太湖湿地昆虫群落结构及多样性
2013-09-07韩争伟曹传旺王步勇
韩争伟,马 玲,曹传旺,张 静,王步勇
(东北林业大学林学院,哈尔滨 150040)
湿地环境中生活着种类众多、数量巨大的昆虫,它们是湿地生态系统中物种多样性的重要组成部分,在湿地环境的物质和能量流动与转化中起着不可忽视的作用[1]。近些年来,关于湿地昆虫的研究已取得很大进展[2-7]。彭筱葳[8]等对九段沙湿地自然保护区不同生境中的昆虫群落进行了研究,表明昆虫群落组成和结构与所处生境中的植被密切相关。尤平[1]等利用蛾类昆虫群落的特征指数对天津湿地的环境进行了评价。顾伟[9]等研究了扎龙湿地不同生境对昆虫群落多样性的影响。Alan P.Bedford[10]等研究了芦苇湿地水位的年际变化对昆虫群落结构的影响。Vassiliki Kati[11]等研究了湿地开发利用对湿地特有昆虫种类的影响。因此,昆虫群落的结构和动态变化应该可以作为湿地环境变化的参考。
太湖湿地是我国东部近海地区最大的湖泊,也是我国著名的风景名胜区,对该地区昆虫群落的研究尚未见有报道。太湖周边地区经济迅速发展的同时,太湖湿地环境却受到了严重的破坏。随着近年来人们对太湖湿地的关注,湿地生境的恢复工作一直在进行中,但湿地原有植被状况的改变和湿地旅游设施的修建也成为现在太湖湿地人为干扰的主导因子。本文选取人工干扰下太湖湿地不同生境的样点,调查和分析昆虫群落的多样性、结构以及稳定性,探讨了昆虫群落在不同环境的生态反应,旨在为太湖湿地昆虫生态学的研究提供基础资料,也为太湖湿地生物多样性的保护和湿地的可持续利用提供理论参考。
图1 样点分布图Fig.1 The sample locations
1 研究地区和研究方法
1.1 研究区域概况
太湖湿地(北纬 30°56'—31°34',东经 119°54'—120°36')位于长江三角洲南缘,是目前我国第二大淡水湖,全水域面积为2428 km2,平均蓄水量为44亿m3,属于北亚热带南部向北亚热带北部过渡的东西季风气候区,四季分明,无霜期长,热量充裕,降水丰沛。
本研究根据太湖湿地的植被类型和人为干扰状况选择了5个不同生境类型的样点,分别是:苏州太湖的灵湖村样点、三山岛样点、三洋村样点、渔阳山样点和无锡太湖的示范区样点;灵湖村样点设置在湖边湿地公园,三山岛样点设置在湖心三山岛,渔阳山样点设置在山区水源保护区,三洋村样点设置在非山区水源保护区,示范区样点设置在国家林业局湿地恢复示范区。各样点分布如图1所示。
各样点植被群落主要有芦苇群落;芦苇、香蒲群落;芦苇、茭草、荻草群落;酢浆草、野豌豆、白三叶等景观草本群落;狗尾草、葎草、空心莲子草等杂草群落;樟树、水杉、垂柳等组成的乔木群落;以及湖岸边樟、柳、水杉等乔木的零星分布。各个样点的植被分布见表1。

表1 各样点植被分布Table1 The vegetation of the samples
1.2 研究方法
1.2.1 调查方法
调查采样时间为2010年的5月至9月和2011年的5月至9月,每月在各样点采样2次,基本为上半月1次下半月1次,每次采样时间为昆虫活动频繁的10:00—15:00。每样点设置3块样地(20 m×20 m),样地排列按照实际地形而定。采样方法以扫网法为主,每样地扫100次,扫网路径为“Z”字形来回扫取,扫网时一个往返为1次,每次往返180°;将采集的所有标本投入到毒瓶中杀死,带回实验室挑拣所有昆虫标本,进行计数和种类鉴定。标本鉴定主要采取形态分类法,所有标本至少鉴定到科,部分疑难标本请教相关专家鉴定,未能鉴定到种的昆虫标本由于数量极少所以将其编号作为单独的种进行处理。蜘蛛类与昆虫群落稳定性的关系极为密切,所以在进行群落稳定性分析时将蜘蛛类群包括在内。
按照昆虫的食性,将昆虫群落大致分为以下4个类群[12]:即捕食性类群、植食性类群、寄生性类群和中性类群(包括腐生性的类群和对植物没有危害也不以其它昆虫为食的类群)。
1.3 数据处理
文中所有数据均使用Excel和SPSS18.0统计分析软件处理(各指标均以2a的综合数据计算所得)。
物种丰富度(S)和个体总数(N),在多样性分析中分别采用其自然对数(ln S和ln N)。多样性应用Shannon-Wiener多样性指数H=- ln(Pi);Simpson优势集中性指数C=i)2,式中Pi为第i种个体占个体总数的比率。
