中国刺参与韩国红刺参杂交及子代发育特性的研究
2013-07-12孙灵毅赵强任利华王有廷王亮贺加贝
孙灵毅,赵强、,任利华,王有廷,王亮,贺加贝
(1.烟台市水产研究所,山东烟台264003;2.山东省海洋水产研究所,山东烟台264006;3.烟台海益苗业有限公司,山东蓬莱265619)
中国刺参与韩国红刺参杂交及子代发育特性的研究
孙灵毅1,赵强1、3,任利华2,王有廷3,王亮1,贺加贝1
(1.烟台市水产研究所,山东烟台264003;2.山东省海洋水产研究所,山东烟台264006;3.烟台海益苗业有限公司,山东蓬莱265619)
分别以刺参Stichopus japonicus中国群体和韩国群体为母本,采用同步催产法,同时收集中国刺参和韩国红刺参的精子、卵子,进行正交 (中国刺参♀×韩国红刺参♂)和反交 (韩国红刺参♀×中国刺参♂),比较两者的受精率,同时对正交受精及早期胚胎发育过程进行连续光镜和荧光显微镜观察。结果表明:中国刺参和韩国红刺参正交和反交的受精率分别为91%和94%;韩国红刺参的精子可以穿过中国刺参的卵膜进行受精,并激活卵子减数分裂使其释放第一极体 (PB1)和第二极体 (PB2),然后受精卵开始进行卵裂;杂交胚胎发育正常,正交和反交的孵化率分别为75%和90%。另外,在试验中发现有多精入卵现象。
中国刺参;韩国红刺参;杂交;受精;胚胎发育
刺参Stichopus japonicus为温带种类,在中国主要分布于辽宁、河北、山东等沿海水域,在朝鲜半岛、日本和俄罗斯远东沿海水域也有分布[1]。近年来,刺参养殖规模不断扩大,由于养殖技术滞后,加之刺参自然资源匮乏,种参质量难以保障。养殖所用亲参没有经过系统的选育和种质改良,种质的混杂和小量亲本累代繁育造成的种质退化,使原有种群的优良性状不能得以保持,出现了生长速度降低、抗病抗逆性能下降等诸多问题。通过引进新品种,积极开展优良品种的遗传育种工作,是尽快解决刺参种质退化的有效途径。
杂交育种是水产动物育种的重要途径。虽然现代生物技术在育种领域中已得到广泛运用,但杂交育种具有操作简便并能综合双亲优良性状等特点,仍是国内外普遍运用而且育种效果比较明显的一种育种技术[2-8]。韩国红刺参与中国刺参是刺参的两个不同地理种群,由于生活环境的不同,韩国红刺参具有夏眠时间短、生长速度快、体壁厚实、体色鲜艳的特点。本研究中,作者以中国刺参和韩国红刺参为母本进行杂交,在光镜和荧光显微镜下对其受精和第一次卵裂过程中核相变化进行观察,并对杂交后代的生长性状进行分析和判断,旨在发挥韩国红刺参的品种优势,达到缩短养殖周期、提高养殖设施利用率、降低养殖成本和增加经济效益的目的,为刺参杂交育种和养殖生产提供科学依据。
1 材料与方法
1.1 材料
试验于2011年5月上旬在烟台市水产研究所科研基地——烟台海益苗业有限公司育苗场进行。试验用韩国红刺参 (简称红刺参)于2011年1月从韩国济州岛沿海收集运至烟台,规格每只为200~300 g,采取沥水无氧运输方式,运输温度为7~8℃,运输时间长达 10 h以上,存活率为100%。中国刺参 (简称刺参)取自烟台市莱山区前七夼海区,暂养于室内强化促熟,用于繁殖。
1.2 方法
1.2.1 亲参的促熟培养 将引进的45只红刺参,在烟台海益苗业有限公司育苗场进行越冬培养、生殖调控和促熟培育。
1)蓄养条件。蓄养池为12 m3,水深为1~1.2 m,水温为7~18℃,盐度为29~31,pH为7.8~8.2,光照为500 lx,溶解氧为5 mg/L以上。
