2012年秋冬季利文斯顿岛南极磷虾渔业CPUE指标变动及其影响因素
2013-07-02朱国平朱小艳孟涛赵勇徐怡瑛吴继魁许柳雄
朱国平,朱小艳,孟涛,赵勇,徐怡瑛,吴继魁,许柳雄
(1.上海海洋大学海洋科学学院,上海201306;2.国家远洋渔业工程技术研究中心,上海210306;3.大洋渔业资源可持续开发省部共建教育部重点实验室,上海201306;4.上海海洋大学食品学院,上海201306)
2012年秋冬季利文斯顿岛南极磷虾渔业CPUE指标变动及其影响因素
朱国平1、2、3,朱小艳2,孟涛2,赵勇4,徐怡瑛2,吴继魁4,许柳雄1、2、3
(1.上海海洋大学海洋科学学院,上海201306;2.国家远洋渔业工程技术研究中心,上海210306;3.大洋渔业资源可持续开发省部共建教育部重点实验室,上海201306;4.上海海洋大学食品学院,上海201306)
基于南极磷虾渔业科学观察员收集的南极磷虾渔业数据,对2012年秋冬季利文斯顿岛南极磷虾渔业单位捕捞努力量产量 (CPUE)的两个指标[单位作业小时产量(CH)和单位扫海面积产量(CPUA)]的变化及其影响因素进行了研究。结果表明:该水域南极磷虾渔业CH和CPUA分别为0~84.0 t/h和0~15.0×103t/km2,平均值分别为10.0 t/h和4.7×103t/km2;作业水域主要集中在利文斯顿岛西部水域;南极磷虾丰度较高的时间在8:00~16:00,而其他时间段则并非该渔业的最佳作业时段;46.2%的作业区域表温为-0.4~-0.2℃,43.4%的作业拖曳深度为40~60 m,78.3%的作业区域均在100 m以浅的水域。
南极磷虾;利文斯顿岛;单位捕捞努力量产量;丰度
南极磷虾为栖息于南极水域的所有磷虾种类的统称,通常所称的南极磷虾为南极大磷虾Euphausia superba Dana,属于甲壳类浮游动物[1],是南大洋的关键物种,既为该海域浮游植物主要的消费者,也是更高营养水平生物最为普遍的摄食对象[2]。南极磷虾具有巨大的渔业潜力[3-4],可用于食用、制药和水产养殖饲料原料等方面[5-6]。南极磷虾资源的开发始于20世纪60年代初,前苏联首先进行试捕,在政府补贴支持下,70年代初逐步形成规模,并于80年代初达到最高峰,年渔获量超过50万t。近年来,随着科技的发展,高效捕捞技术得到较大的改善,磷虾渔业作业范围也随之进一步拓展。中国于2009年首次进行开发性南极磷虾的资源调查,产量近2 000 t,至2011年,中国南极磷虾产量已达1.8万t。
通常情况下,商业性渔业数据主要是一些表达单位捕捞努力量产量 (CPUE)的渔业指标,如单位捕捞天数产量 (catch per fishing day,CFD)、单位作业小时产量 (catch per hour,CH)、单位网次产量 (catch per haul,CPH)、单位扫海面积产量(catch per unit sweeping area,CPUA)及单位拖曳水体产量 (catch per unit volume,CPUV)等。这些指标可用于分析南极磷虾资源丰度的相对变动[7],但这些指标受季节、海区、渔船作业模式及产品形成等因素的影响,具有较大的波动性,给准确分析南极磷虾资源状况带来了较大的困难[8]。目前,上述一些渔业指标仍为养护南极海洋生物资源委员会 (CCAMLR)评估南极磷虾渔业的主要手段[9]。不同的学者使用的渔业指标不尽相同[3,7-8,10-11],至今仍存在一定的争议[12]。但环境因素与南极磷虾资源变动之间的关系一直以来都是CCAMLR生态系统监视工作组 (WG-EMM)关注的议题[10]。