Berger-Parker优势度指数:I=Nmax/N,式中Nmax为优势类群的数量,N为全部的种群数量。
均匀度(Evenness,J)采用Pielou公式:J=H'/ln S;式中H'为Shannon-Wiener多样性指数,S为群落中物种数[1,13]。
群落相似性系数:D=2W/(a+b),式中D为群落系数;W为样地A和B共有种的种数;a为样地A的全部种数;b 为样地 B 的全部种数[1,14]。
以2a来调查数据为样本,以昆虫(按照食性)物种数和个体数为变量,进行主分量分析,并且计算各因子的特征向量和负荷量值[15-17]。通过排序将各样点昆虫群落作为点,并且用各群落属性作为坐标轴,在二维空间中按照各昆虫群落的相似关系将其排列[14]。
群落相对稳定性的分析,采用群落物种数和个体数之比(St/Si)和天敌类群种数和植食性类群种数之比(Sn/Sp)表示;St/Si反映种间数量上的制约作用,Sn/Sp反映食物网关系的复杂程度和相互制约的程度[18]。
2 结果与分析
2.1 太湖湿地昆虫群落组成结构
实验调查中共获得昆虫标本24968号(另有蜘蛛类标本385号),隶属于13目,88科,301属,318种(表2);其中半翅目 (Hemiptera)昆虫60种,占总种数的18.86%;鳞翅目(Lepidoptera)昆虫58种,占总种数的18.24%;鞘翅目(Coleoptera)昆虫60种,占总种数的18.86%;膜翅目(Hymenoptera)昆虫56种,占总种数的17.61%;直翅目(Orthoptera)25种,占总种数的7.86%;5个目的昆虫种数占总种数的81.44%。各昆虫类群数量上,蜻蜓目(Odonata)占总数量的23.09%;半翅目占总数量的19.23%;直翅目占总数量的18.12%;鞘翅目占总数量的12.75%;膜翅目占总数量的8.33%;鳞翅目占总数量的6.69%,这6个目昆虫数量占昆虫总数量的 88.21%。
太湖湿地中的优势类群包括:半翅目的蝽科Pentatomidae、缘蝽科Coreidae以及叶蝉科Cicadellidae;鳞翅目的粉蝶科 Pieridae、蛱蝶科 Nymphalidae和螟蛾科 Pyralidae;鞘翅目的瓢虫科 Coccinellidae和叶甲科Chrysomelidae;直翅目的剑角蝗科Acrididae、锥头蝗科Pyrgomorphidae和蟋蟀科 Gryllidae;膜翅目的蜜蜂科Apidae以及蜻蜓目的蜻科Libellulidae和蟌科Coenagrionidae等。
常见的昆虫种类为:麻皮蝽(Erthesina fulllo(Thunberg))、华麦蝽(Aelia nasuta Wagner)、菜蝽(Eurydema dominulus(Scopoli))、稻棘缘蝽(Cletus punctiger Dallas)、菜粉蝶(Pieris rapae(Linnaeus))、斑缘豆粉蝶(Colias erate(Esper))、樟青凤蝶(Graphium sarpedon Linnaeus)、黄钩蛱蝶(Polygonia c-aureum(Linnaeus))、七星瓢虫(Coccinella sptempunctata Linnaeus)、龟纹瓢虫(Propylea japonica(Thunberg))、大黑鳃金龟(Holotrichia oblita(Faldermann))、中华剑角蝗(Acrida cinerea(Thunberg))、短额负蝗(Atractomorpha sinensis Bolivarr)等。
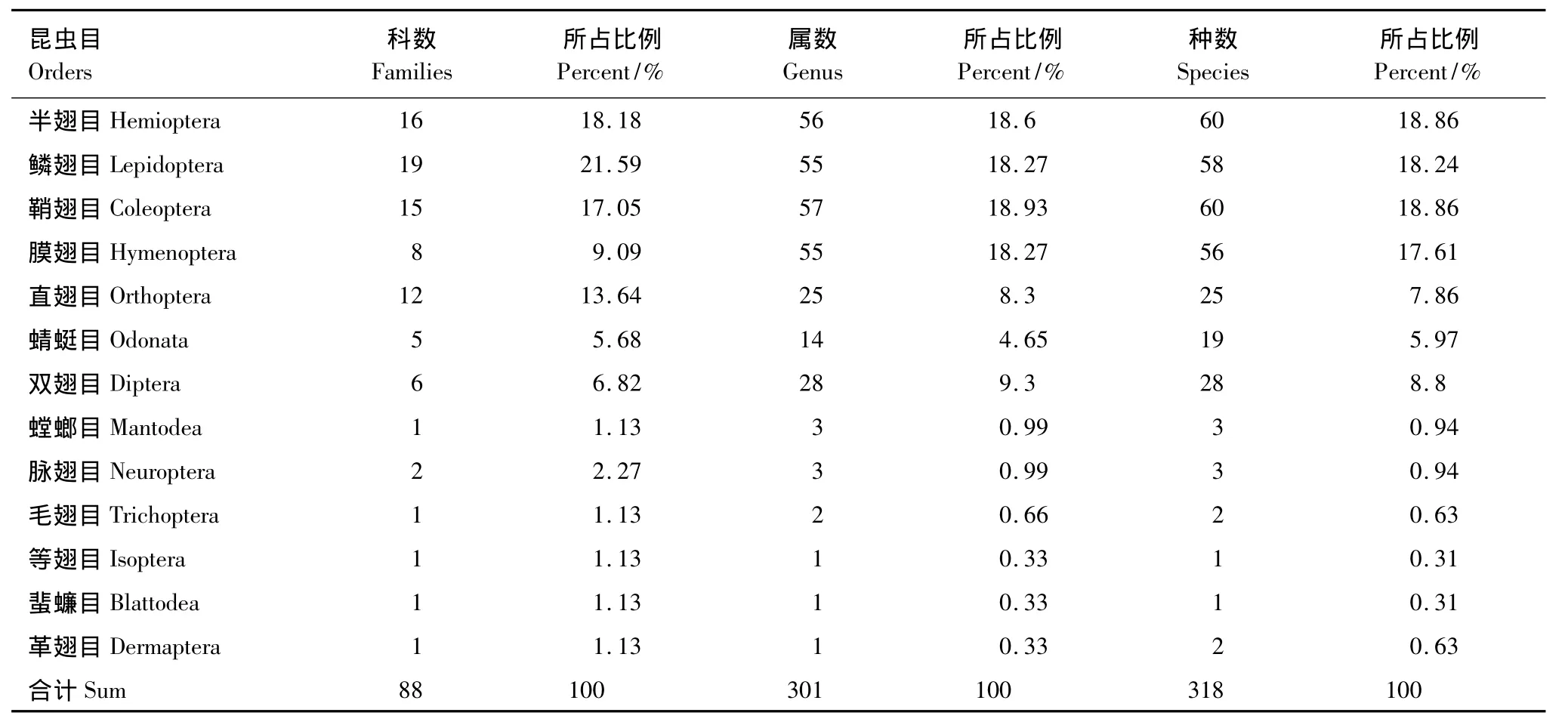
表2 太湖湿地昆虫群落结构Table2 The structure of insect communities from Taihu Wetland
2.2 昆虫群落多样性特征指数