2)饵料投喂与水质调节。根据亲参的摄食状况以及水温变化,按参体质量的3%~6%每日分早晚2次投喂鼠尾藻粉、配合饲料以及海泥。早投1/3,晚投2/3。当部分个体开始出现排放现象时停止投喂。每天早晚各换水1次,每次换水量为1/3~1/2,换水温差不超过0.5℃。发现死亡个体及时拣出。每隔3 d排干池水1次,彻底清除池底排泄物和残饵。
1.2.2 试验设计 试验设正交组 (刺参♀×红刺参♂)、反交组 (红刺参♀×刺参♂)和自交 (刺参♀×刺参♂)3组。
1)亲参催产和人工受精。2011年5月4日,有2只雄性红刺参个体出现少量排放,表明其性腺发育基本成熟。此时,停止饲料投喂,并将成熟个体移至容积为12 L的塑料桶中单独饲养。同时,每天选取200只待产的刺参,以备排放时分别拣出,与红刺参同步获取精液和卵子。5月12日晚,对刺参和红刺参同步进行阴干、流水刺激催产。红刺参与刺参均出现集中排放现象,排放时水温为18.2℃,排放持续时间约2 h。
2)显微观察。受精过程开始时,在1、2、4、6、8 min等时间段分别取样1次;10~60 min,每10 min取样1次;60~120 min,每隔10~20 min取样1次,每次取卵不少于1 000粒。
普通光镜观察的样品直接滴于载玻片上,在光镜下观察并拍照。另将所取样品用提前配制好的体积分数为4%的多聚甲醛液固定,保存于冰箱 (4℃)中,用于荧光显微观察。荧光显微镜观察的样品先用0.1 mol/L磷酸缓冲液滴 (pH 7.2)冲洗2~3次,然后用一次性移液管转到3 mL离心管中,加入配好的Hoechst33258荧光染料 (购自北京泛博生物化学有限公司),在黑暗环境下染色5~10 min后,滴于载玻片上,加盖玻片,经轻微压片后在NikonE600W荧光显微镜的紫外光 (350 nm)下观察,并用佳能G10拍照。每个样品观察均重复3次。
1.3 数据处理
在稚参期 (60、75、90、105日龄)随机抽取30只个体,测量体长,称取体质量。用Excel软件统计各组的体长和体质量,数据均以平均值±标准差表示。
2 结果与分析
2.1 正交受精及早期胚胎发育的显微观察
2.1.1 卵子的形态 成熟的卵子 (图1-A)呈圆形,直径为105~130 μm,卵质均匀。在荧光显微镜下被染成亮蓝色,卵质为浅蓝色。
2.1.2 受精 在光镜下清晰可见,精子围绕在卵子周围 (图2-A、B)。视野中,大部分精子可入卵,入卵的位置是随机的,随后精子发生顶体反应,头部逐渐进入胞质 (图1-B)。刺参的卵子和红刺参的精子受精后在受精卵的四周有受精膜举起(图1-C)。受精10~13 min后,在卵子的动物极出现第一极体 (图1-D,图2-C),再过20~25 min出现第二极体 (图2-D、E)。
2.1.3 卵裂 在20~22℃水温下进行培养,大部分杂交参的卵裂是比较典型的辐射型全等裂,但也有部分受精卵的形状发生改变,呈椭圆形 (图1-L)。卵子在受精后1 h左右开始进行第一次分裂(图1-E,图2-F),其分裂面通过卵轴,分裂成2个大小完全相等的细胞 (图1-F、G,图2-G、H)。受精后2 h左右进行第二次分裂 (图2-I),其分裂面仍与卵轴平行,并和第一次分裂面垂直,形成4细胞 (图1-H、I,图2-J、K)。受精后2 h 20 min左右进行第三次分裂,其分裂面位于卵子赤道附近并与卵轴垂直,形成大小完全相等、排列为上下二层 (每层为4个细胞)的8个细胞 (图1-J)。而后受精卵继续分裂,形成16个细胞 (图2-L),细胞数量呈几何级数增长。受精后4 h 20 min,胚胎细胞慢慢地形成中空的囊胚 (图2-M)。当囊胚的植物极变平,所形成的原肠原基开始内凹,内凹的程度由浅变深,此时细胞发育进入原肠期 (图2-N、O)。另外,还发现有多精入卵现象(图1-K)。
2.2 杂交的受精率及胚胎发育进程