一些学者也尝试利用渔业指数分析环境因素对南极磷虾资源所产生的影响[3,13-14]。作为南极磷虾渔业的主要渔场之一,南极利文斯顿岛水域一直是南极科学考察的重点水域,也是南极磷虾资源丰度较高的水域。至目前为止,很少有学者对秋冬季节利文斯顿岛周围水域南极磷虾渔业的渔业指标及其影响因素进行过分析。为此,本研究中利用 2011—2012年渔季中国南极磷虾渔业科学观察员所获取的数据,对南极磷虾渔业指标及其影响因素 (表温、水深和拖曳深度等)进行分析,旨在为进一步规模性开发南极磷虾资源提供基础数据,并为海上生产提供指导性参考。
1 材料与方法
1.1 材料
所有样本均取自于2012年5月25日—6月12日在南极利文斯顿岛水域 (59°06'~61°52'W, 62°31'~63°41'S)(图1)。
调查船为中国大型拖网渔船 “开利”轮。取样工具为商业性中层磷虾拖网,系四片式单船中层拖网,网具参数分别为上纲113 m,下纲113 m,网囊网目尺寸为2.0 cm。
1.2 方法
本次调查为渔业调查,故未采取走航式调查,仅为随机调查。作业时,按照CCAMLR要求,记录起放网的时间和位置、开始和结束捕捞的时间和位置、拖曳深度、拖速、起放网时的表温 (SST)和海底深度等参数[3]。作业网次共106网。
本研究中采用两个渔业指标CH和CPUA显示利文斯顿岛水域南极磷虾渔业变化及其影响因素,其中CH数据处理方法同朱国平等[3]的方法,但作业时间为小时,故单位为t/h;CPUA采用下述公式估算:

式中:CPUA为单位作业面积产量 (t/km2);C为每个网次的磷虾产量 (t/net);T为每个网次的捕捞时间 (h/net),即开始捕捞至结束捕捞所需的时间,一般为0.5~2 h;v为船速 (kn),通常为2.0~3.0 kn;M为网口宽度 (m)。将CH划分成5个等级,分别为 [0~5)、[5~10)、[10~15)、[15~20)、≥20 t/h;将CPUA以2×103t/km2划分成8个等级,分析两个渔业指标的分布情况。
时段:以2 h为间隔划分1天中的24 h,自0时起共计12个时段,分析不同时段内南极磷虾渔业指标的变化情况。
表温:由于捕捞期间的表温范围为-1.8~0.4℃,故以0.2℃为间隔将表温划分成12个区间,据此分析表温对渔业指标的影响。
水深:捕捞水域靠近利文斯顿岛,海底深度多在100 m以浅,但有些水域水深超过了800 m,为此将海底深度以100 m为间隔划分,共有8个区间,据此分析不同海底深度 (不同水深)南极磷虾渔业指标的变化。
拖曳深度:拖曳深度依南极磷虾分布和集群水层而变化,也会因海区而有所不同。为了分析拖曳深度与渔业指标之间的关系,本研究中以20 m为间隔将拖曳深度划分成6个等级,分别为≤20、(20~40)、 [40~60)、 [60~80)、 [80~100)、≥100 m。
以上指标若为平均值,则利用Bootstrap法估算该平均值的95%置信区间,每组数据进行重抽样 (resampling),并迭代1 000次,以降低观察次数不足带来的不确定性。利用配对样本t检验分析CPUE指标 (CH和CPUA)是否存在显著性差异(显著性水平设为0.05),以变异系数 (CV)表示各因素不同区间CH和CPUA的变化情况。利用多样本Kruskal Wallis(K-W)检验法分析各影响因素不同范围内CH和CPUA是否存在差异 (显著性水平设为0.05)。
2 结果
2.1 总体情况