从表3可以看出,太湖湿地各样点昆虫群落的多样性指数均比较高。示范区和渔阳山样点昆虫群落多样性明显高于其它样点,灵湖村昆虫群落多样性指数最低,但其物种丰富度指数却最高;同时,灵湖村昆虫群落中优势类群的数量多,优势度指数和优势集中性指数高。各样点昆虫群落的多样性指数和均匀度指数特征值的比较为:示范区>渔阳山>三山岛>三洋村>灵湖村;优势集中性指数的特征值比较为:示范区<渔阳山<三洋村<三山岛<灵湖村,说明示范区和渔阳山的昆虫群落物种丰富,内部结构比较复杂,相对其它样点来说更加稳定。
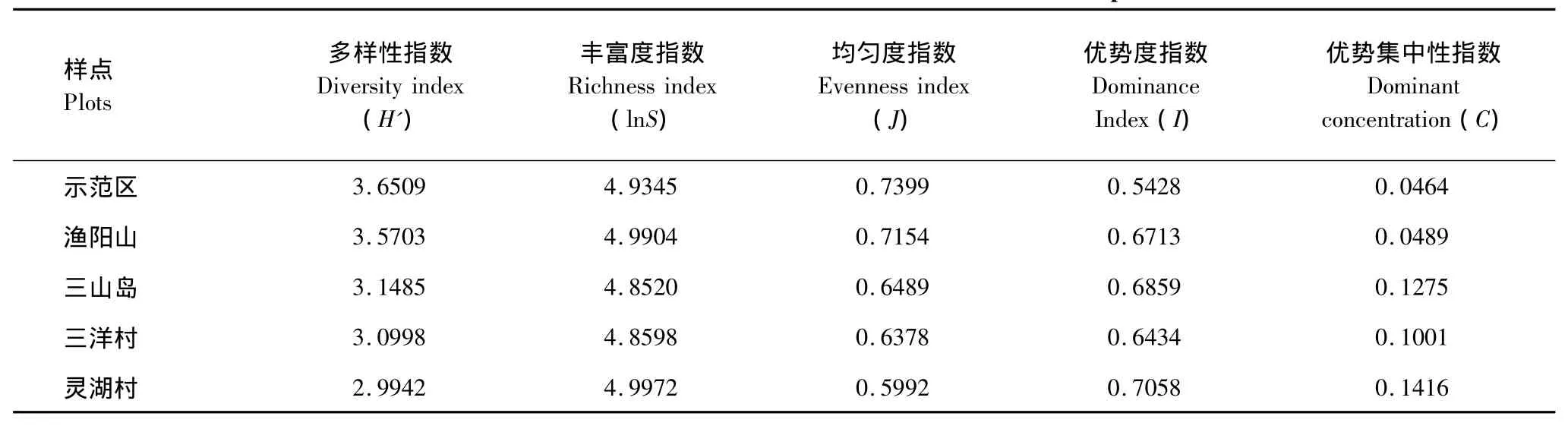
表3 各样点昆虫群落特征值Table3 Characteristic values of insect communities in different plots
2.3 昆虫群落多样性特征指数时间动态
从图2可以看出各样点昆虫群落多样性指数在6月份之前的波动小且相互比较接近,6月之后波动变大;灵湖村的多样性指数最高(2.6918)出现在6月下半月;三山岛多样性指数在7月上半月达到最高(3.5357),之后曲折下降;三洋村多样性最高(3.0235)出现在6月下半月,全年多样性指数变化幅度大。从时间序列来看,灵湖村、三山岛和三洋村3样点的多样性多位于示范区和渔阳山样点多样性曲线下方,波动幅度也相对较大。渔阳山多样性在9月上半月达到最高(3.2405),而示范区的多样性最高(3.5566)出现在9月下半月,这与此时样点中的膜翅目昆虫种类增多,昆虫群落均匀度最大有很大关系,并且两样点在时间序列上的波动幅度相对较小。从整体来看渔阳山样点的多样性指数随时间变化的波动最小,说明渔阳山样点昆虫群落是比较稳定的。
灵湖村昆虫群落的均匀度指数在6月下半月达到最高(0.7767);三洋村的均匀度指数最大(0.7989)也出现在6月下半月,其它各月的变化表现不一,波动大。三山岛均匀度指数最高(0.8948)出现在7月上半月,其它各月曲折减小。示范区均匀度的最高值(0.8552)和渔阳山均匀度的最高值(0.8162)分别出现在9月的下半月和上半月,并且在时间序列上波动小,表现平稳。总之,昆虫群落的均匀度指数变化趋势和多样性指数保持一致,多样性指数高或低时,均匀度指数也相应的表现为高或者是低。
灵湖村昆虫个体数量最高出现在9月上半月,最低出现在6月下半月;三山岛昆虫个体数量最高出现在9月下半月,最低则出现在7月上半月。虽然灵湖村和三山岛的个体数量最高都出现在9月,但是前者在8月下半月和9月下半月之间是呈倒“V”型,波动大,而后者在此期间呈逐渐上升趋势。从时间序列来看,三洋村、示范区和渔阳山样点的昆虫个体数量呈曲折前进的趋势,而且波动的幅度小。灵湖村和三山岛样点昆虫群落个体数量在9月份变化剧烈,是因为个别昆虫类群数量受小环境影响突然增多,例如:三洋村在9月份时,鞘翅沫蝉(Lepyronia coleopterata(Linnaeus))会突然大量出现,导致昆虫群落数量升高。
各样点昆虫物种数随时间变化。渔阳山的昆虫物种数最高值出现在7月下半月,在此之前物种数呈逐渐上升趋势,之后则曲折减少。示范区的物种数最高值在9月下半月出现,之前一直是曲折上升的。灵湖村、三山岛和三洋村的物种数量各月份的波动幅度各有不同。各样点昆虫物种数量的波动,是由各样点环境中的植被类型、人为干扰以及气候变化等多因素综合的结果。
昆虫群落优势集中性指数的变化趋势与多样性指数正好是相反的。从图2中可以看出,示范区和渔阳山的优势集中性指数曲线位于其它3样点的优势集中性曲线下方,并且表现平稳,而其它3样点的曲线则表现不稳定,波动幅度大。
昆虫群落的均匀度指数和丰富度指数与昆虫群落的多样性指数之间关系是密切的[19]。太湖湿地各样点昆虫群落的均匀度指数和多样性指数表现一致,呈正相关(r=0.792-0.99,P<0.01)(表4)。昆虫群落的丰富度指数与多样性指数也呈正相关,不过只有示范区和渔阳山的相关系数在0.01的水平上。
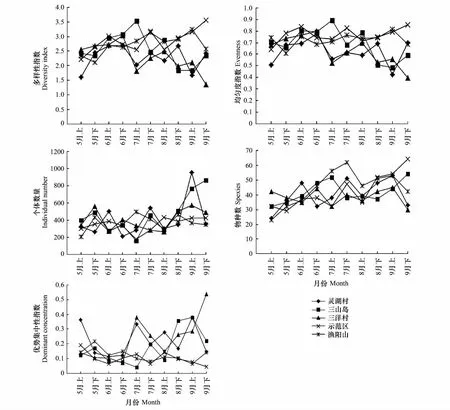
图2 各样点昆虫群落多样性指数(H')、均匀度指数(J)、个体数量(ln N)、物种丰富度指数(ln S)和优势集中性指数(C)的时间动态Fig.2 The time-pattern tendency of diversity index(H'),evenness index(J),individual number(ln N),specific richness(ln S)and dominant concentration index(C)of insect communities in different plots

