快,且一致性高,因此用打孔器的方法便可缩短测量时间,加快筛选进度(图1)。

图1 不同接种方法接种野生型菌株CSS-01s 2d后在苹果果实上的病斑直径Fig.1 Lesion diameter on apple fruit at 2days after inoculation with wild type strain CSS-01sby different methods
2.2 在苹果果实上不同菌龄对转化子致病力的影响
在苹果果实上,接种16h后,菌龄为1、2、3d的野生型菌株和转化子均出现棕黄色病斑,菌龄为4d的CSS-01s也出现棕黄色病斑,然而5个转化子均未出现棕黄色病斑,接种64h后,5个不同菌龄的转化子和CSS-01s均使苹果果实发病。其中菌龄为1、2、3、4d的CSS-01s在苹果果实上的病斑直径分别为1.60、0.87、1.0cm 和0.56cm,所以随着菌龄的增加,病原菌侵染苹果果实并使其产生病斑的时间相应延长。但是通过比较菌龄为1d的5个转化子和CSS-01s的病斑直径可以看出和CSS-01s具有极显著差异的转化子为k31、k32、k34、k37,而通过比较菌龄为2、3、4d的评价结果可以发现,和CSS-01s具有极显著差异的转化子均为k32、k37。所以菌龄会影响菌株的发病快慢,原因可能是菌龄与菌丝的活力等有关。如图2,菌龄为1d的CSS-01s菌落颜色为白色,气生菌丝稀疏,刚长满培养皿。而菌龄为2、3、4d的CSS-01s菌落颜色逐渐由白色变为褐色,气生菌丝越来越致密。通过比较4个菌龄的筛选结果,发现菌龄为2d的接种结果已经稳定(图3)。

图2 不同菌龄的CSS-01s在PDA上菌落形态Fig.2 Colony morphology of CSS-01swith different cell age on PDA plate

图3 不同菌龄的转化子与野生型接种2d后在苹果果实上的病斑直径Fig.3 Lesion diameter on apple fruit at 2days after inoculation with wild type strain CSS-01s and the transformants with different cell ages
2.3 在苹果果实和葡萄绿枝条上致病力结果的比较
通过比较50株REMI转化子与野生型菌株CSS-01s在苹果果实上的病斑大小发现,CSS-01s在‘富士’苹果果实上的病斑直径为5.58cm,有13株转化子与CSS-01s致病力没有显著差异,而有37株比CSS-01s的致病力减弱。参照燕继晔[7]等绿枝条接种的方法将50株转化子和CSS-01s接种葡萄绿枝条,通过比较转化子和CSS-01s在葡萄绿枝条上的病斑长度,发现CSS-01s接种的病斑长度为7.0cm,比其致病力增强的转化子有21株,致病力减弱的有26株,没有显著差异有3株。通过比较50株转化子在这两种寄主上的筛选结果发现,用葡萄绿枝条筛选的致病力减弱的转化子有25株与在苹果上的筛选结果一致。由此可见,在葡萄绿枝条上致病力减弱的转化子有96.15%在‘富士’苹果果实上致病力也是减弱的,但是致病力增强的转化子在苹果果实上却没有筛选到。
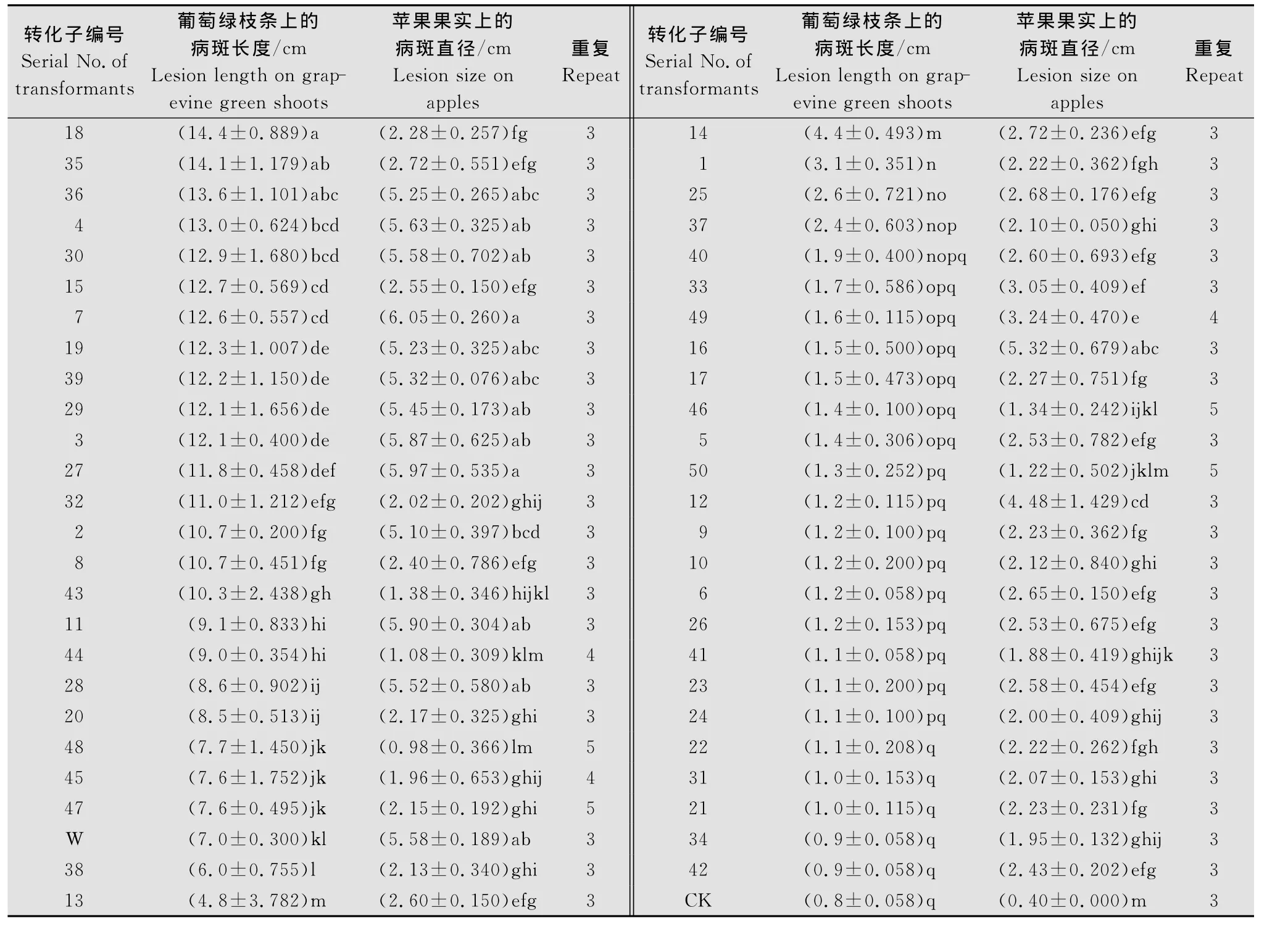