配对样本t检验分析显示,CH和CPUA之间存在显著性差异 (t=5.906,df=105,P<0.001),表明在表达CPUE时两个渔业指标无法统一,因此,分别就两个渔业指标的变化及其影响因素进行分析。
2012年秋冬季利文斯顿岛南极磷虾的CH和CPUA分别为0~84.0 t/h和0~15.0×103t/km2,平均值分别为10.0 t/h(95%置信区间为8.10~12.28 t/h)和4.7×103t/km2[95%置信区间为(4.18~5.37)×103t/km2]。67.0%的CH值处于10 t/h以下,且 (5~10]t/h所占的比例最高(占36.8%),其次为 (0~5]t/h(占30.2%),而 88.7%的 CPUA值处于 8×103t/km2以下, (2~4)×103t/km2和 [4~6)×103t/km2所占的比例相近 (均为30%左右),且明显高于其他范围。
作业水域主要集中在利文斯顿岛西部水域(图2)。两个渔业指标显示的结果存在较大的差异,CH最大值 (84.0 t/h)靠近南极半岛,而CPUA最大值 (15.0×103t/km2)则在利文斯顿岛西部水域。CH值在20 t/h以上这个区间内,各值之间的差异最大 (CV=0.536),而CPUA在 (0~2) ×103t/km2时,各值间的差异最大 (CV= 0.569)。

图1 作业位置Fig.1 Fishing locations
2.2 时段
从整体上来看,平均CH和平均CPUA随时间的推移呈现相似的趋势,即8:00之前,两个渔业指标均为平稳,分别处于6 t/h和2×103t/km2左右,随后尽管趋势稍有不同,但基本上均开始增长,分别在12:00~14:00时和14:00~16:00时达到最大值后开始下降 (图3)。由此可知,南极磷虾适宜作业时间应该在8:00~16:00。K-W检验结果表明,不同时段 CPUA值差异不显著(χ2=20.413,df=11,P=0.040>0.05),而CH值则呈显著性差异 (χ2=31.102,df=11,P=0.001<0.05)。
2.3 表温
46.2%的作业时间时内,表温为-0.4~-0.2℃。除了-1.2~-1.0℃外,其他表温范围内,CH和CPUA的趋势较为一致 (图4)。当表温为-1.2~-1.0℃时,CH达到最低 (4.2 t/h),而CPUA值则为最高 (7.7×103t/km2),造成这个差距的原因可能是样本量较少 (n=1)。K-W检验结果表明,不同表温段CPUA值 (χ2=12.475,df= 6,P=0.052>0.05)和CH值 (χ2=5.268,df=6, P=0.510>0.05)差异均不显著。
2.4 海底深度

图2 利文斯顿岛水域南极磷虾CH和CPUA的空间分布Fig.2 The spatial distribution of fishing indicators(CH and CPUA)for Antarctic krill fishery
78.3%的作业区域均在100 m以浅的水域。当海底深度在400 m以浅水域时,CPUA均处于5.0× 103t/km2以下,并随着海底深度的增加而增加;当海底深度为500~600m时,CPUA达到最高,为9.1×103t/km2。而CH则波动较大 (图5)。KW检验结果表明,不同海底深度段CPUA值 (χ2= 6.367,df=5,P=0.272>0.05)和CH值 (χ2= 4.568,df=5,P=0.471>0.05)差异均不显著。
2.5 拖曳深度