表4 多样性特征指数间的相关系数Table4 The correlation coefficients(r)of the indices of diversity
2.4 昆虫群落的主分量分析
为了能够分析影响昆虫群落的内部机制和主导因素,文中以捕食性昆虫的物种数为X1、以捕食性昆虫的个体数为X2、以植食性昆虫的物种数为X3、以植食性昆虫的个体数为X4、以寄生性昆虫的物种数为X5、以寄生性昆虫的个体数为X6、以中性昆虫的物种数为X7、以中性昆虫的个体数为X8(各类群分布见表5),分别对各样点的昆虫群落进行主分量的分析,各因子特征向量值见表6。
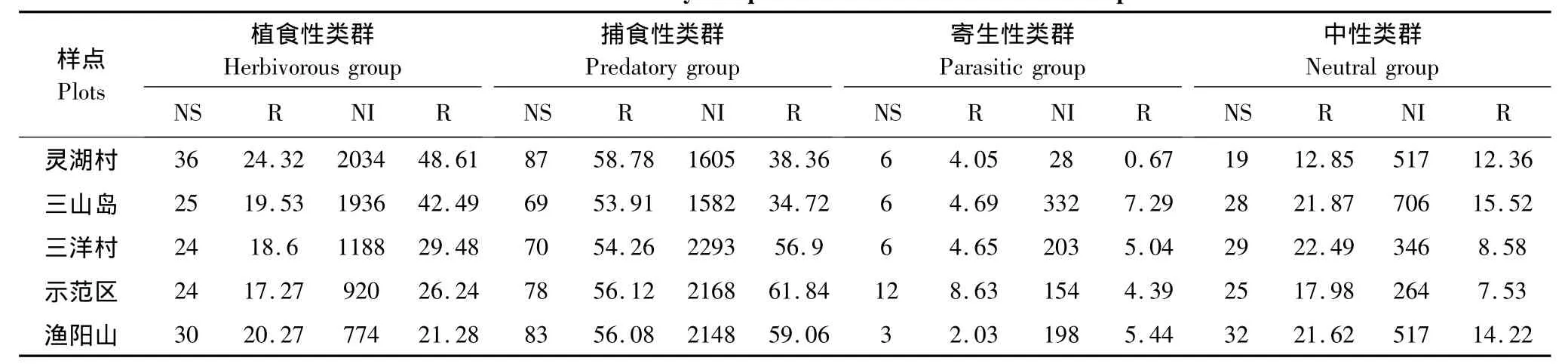
表5 各样点昆虫群落组成及结构Table5 Insect community composition and structure in different plots
从表6中可以看出:5个样点的昆虫群落中前3个主分量的累计贡献率均超过了80%,其中三山岛昆虫群落的累计贡献率高达97.4%;灵湖村昆虫群落的第一主分量代表了捕食性昆虫个体数、植食性昆虫物种数和寄生性昆虫个体数的综合因子,贡献率为42.4%,这也说明了捕食性昆虫个体数、植食性昆虫物种数以及寄生性昆虫个体数对群落影响的重要性;第二主分量主要代表了植食性昆虫个体数、寄生性昆虫物种数和中性昆虫物种数的综合因子;第三主分量代表了寄生性昆虫物种数因子。3个主分量的累计贡献率为90.2%,第三主分量的贡献率(11.1%)低于第一主分量贡献率(42.4%)和第二主分量贡献率(36.7%),结果表明灵湖村昆虫群落中捕食性昆虫个体数、植食性昆虫物种数以及寄生性昆虫个体数是影响昆虫群落的主要因子。
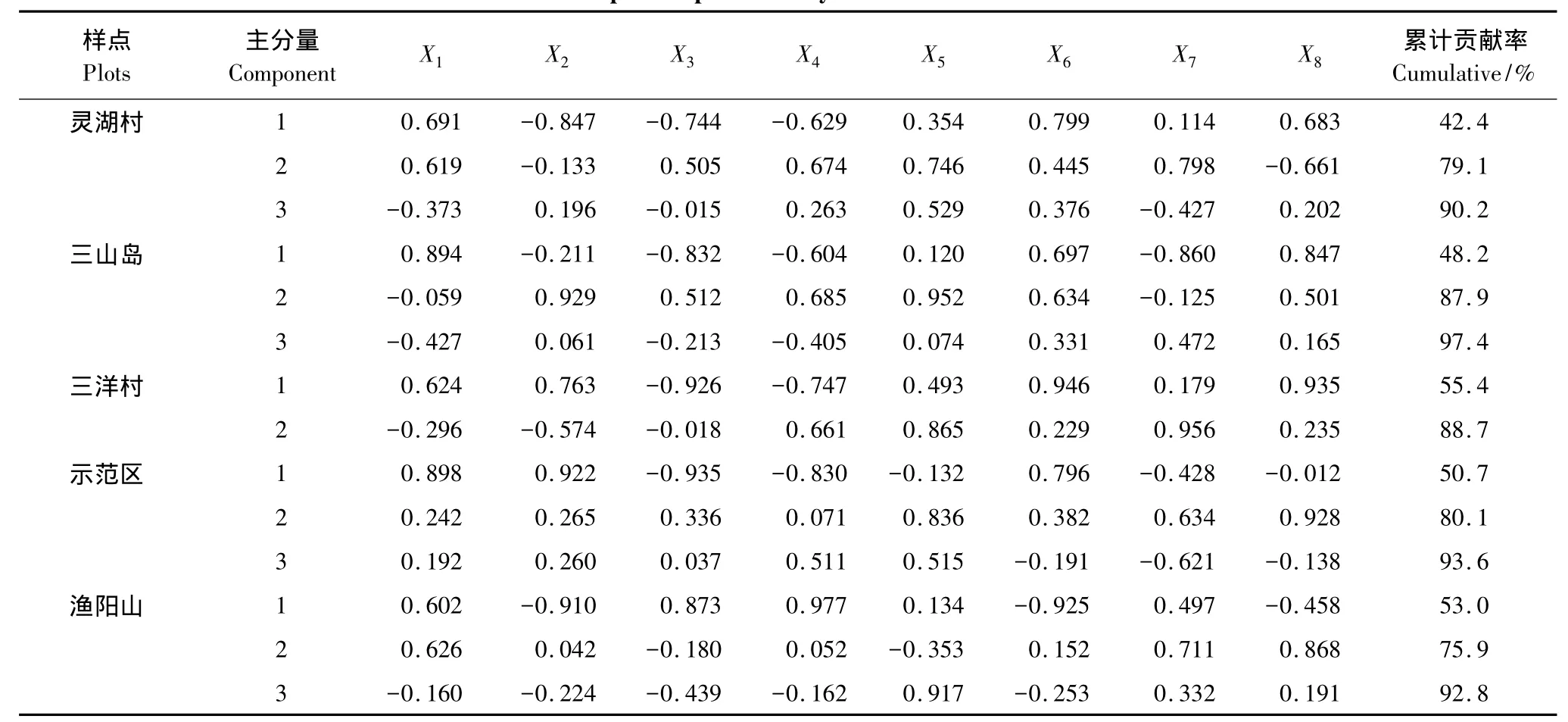
表6 各样点昆虫群落的主分量分析Table6 Principal component analysis of the insect communities
三山岛昆虫群落的第一主分量代表了捕食性昆虫的物种数、植食性昆虫的物种数以及中性昆虫的物种数和个体数的综合因子;第二主分量代表了捕食性昆虫的个体数和寄生性昆虫的物种数的综合因子;第三主分量代表了捕食性昆虫物种数和中性昆虫物种数的综合因子。3个主分量中第一主分量的贡献率最高为48.2%,说明三山岛昆虫群落的变化主要受捕食性昆虫物种数、植食性昆虫物种数以及中性昆虫物种数和个体数影响。
三洋村昆虫群落的第一主分量代表了植食性昆虫的物种数、寄生性昆虫的个体数和中性昆虫的个体数的综合因子;第二主分量代表了寄生性昆虫物种数和中性昆虫物种数的综合因子,说明了三洋村昆虫群落中捕食性昆虫和植食性昆虫对群落变化产生的影响较小,主导因子是寄生性昆虫和中性昆虫类群。
示范区昆虫群落第一主分量代表了捕食性昆虫的物种数、个体数和植食性昆虫物种数与个体数的综合因子;第二主分量代表了寄生性昆虫的物种数和中性昆虫的个体数的综合因子;第三主分量代表了植食性昆虫个体数以及寄生性昆虫物种数的综合因子。3个主分量的累计贡献率为93.6%,其中第一主分量的贡献率最高为50.7%,而第一主分量代表的是捕食性昆虫和植食性昆虫的综合因子,所以可说明捕食性昆虫和植食性昆虫主导了示范区的昆虫群落变化,寄生性昆虫和中性昆虫对群落的影响较弱。
渔阳山昆虫群落的前3个主分量累计贡献率为92.8%。第一主分量代表的是捕食性昆虫个体数与植食性昆虫物种数和个体数,以及寄生性昆虫个体数的综合因子;第二主分量代表的是中性昆虫个体数因子;第三主分量代表了寄生性昆虫个体数因子,其中第一主分量的贡献率最高达53%。从各因子的相关系数可以看出渔阳山昆虫群落中捕食性昆虫和植食性昆虫以及寄生性昆虫的个体数量要比他们的物种数量更加重要,昆虫群落变化的主导因子也集中在捕食性、植食性以及寄生性昆虫的个体数量上。
根据以上主分量分析的结果可知,影响灵湖村昆虫群落、三山岛昆虫群落、示范区昆虫群落以及渔阳山昆虫群落变化的主导因子集中在捕食性和植食性类群上;在三洋村昆虫群落中寄生性和中性类群的重要性要比植食性和捕食性类群高。