表1 随机选取的50株转化子和野生型菌株在葡萄绿枝条和苹果果实上的病斑大小Table 1 Lesion size on grapevine green shoots and apple fruit after inoculation with wild type strain and the 50transformants selected randomly
3 结论与讨论
对枝干病害病原菌致病力的评价,通常采用接种绿枝条的方法。Chang等[9]、Rozsnyay和 Apostol[10]分别用桃和樱桃绿枝条作为寄主对病原菌的致病性进行评价,发现评价结果与田间活体接种结果一致。林月莉等将B.dothidea分别接种苹果绿枝条、苹果果实及苹果叶片,3种材料的接种结果与田间接种苹果枝条的试验结果一致[11]。葡萄溃疡病菌转化子致病力的评价也可应用一年生葡萄枝条,Urbez-Torres[12]在2008年用一年生葡萄绿枝条比较 Lasiodiplodia theobromae (B.rhodina)和Diplodia seriata (B.obtusa)的致病力强弱;本研究室燕继晔[7]等在2012年利用同样的方法评价了分离自4个不同地方的B.dothidea单孢菌株在25个不同葡萄品种上的致病力。但在进行大规模转化子致病力研究时,由于葡萄绿枝条取材受季节的限制,且需求量大,使得研究工作不能顺利进行。因此能够找到一种可以不受季节限制,并且易获得的接种材料替代葡萄绿枝条便会加快对转化子库致病力突变体的筛选。本研究室已利用此方法完成了25个葡萄品种对B.rhodina的抗性评价,发现‘夏黑’为感病品种,且预试验发现B.rhodina也可以侵染‘富士’苹果果实。所以本研究试图将苹果果实作为葡萄绿枝条的替代物,结果发现在葡萄绿枝条上致病力减弱的转化子有96.15%在‘富士’苹果果实上的致病力也是减弱的。所以‘富士’苹果果实可以用来对转化子库进行初筛获得致病力减弱的突变体,为进一步研究致病基因提供可能。用苹果果实代替绿枝条接种,可缩短致病力突变体的筛选周期。
在苹果果实上没有筛选到致病力增强的转化子,推测原因是由于B.rhodina在两种寄主上的适应力、侵染力不同,较葡萄枝条而言,野生型CSS-01s在‘富士’苹果果实上的致病力强且发病迅速、无法区别致病力增强的转化子。
本研究初步建立了利用‘富士’苹果果实作为寄主替代物筛选致病力减弱转化子的筛选体系,但该体系尚不完善还需进一步优化,如:苹果果实品种、品质等等都需进一步优化。若能优化筛选体系,便为后期进一步研究致病基因提供支持,进而为解析葡萄溃疡病菌致病机理奠定基础,从而控制该病害在田间的发生,保障葡萄产业的顺利发展。
[1] Yan J Y,Li X H,Kong F F,et al.Occurrence of grapevine trunk disease caused by Botryosphaeria rhodinain China[J].Plant Disease,2011,95(2):219.
[2] Urbez-Torres J R.The status of Botryosphaeriaceae species infecting grapevines[J].Phytopathologia Mediterranea,2011,50(4):5-45.
[3] Siebert J B.Eutypa:The economic toll on vineyards[J].Wines & Vines,2001(4):50-56.