图3 利文斯顿岛水域南极磷虾渔业指标随时段的变化Fig.3 The variations in the fishery indicators for Antarctic krill fishery in Livingston Island with time

图4 利文斯顿岛水域南极磷虾渔业指标随表温的变化Fig.4 The variations in the fishery indicators for Antarctic krill fishery in Livingston Island with SST

图5 利文斯顿岛水域南极磷虾渔业指标随海底深度的变化Fig.5 The variations in the fishery indicators for Antarctic krill fishery in Livingston Island with sea depth
43.4%的作业拖曳深度在40~60 m。整体上来看,不同拖曳深度范围内,CPUA值较为稳定,基本上维持在 (4~5)×103t/km2,而CH值则不同 (图6),其原因可能也是样本不足造成的。当拖曳深度大于20 m时,CH和CPUA值的变化趋势一致,均在80~100 m时达到最大值,分别为19.2 t/h和7.2×103t/km2。K-W检验结果表明,不同拖曳深度段CPUA值 (χ2=3.180,df=5,P= 0.672>0.05)和CH值 (χ2=4.891,df=5,P= 0.429>0.05)差异均不显著。
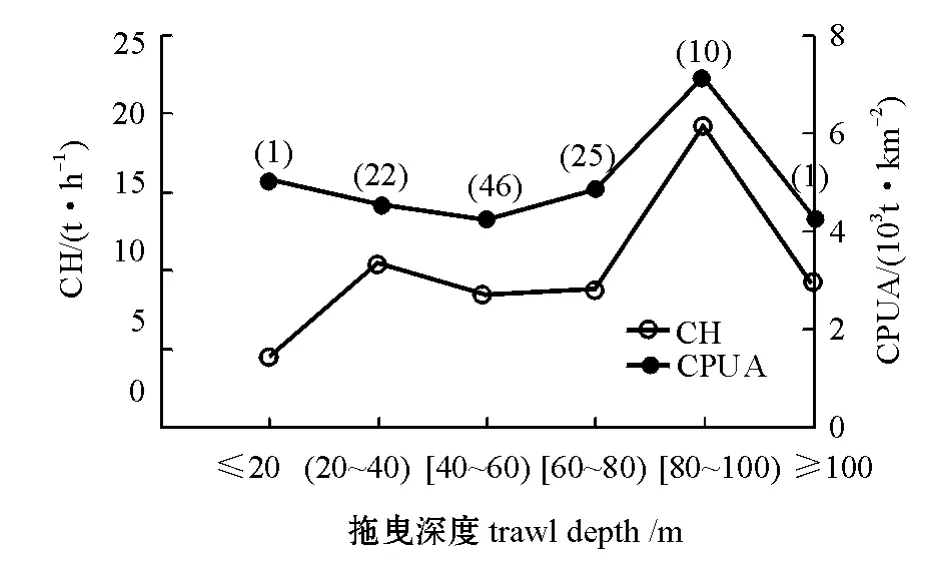
图6 利文斯顿岛水域南极磷虾渔业指标随拖曳深度的变化 (图中括号内为样本量)Fig.6 The variations in the fishery indicators for Antarctic krill fishery(the number in the bracket is sample size)in Livingston Island with trawling sea depth
3 讨论
3.1 渔业指标及其与资源丰度间的关系
许多研究人员对南极磷虾渔业CPUE指标进行过分析[11,15-16],这些指标包括CFD、CH、CPH和单位延长捕捞天数产量 (catch per extended fishing day,CEFD)等,但各指标间的统一仍存在一定的问题,上述指标所使用的初始数据为每个网次的产量 (简称网次产量)和每个网次的所用时间 (简称网次时间)。虽然CFD、CEFD、CFD与CH月平均值之间存在着较强的正相关,但由于渔业策略(人类消费、虾肉、冻虾等)所引起的网次持续时间的波动较大,造成这些指数的日均值之间的关系却非常弱。若针对不同的策略单独考虑,则网次时间、日均 CFD和日均 CH具有较强的正相关[7]。前苏联在20世纪70年代末期至1991年这段时间内,单日网次为1~15网,每日2~4网较为常见,网次时间为0.1~16 h,多数在3~8 h。若最终产品的要求较高,则网次时间较短,单日网次数会较高。朱国平等[3]认为,2009年夏季中国南极磷虾渔业作业网次也基本上维持在10网以下,其中单日2~4网居多,网次时间多为 1.5 h以下(77.3%),最长拖曳时间不足3 h。本研究中,虽然最终产品多为冻虾或虾粉 (产品质量要求不高),但单日网次也基本上处于10网以下,每日7~9网的作业天数占总作业天数的52.6%,网次时间为0.5~2.5 h的占总网次时间的76.4%,最长拖曳时间不足5 h,网次时间与每个网次的CH (简称网次CH)之间呈负相关 (Pearson相关检验,r=-0.374),日均网次时间与网次CH之间也呈负相关 (r=-0.123),而网次时间与每个网次的CPUA(简称网次CPUA)之间却呈较弱的正相关(r=0.148),日均网次时间与网次CPUA之间则呈较强的正相关 (r=0.491),导致这个结果的原因之一可能是渔业策略 (冻虾及虾粉需求)不同。