2.5 昆虫群落的排序
本研究采用极点排序法,利用昆虫群落的相异性系数(表7),计算出各群落的相关数值[20](表8),并对各样点昆虫群落进行排序。

表7 各样点昆虫群落的相异性系数Table7 The coefficient of dissimilarity of insect communities
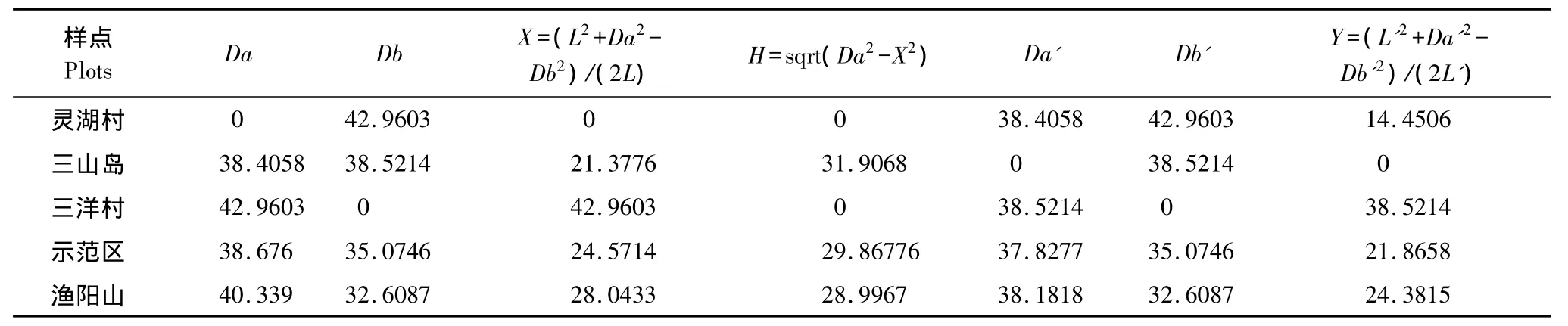
表8 各样点昆虫群落排序指标值Table8 The indices of ordination of insect communities
从图3的排序结果可以看出,示范区和渔阳山的昆虫群落接近,而且两者与其它3个样点都相距较远,昆虫群落差异大;灵湖村、三山岛和三洋村彼此之间的距离也都很大,昆虫群落的差异大。
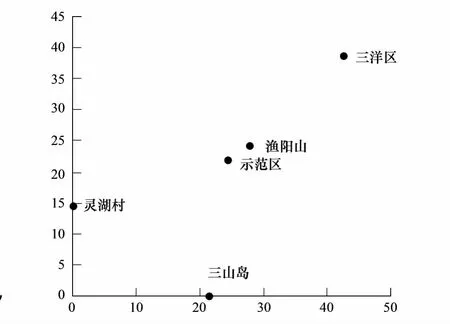
图3 各样点昆虫群落极点排序Fig.3 Polar ordination of insect communities
2.6 昆虫群落的稳定性分析
昆虫群落的相对稳定性一直以来受到群落生态学家的关注,对于其表述和成因也是各有不同[21-23]。本文中用 St/Si表示昆虫群落物种数和个体数的比值;用Sn/Sp表示昆虫群落中天敌类群种数和植食类群种数的比值。St/Si实际上是平均每物种个体数的倒数,可以反映种间数量上的制约作用,文中将各月份的物种数和个体数取平均值进行计算,这样就可以反映昆虫群落全年种间数量上的变化;Sn/Sp可以反映群落内部食物网的复杂程度以及相互制约的程度。从表9中可以看出,各样点之间的St/Si和Sn/Sp序列之间是存在差异的;St/Si的比值序列是渔阳山>示范区>灵湖村>三洋村>三山岛,渔阳山和示范区的比值明显高,说明这2样点昆虫群落物种数较多而个体数量少,昆虫群落没有突发性;Sn/Sp的比值序列是灵湖村>三山岛>渔阳山>三洋村>示范区,示范区的比值最小,说明样点内天敌昆虫所占的比例低,群落内部的网络关系不如灵湖村的复杂,从而使该样点昆虫群落降低了对外界扰动的缓冲能力。从总体来看渔阳山和灵湖村样点昆虫群落的稳定性相比其它3个样点较高。
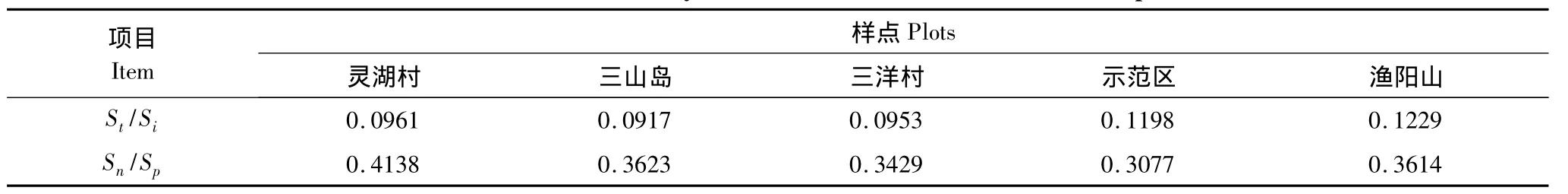
表9 不同样点昆虫群落的相对稳定性数值Table9 The relative stability value of insect communities in different plots
3 结论与讨论
太湖湿地共有318种昆虫,隶属于13目88科301属。昆虫群落物种丰富,结构复杂,优势类昆虫为半翅目、鳞翅目、鞘翅目、双翅目以及蜻蜓目等。因太湖流域属季风气候区,降水丰沛,植被繁茂,这就为以植物为食的半翅目、鳞翅目提供了优良的生存环境,使其种群得到发展壮大;湿地的水环境又为双翅目和蜻蜓目昆虫的生存提供了条件。植食性昆虫类群的繁荣也为捕食性昆虫和寄生性昆虫以及湿地其它动物提供了良好的营养基础,是湿地生物群落稳定和形成复杂结构的基础。
昆虫群落多样性特征指数是用来测定群落组织水平的重要指标,它不但反映了群落中物种的富集度、变异程度和均匀度等,而且还在不同程度上反映了不同地理、自然环境以及群落的发展状况[24,14]。太湖湿地各样地中示范区的昆虫群落多样性指数最高,渔阳山次之,而且两者明显高于其它样地;同时,两者的丰富度指数和均匀度指数也偏高,这是样地环境对昆虫群落综合作用的结果,也反映了两样地昆虫群落的稳定性相对较高。灵湖村昆虫群落的多样性指数最低,但群落物种丰富度最高,均匀度最低,说明群落物种多样性和物种丰富度与均匀度的关系复杂。
从时间序列来看,多样性指数和均匀度指数的变化趋势是一致的,而且相关性分析表明二者呈正相关,且相关性很强(相关系数r=0.792-0.99,P<0.01);多样性指数与丰富度指数虽然也呈正相关,但相关性低(相关系数r=0.249-0.874).多数学者认为群落多样性指数和均匀度指数是否保持一致,可以说明群落的稳定性[9,4]。贺达汉[25]等对荒漠昆虫群落的研究中发现群落多样性和均匀度是一致的,认为群落结构是稳定的;尤平[1]等对天津湿地蛾类群落的研究中发现团泊洼和北大港水库的群落多样性和均匀度不一致,表明了群落环境的不稳定性。本文认为太湖湿地昆虫群落的稳定性需要通过多个指标的综合反映才能确定。