在显微镜下,对受精卵的胚胎发育计数得到,刺参和红刺参正、反交的受精率分别为91%和94%,而刺参自交组受精率达到99%以上。通过对亲本自交组与杂交组的受精率及早期胚胎发育进程的观察和比较,结果显示,反交组受精卵的早期胚胎发育进程与自交组几乎同步,而正交组在时间上却有着明显的延迟。正交组受精卵出现第一极体的时间普遍较自交组和反交组推迟了3~6 min,而出现第一次卵裂的时间则平均推迟了5~10 min。在同样温度条件下,杂交组受精卵的孵化时间均比自交组延长0.5~1.0 h(表1)。
2.3 正交与反交组受精卵孵化率的比较
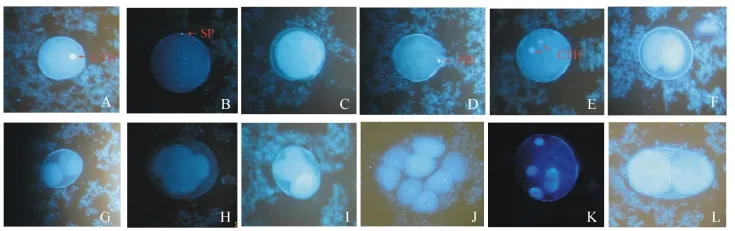
图1 中国刺参♀×韩国红刺参♂受精及早期胚胎发育过程的荧光观察Fig.1 Cytological fertilization process and early embryonic development in the cross Chinese sea cucumber♀×Korean red sea cucumber♂under fluorescent microscope

图2 中国刺参♀×韩国红刺参♂受精及早期胚胎发育过程的光镜观察Fig.2 Cytological fertilization process and early embryonic development in the cross Chinese sea cucumber♀×Korean red sea cucumber♂under light microscope
催产过程中,红刺参12只排放 (♀5只,♂7只)、刺参89头排放 (♀37只,♂52只)。按照杂交设计方案,获得反交受精卵约240万粒,初孵幼体约216万只,孵化率约为90%;正交受精卵约9 500万粒,初孵幼体约7 125万只,孵化率约为75%;自交受精卵约2 400万粒,初孵幼体约2 360万只,孵化率约为98.5%。

表1 各组胚胎及幼体发育历程(20~22℃)Tab.1 The embryonic and larval development of the sea cucumber Stichopus japonicus in three experimental groups (20-22℃)
2.4 杂交与自交幼体生长情况的比较
经过 105 d的苗种培育,稚参 60、75、90、105日龄的体长、体质量的生长情况见图3。课题阶段性验收时,随机选取了30只幼体进行体长测量和称重。结果显示,自交组幼参的体长为4~16 mm,平均体长为9.1 mm,平均体质量为20 mg;正交组幼参的体长为5~15 mm,平均体长为9.8 mm,平均体质量为25.2 mg;反交组幼参的体长为7~14 mm,平均体长为10.8 mm,平均体质量为33.4 mg。反交组幼参的生长优势明显。

图3 杂交与自交稚参的生长Fig.3 The growth of juveniles of inbreed sea cucumber and hybrids
3 讨论
3.1 刺参与红刺参杂交可行性的分析
在水产苗种培育领域,杂交育种在新品系的建立、品种改良等生产过程中发挥着越来越重要的作用,而棘皮动物的杂交胚胎研究尚处于起步阶段。江曙光等[9]选择遗传差异大的中国刺参和俄罗斯远东海域刺参进行杂交,通过多种杂交组合筛选出杂交优势明显的子一代;胡美燕等[10]对中国刺参与日本红刺参杂交子一代的早期生长进行了研究,仅在杂交受精率、孵化率方面作了试验比较,但未涉及杂交子代胚胎发育的细胞学。而在贝类方面,相关方面的研究较多,如王卫军等[11]进行了栉孔扇贝和长牡蛎杂交受精及早期胚胎的细胞学研究,董迎辉等[12]进行了文蛤受精及其早期胚胎发育过程的细胞学观察,孙振兴等[13]进行了皱纹盘鲍与盘鲍杂交效果的分析。中国刺参与韩国红刺参的杂交不仅可以丰富棘皮动物杂交品种,同时还在刺参杂交子代胚胎发育研究方面填补了空白。