[4] Li Xinghong,Yan Jiye,Kong Fanfang,et al.Botryosphaeria dothidea causing canker of grapevine newly reported in China[J].New Disease Reports,2010,21:25.
[5] Brown-Rytlewski D E,Mcmanus P S.Virulence of Botryosphaeria dothidea and Botryosphaeria obtusa on apple and management of stem cankers with fungicides[J].Plant Disease,2000,84(9):1031-1037.
[6] Urbez-Torres J R,Gubler W D.Pathogenicity of Botryosphaeriaceae species isolated from grapevine cankers in California[J].Plant Disease,2009,93(6):584-592.
[7] Yan Jiye,Xie Yue,Yao Shengwei,et al.Characterization of Botryosphaeria dothidea,the causal agent of grapevine canker in China[J].Australasian Plant Pathology,2012,41(4):351-357.
[8] Pavlic D.Botryosphaeria species on native South African Syzygium cordatumand their potential threat to Eucalyptus[D].University of Pretoria,2004.
[9] Chang L S,Iezzoni A F,Adams G.Excised-shoot assay for tolerance of peach to Leucostoma persoonii[J].HortScience,1989,24(6):1011-1012.
[10]Rozsnyay Z S,Apostol J.Breeding for sweet and sour cherry disease resistance in Hungary[C].ISHS Acta Horticulturae 667:ⅣInternational Cherry Symposium.2001.
[11]林月莉,黄丽丽,高小宁,等.苹果轮纹病室内快速评价体系的建立[J].植物保护学报,2011,38(1):37-41.
[12]Urbez-Torres J R,Leavitt G M,Guerrero J C,et al.Identification and pathogenicity of Lasiodiplodia theobromae and Diplodia seriata,the causal agents of bot canker disease of grapevines in Mexico[J].Plant Disease,2008,92(4):519-529.