各样点环境条件存在差异,从而影响了在环境中生存的昆虫群落的属性。通过极点排序,可以直观的看到渔阳山和示范区昆虫群落的距离最近,其它各样点分布在周围,且相互之间距离较大,这都表明各样点昆虫群落的属性差异大,这是各样点环境综合作用的结果。因此,昆虫群落的极点排序也是对各样点环境条件关系的直观反映,从而也就说明了渔阳山和示范区的环境状况是最接近的,其它样点的环境状况差异明显。尤平[1]等在研究天津湿地蛾类群落时利用极点排序法对群落属性进行了分析,研究也表明了昆虫群落的极点排序结果可以直观的反映样点间的环境条件关系。顾伟[9]等则是利用聚类分析对扎龙湿地不同生境昆虫群落的属性进行了分析,但分析结果不如极点排序二维坐标直观。
昆虫群落的结构组成可以对昆虫群落的稳定性产生影响。太湖湿地昆虫群落的主分量分析表明只有三洋村昆虫群落的主导因子集中在寄生性和中性昆虫类群上,这也说明了太湖湿地生境中昆虫群落主要受捕食性和植食性类群的影响,也反映了太湖湿地昆虫群落结构比较复杂,对外界干扰的缓冲能力和自我调节能力较好,能够维持湿地昆虫群落的相对稳定性。顾伟[9]和张晓明[26]等在各自的研究中均认为影响昆虫群落的主导因子为捕食性昆虫类群时,群落稳定性高。
通过对各样点昆虫群落多样性特征指数、特征指数随时间的变化、群落主分量以及群落稳定性的综合分析之后认为,各昆虫群落相对稳定性的大小为:渔阳山>示范区>三山岛>灵湖村>三洋村。渔阳山昆虫群落物种丰富;在时间序列上多样性指数表现稳定;群落变化的主导因子集中在捕食性和植食性类群;群落物种数与个体数的比值St/Si和天敌类群种数与植食类群种数的比值Sn/Sp均偏高,这都表明了渔阳山昆虫群落结构复杂相对稳定性高。示范区样点的稳定性仅次于渔阳山,说明湿地恢复工程对太湖湿地生物的保护效果明显。三洋村样点由于植被类型相对简单,人为干扰多,致使昆虫群落多样性指标偏低,各指标在时间序列上波动大;群落中寄生性和中性类群是影响群落的主导因子;St/Si和Sn/Sp比值偏低,最终导致群落结构稳定性相对最低。
湿地植被和人工干扰是影响太湖湿地昆虫群落的重要因子,关于它们与太湖湿地昆虫群落定量的相关性以及昆虫群落内部的竞争与平衡机制都有待于进一步的研究。
[1] You P,Li H H.Species richness and diversity of moth communities in Tianjin Wetland;implications for environmental management.Acta Ecologica Sinica,2006,26(3):629-637.
[2] Zhong Y X,Fu B Q.Preliminary analysis on composition features of insect community in Baihe River wetland,Beijing in Summer.Journal of Capital Normal University:Natural Science Edition,2007,28(3):70-77.
[3] Liu P,Zhong Y X,Fu B Q,Chou X D,W H.Diversity and ecological distributions of Hymenoptera communities in Yeyahu Wetland,Beijing.Journal of Capital Normal University:Natural Science Edition,2008,29(2):38-44.
[4] Song W J,Ma L,Wang H,Han X B.Diversity of insect communities by the lakeside in Zhalong Nature Reserve.Journal of Northeast Forestry University,2007,35(7):80-81.