本研究中通过光学显微镜和荧光显微镜观察刺参和红刺参两个不同地理种群的受精和早期胚胎发育过程,以及受精卵和孵化率的统计数据,推断两者的杂交是可行的。显微观察显示,精子和卵子能够正常受精,精子的入卵激活卵子进行减数分裂,从而排出第一极体和第二极体,经过卵裂期、囊胚期、原肠期,进而孵化为初耳幼虫,完成杂种早期胚胎发育进程。虽然发育时间较亲本对照组有延迟,但杂交组的正交、反交受精率分别达到91%和94%,孵化率分别达到75%和95%。说明两者之间的大规模杂交育种是可以进行的。
在荧光显微观察中,发现有多精入卵 (图1-K)和细胞发育呈椭圆形 (图1-L)等特别现象出现。多精入卵现象在贝类杂交中普遍存在,朱东丽等[14]在泥蚶♀×毛蚶♂杂交中也发现了多精入卵现象。本试验结果表明,同种参自交时,单个精子入卵后可以阻止其他精子的再入卵,因而很少发生多精入卵现象。而在杂交过程中,单个红刺参精子可以进入并激活刺参卵子,但精卵异质使入卵的精子并不能阻止其他精子进入,从而出现多精入卵现象,以致于细胞异常分裂而使胚胎发育终止。这也是杂交受精率低于正常自交的原因之一。但目前对于过量异质精子入卵造成细胞分裂紊乱以致发育中断的机理尚不清楚,还有待进一步研究和探讨。
3.2 刺参与红刺参杂交子代的杂种优势评估
杂交育种可以综合亲本的优良性状,增加变异性,并且杂交子代在成活率、生长速度、抗逆性等方面表现出一定的杂种优势。种间杂交虽然能够产生大的遗传变异,但由于技术难度高,很容易出现杂交不亲和。很多杂交研究中,出现幼体发育到一定阶段就停止而死亡的现象。相比之下,同种的不同品系或不同地理群体间的同种远交则能克服杂交后代生殖、发育和生长的障碍,在生长发育和成活率方面表现出一定的优势。目前,国外有关方面的研究主要集中在牡蛎、扇贝以及一些鱼类上[15-19];国内对马氏珠母贝 Pinctada martensii、皱纹盘鲍Haliotis discus hannai、栉孔扇贝Chlamys farreri、杂色鲍H.diversicolor supertext等开展了不同地理群体的杂交[20-29]。
基因异质性是杂种优势后代表现的根本原因,本试验中所用的刺参与红刺参作为不同的地理种群,表现出各自不同的性状特质。从图3可以看出,杂交子代经过105 d的生长发育,表现出一定程度的杂种优势。从试验结果看,杂交使群体的生长得到改良,反交组的生长优势明显,高于正交组和自交组。正、反交子代在杂种优势上表现不同的原因可能是由于胚胎发育初期,母本所携带的基因片段发挥作用所导致。刘小林等[24]在栉孔扇贝不同地理种群杂交效果的研究中也提出了母性效应对早期胚胎发育影响的理论。这一理论的确定还有待于今后其他品种杂交结果的证实。杂种优势或杂种活力的存在,能显著改善和增强亲本的优势性状,表现在生活力强、适应性广,有较强的抗逆力和竞争力,从而在大规模生产中起到不可低估的作用。
尽管正、反交受精卵在胚胎发育过程中并没有表现出幼体变态快的特点,甚至正交组幼体的变态还有所延迟,但在生长到稚参后,杂交组的生长明显加快。在105日龄时,杂交优势表现明显,这与红刺参具有适温范围广、生长速度快、夏眠时间短的生理特性相关。红刺参作为父本和母本的正、反交,其子代能否继承亲代的夏眠时间短、抗病性强、出肉率高的特点,还有待进一步探讨和研究。
[1]廖玉麟.中国动物志棘皮动物门海参纲[M].北京:科学出版社,1997:148-150.