[5] Bao X,LǚX G,Zhang F.Insect diversity in different forests near the annular wetland of Sanjiang Plain.Journal of Arid Land Resources and Environment,2010,24(2):137-140.
[6] Bao X,LǚX G,Zhang F.Biodiversity and seasonal change of in sects in annular wetlands in Sanjiang Plain.Journal of Northeast Forestry University,2009,37(5):100-101.
[7] Wang X P,Yang F,Yang G J.Composition and diversity of in sect community in yellow river wetland of Yinchuan.Sichuan Journal of Zoology,2008,27(5):827-831.
[8] Peng X W,Go H,Dong H Q,Wu Q H.Study on diurnal insect communities in different habitats at the Jiuduansha wetland national nature reserve.Journal of Fudan University:Natural Science Edition,2006,45(6):784-789.
[9] Gu W,Ma L,Ding X H,Zhang J,Han Z J.Insect diversity of different habitat types in Zhalong Wetland,Northeast China.Chinese Journal of applied Ecology,2011,22(9):2405-2412.
[10] Bedford A P,Powell I.Long-term changes in the invertebrates associated with the litter of Phragmites australis in a managed reed-bed.Hydrobiologia,2005,549(1):267-285.
[11] Kati V,Mani P,von Helversen O,Willemse F,Elsner N,Dimopoulos P.Human land use threatens endemic wetland species:the case of Chorthippus lacustris(La Greca and Messina 1975)(Orthoptera:Acrididae)in Epirus,Greece.Journal of Insect Conservation,2006,10(1):65-74.
[12] Hao SG,Zhang X X,Chen X N,Luo Y J,Tian X Z.The dynamics of biodiversity and the composition of nutrition classes and dominant guilds of arthropoda community in paddy field.Acta Entomologica Sinica,1998,41(4):343-353.
[13] You P,Li H H,Wang SX,Xu JS.Diversity of the moth community in the Qilihai wetland,Tianjin.Acta Entomologica Sinica,2003,46(5):617-621.
[14] Sun R Y.Principles of animal ecology.3rd edition.Beijing:Beijing Normal University Press,2001.