[2]刘小林,常亚青,相建海,等.栉孔扇贝不同种群杂交效果的研究:Ⅱ.中国种群和俄罗斯种群及其杂种 F1中期生长发育[J].海洋学报,2005,27(2):135-140.
[3]杨璞,杨爱国,单伟华,等.栉孔扇贝(♀)×虾夷扇贝♂杂交子代胚后发育过程中遗传构成变化研究[J].渔业科学进展, 2009,30(2):66-70.
[4]吴仲庆.水产生物遗传育种学[M].3版.厦门:厦门大学出版社,2000.
[5]张国范,刘晓,阙华勇,等.贝类杂交及杂种优势理论和技术研究进展[J].海洋科学,2004,28(7):54-60.
[6]Okumura S I,Kimura K,Sakai M,et al.Chromosome number and telomere sequence mapping of the Japanese sea cucumber Apostichopus japonicus[J].J Fish Sci,2009,75(1):249-251.
[7]邱洋洋,陆海燕,宓华明,等.金鱼红白虎头和红白蝶尾及其杂交F1代的体色和体型的分析[J].大连海洋大学学报,2011, 26(3):209-214.
[8]刘毅辉,陈永乐,朱新平,等.翘嘴鳜、斑鳜及其杂交后代的胚胎和胚后发育比较[J].大连海洋大学学报,2012,27(1):6-11.
[9]江曙光,常亚青,田燚,等.仿刺参多种杂交方式下杂交效果的初步研究[J].大连水产学院学报,2009,24(3):247-250.
[10]胡美燕,李琪,孔令锋,等.中国刺参与日本红刺参杂交子一代的早期生长比较[J].中国海洋大学学报:自然科学版, 2009,39(S1):375-380.
[11]王卫军,杨爱国,刘志鸿,等.栉孔扇贝♀和长牡蛎♂杂交受精及早期胚胎的细胞学研究[J].海洋水产研究,2008,29(6): 78-83
[12]董迎辉,林志华,柴雪良,等.文蛤受精及其早期胚胎发育过程的细胞学观察[J].动物学报,2007,53(4):700-709.
[13]孙振兴,常林瑞,宋志乐.皱纹盘鲍与盘鲍杂交效果分析[J].水产科学,2005,24(8):1-3
[14]朱东丽,林志华,董迎辉,等.泥蚶(♀)×毛蚶♂受精及胚胎发育过程的初步研究[J].渔业科学进展,2010,31(5):69-75.
[15]Banks M A,McGoldrick D J,Borgeson W,et al.Gametic incompatibility and genetic divergence of Pacific and Kumamoto oysters Crassostrea gigas and C.sikamea[J].Mar Biol,1994,121:127-135.
[16]Patrik S,Arnaud H,Olivier L M,et al.A compairative field study of growth,survival and reproduction of Crassostrea gigas,C.angulata and their hybrids[J].Aquat Living Resour,2002,15:243-250.
[17]滕爽爽,李琪,孔令峰.岩牡蛎和长牡蛎杂交的受精细胞学观察[J].中国海洋大学学报,2009,39(9):338-342.
[18]桂建芳,梁绍昌,朱蓝菲,等.鱼类远缘杂交正反交杂种胚胎发育差异的细胞遗传学分析[J].动物学研究,1993,14(2): 171-177.
[19]李炎路,陈超,翟介明,等.鱼类杂交育种技术及其在石斑鱼类中的应用[J].海洋渔业,2010,34(1):102-109.
[20]王爱民,王嫣,顾志峰,等.马氏珠母贝(Pinctada martensii)2个地理群体杂交子代的杂种优势和遗传变异[J].海洋与湖沼,2010,41(1):140-146.
[21]张国范,王继红,赵洪恩,等.皱纹盘鲍中国群体和日本群体的自交与杂交F1的RAPD标记[J].海洋与湖沼,2002,33 (5):484-491.
[22]常亚青,刘小林,相建海,等.栉孔扇贝中国种群与日本种群杂交子一代的早期生长发育[J].水产学报,2002,26(5):385-390.
[23]Liu X L,Chang Y Q,Xiang J H.Hybridization effects of three populations of Chlamys farreri from China,Korea,and Japan[J].J Shellfish Res,2004,23(2):501-505.
[24]刘小林,常亚青,相建海,等.栉孔扇贝不同种群杂交效果的初步研究:Ⅰ.中国种群与俄罗斯种群的杂交[J].海洋学报, 2003,25(1):93-99.
[25]常亚青,刘小林,相建海,等.栉孔扇贝不同种群杂交效果:Ⅲ.中国种群与俄罗斯种群1~2龄的生长发育[J].海洋学报,2006,28(2):114-120.
[26]Zheng H P,Zhang G F,Guo X M,et al.Heterosis between two stocks of the bay scallop Argopecten irradians irradians Lamark (1819)[J].J Shellfish Res,2006,25(3):807-812.
[27]Liu X L,Chang Y Q,Xiang J H,et al.Studies on hybridization effects of different geographic populations of Chlamys farreri:Ⅱ. The medius term growth and development of Chlamys farreri populations from China and Rssia and their reciprocal crosses[J].Acta Oceanologica Sinica,2003,22(2):255-263.
[28]游伟伟,柯才焕,蔡明夷,等.杂色鲍日本群体与台湾群体杂交效果的初步研究[J].厦门大学学报:自然科学版,2005,44 (5):701-705.
[29]石耀华,王嫣,曲艳波,等.杂色鲍野生群体与养殖群体自交和杂交子一代的RAPD分析[J].海洋水产研究,2007,28(4): 47-52.
Crossing and offspring development of sea cucumber Stichopus japonicus between Chinese and Korean populations
SUN Ling-yi1,ZHAO Qiang1,3,REN LI-hua2,WANG You-ting3,WANG Liang1,HE Jia-bei1
(1.Yantai Fisheries Research Institute,Yantai 264003,China;2.Shandong Marine Fisheries Research Institute,Yantai 264006,China; 3.Yantai Haiyi Seed-production Co.Ltd.,Penglai 265619,China)
The sperm,and eggs of sea cucumber Stichopus japonicus derived from China and Korea were collected by synchronous spawning induction,and direct cross and reciprocal crosses were conducted between the two populations of sea cucumber.The fertilization was observed in the reciprocal crosses and fertilization and early embryonic development were successively observed in the direct cross between Chinese sea cucumber♀×Korean red sea cucumber♂under light microscopy and fluorescence microscopy.The sperm of Korean red sea cucumber was found to attach to the egg surfaces of Chinese sea cucumber rapidly and the acrosomal reaction was observed successfully. The inerspecifc sperm activated the meiosis of Chinese sea cucumber egg and induced the releasing of polar body 1 (PB1)and polar body 2(PB2),and then the male and female pronuclei fused into zygotonuceus and the cleavage began subsequently.The fertilization rate was found to be 91%in the direct cross between Chinese sea cucumber♀×Korean red sea cucumber♂and 94%in the reciprocal crosses(Korean red sea cucumber♀×Chinese sea cucumber♂).Meantime,normal embryonic development was observed,with hatching rate of 75%in the direct cross and 90%in the reciprocal cross.In addition,polyspermy was observed in this experiment.
Chinese sea cucumber;Korean red sea cucumber;hybridization;fertilization;embryogenesis
S968.9
A
2012-08-15
山东省科技发展计划项目 (2010GHY10519)
孙灵毅 (1963-),女,高级工程师。E-mail:sunly1963@163.com
2095-1388(2013)03-0281-06