[15] Zhang C Z,Yao L Z,Li Q,Li Z Y,Song J X,Ruan Y Z.Insect community structure and its dynamics in Chinese prickly ash plantations at different altitudes in Yunnan Province.Chinese Journal of Applied Ecology,2006,17(5):915-919.
[16] Liu Y H,Ning Z R,Liu Y.Temporal and spatial structure of carabid community in agricultural landscape of Dongbeiwang,Beijing.Chinese Journal of Applied Ecology,2004,15(1):85-90.
[17] Gao B J,Shen SG,Wang Z W,Zhang Y H.Studies on the temporal structure and dynamis of the insect communities in gardens.Acta Ecologica Sinica,1998,18(2):193-197.
[18] Gao B J,Zhang Z Z,Li Z Y.Studies on the influence of the closed forest on the structure,diversity and stability of insect community.Acta Ecologica Sinica,1992,12(1):1-7.
[19] Liu W P,Deng H L.The butterfly diversities in Muli,Sichuan province.Acta Ecologica Sinica,1997,17(3):266-271.
[20] Zhao Z M,Guo Y Q.Principle and Methods of community Ecology.Chongqing:Publishing House of Scientific and Technological Documentation,Chongqing Branch,1990:134-160.
[21] Wang SF,Lei G C.Preliminary study on structure and succession of canopy insect community in masson pine forest.Journal of Central-South Forestry College,1986,6(1):1-11.
[22] Ren L Z,Wang SF.A study on the insect community and it's structure of time and space in stands of pinus massoniana.Forest Research,1988,1(4):398-403.
[23] Goodman D.The theory of diversity-stability relationships in ecology.The Quarterly Review of Biology,1975,50(3):237-266.
[24] Zhao Z,Ren BZ,Xi GS.A preliminary study on insect biodiversity of Zhuanshan lake area in Siping.Chinese Journal of Ecology,2002,21(3):65-68.
[25] He D H,Tian C,Ren G D,Hao F M,Ma S Y.A preliminary study on the community structure of desert forage grassland.Chinese Journal of Grassland,1988,(6):24-28.
[26] Zhang X M,L Q,Chen G H,Yang J,Gao X,Song JX.Insect community structure and its stability in a Zanthoxylum bungeanum garden with different planting pattern.Chinese Journal of Applied Ecology,2009,20(8):1986-1991.
参考文献:
[1] 尤平,李后魂.天津湿地蛾类丰富度和多样性及其环境评价.生态学报,2006,26(3):629-637.
[2] 仲雨霞,付必谦.北京白河湿地夏季昆虫群落组成特征的初步分析.首都师范大学学报:自然科学版,2007,28(3):70-77.
[3] 刘萍,仲雨霞,付必谦,仇雪东,王昊.北京野鸭湖湿地膜翅目群落多样性及生态分布.首都师范大学学报:自然科学版,2008,29(2):38-44.
[4] 宋文军,马玲,王慧,韩小兵.扎龙自然保护区湖泊边昆虫群落多样性.东北林业大学学报,2007,35(7):80-81.
[5] 暴晓,吕宪国,张帆.三江平原典型湿地不同林型昆虫物种多样性研究.干旱区资源与环境,2010,24(2):137-140.
[6] 暴晓,吕宪国,张帆.三江平原环形湿地昆虫种类多样性与季节动态.东北林业大学学报,2009,37(5):100-101.
[7] 王新谱,杨锋,杨贵军.黄河湿地银川段昆虫群落组成及其多样性研究.四川动物,2008,27(5):827-831.
[8] 彭筱葳,高慧,董慧琴,吴千红.九段沙湿地国家自然保护区不同生境中昼行性昆虫群落研究.复旦学报:自然科学版,2006,45(6):784-789.
[9] 顾伟,马玲,丁新华,张静,韩争伟.扎龙湿地不同生境的昆虫多样性.应用生态学报,2011,22(9):2405-2412.
[12] 郝树广,张孝義,程遐年,罗跃进,田学志.稻田节肢动物群落营养层及优势功能集团的组成与多样性动态.昆虫学报,1998,41(4):343-353.
[13] 尤平,李后魂,王淑霞,徐家生.天津七里海湿地蛾类多样性.昆虫学报,2003,46(5):617-621.
[14] 孙儒泳.动物生态学原理(第三版).北京:北京师范大学出版社,2001:394-403.
[15] 张承祚,姚利忠,李强,李正跃,宋家雄,阮应珍.云南不同海拔花椒园昆虫群落结构及动态.应用生态学报,2006,17(5):915-919.
[16] 刘云慧,宁振荣,刘云.北京东北旺农田景观步甲群落结构的时空动态比较.应用生态学报,2004,15(1):85-90.
[17] 高宝嘉,申曙光,王正文,张玉会.园林昆虫群落时间结构及动态研究.生态学报,1998,18(2):193-197.
[18] 高宝嘉,张执忠,李镇宇.封山育林对昆虫群落结构及多样性稳定性影响的研究.生态学报,1992,12(1):1-7.
[19] 刘文萍,邓合黎.木里蝶类多样性的研究.生态学报,1997,17(3):266-271.
[20] 赵志模,郭依泉.群落生态学原理与方法.重庆:科学技术文献出版社重庆分社,1990:134-160.
[21] 王淑芬,雷光春.马尾松林冠层昆虫群落结构及演替研究初报.中南林学院学报,1986,6(1):1-11.
[22] 任立宗,王淑芬.马尾松林昆虫群落及其时空结构的研究.林业科学研究,1988,1(4):398-403.
[24] 赵卓,任炳忠,奚耕思.四平转山湖水库地区昆虫多样性的初步研究.生态学杂志,2002,21(3):65-68.
[25] 贺达汉,田畴,任国栋,郝峰茂,马世瑜.荒漠草原昆虫的群落结构及其演替规律初探.中国草地,1988,(6):24-28.
[26] 张晓明,李强,陈国华,杨洁,高鑫,宋家雄.不同种植模式花椒园昆虫群落的结构及稳定性.应用生态学报,2009,20(8):1986-1991.