将南极磷虾渔业指标 (如CPUE)与资源丰度之间建立联系的一个关键要求是考虑不同空间尺度上磷虾所体现的集群行为。磷虾是一种专性集群的海洋甲壳类生物,依形成方式及空间形态可将这些集群划分成多种类型[17],而这些集群类型也对应着不同的磷虾丰度,从表面上看,CH与密度之间呈现合理的线性关系,但若捕捞能力波动引起饱和效应,则会导致两者之间的关系呈非线性[18]。早期磷虾渔业在很大程度上类似于捕捞分散鱼类的底拖网渔业,而捕捞单个集群的作业则与标准作业有着较大的差别,日本拖网渔船并非以具体的集群为目标,此时的CH不仅仅是表示集群中的磷虾密度,而且是表示整个研究区域内所有集群的平均密度,因此可认为,CH可作为较大范围磷虾密度的一个良好指标。Butterworth等[19]曾提议,CH应该作为较大范围磷虾集群密度的一个指标,而不能作为单个磷虾集群的密度指标,但其又在该报告讨论中表示这个结果可能不正确。事实上,在实际渔业生产中,仅一个集群的磷虾就足以保证一个网次的磷虾产量。因此,如果将CH作为集群密度的指标,首先应该记录每网次拖曳的集群数量,如果一个网次捕捞几个集群,则应该在计算时考虑到这个情况。如在渔期开始时,由于集群较小,这种情况可能有所不同,一个网次可能会捕捞多个集群。在捕捞过程中,当一个集群捕捞完成后,渔探仪未发现下一个集群,这时可能会有两种后续操作,一个是仍然将网具放在水中拖曳,而另一种则是起网待寻找到其他的集群时再下网 (即增加了网次数量)。针对后一种情形,以CH解释该海区磷虾密度便显得更为复杂。
限于渔船的加工能力不同,将单日捕捞产量作为磷虾丰度指标并不可取,但网次捕捞产量也显然不宜作为监视磷虾丰度趋势的可用指数。当渔船转移到一个新的渔区时,第一网的产量通常较低,因为此时所关注的是该海区磷虾的质量如何,而非一味追求高产。更为重要的是,网次产量通常最多维持在5~10 t,原因有两个:一是更多的网次产量会因磷虾遭挤压而致使产品质量下降,二是作业受渔船加工能力的限制。因此,从某种意义上来讲,网次产量应该是网具饱和的一个体现。
Kawaguchi等[10]采用CPUV并利用日本南极磷虾渔业数据对该渔业指标的长期波动进行了分析。分析认为,以CPUA(或CPUV)作为磷虾绝对密度可能会因以下几种原因造成过低估算的结果: 1)过低估计拖曳持续时间;2)过高估计网口拖曳面积;3)过高估计滤水率;4)大个体磷虾的规避行为。但如果上述几种情况能够较好地得到处理,则CPUA将是一种非常有用的磷虾绝对密度指标。
当在各个大规模南极磷虾集群中所耗费的找虾时间并非主要因素时,简单的渔业指标则会是预测南极磷虾丰度非常有用的工具[15],因为在南极磷虾渔场内,拖网作业时覆盖的水面要远远大于南极磷虾的集群范围,这就表明作业区域并非仅仅为传统意义上的渔场,还可直接定义为捕捞特定的南极磷虾集群[15]。总之,CH可作为渔业丰度的一个指标,但仍需要考虑一些因素,这些因素包括拖网渔船的独立性及季节性模式、作业方式和产品形式等,从而改善渔业指标数据的使用。一些研究人员也用该指标对南极磷虾渔业生产进行了分析和预测[3,10,20]。通过本研究分析可知,在样本量足够的情况下,CH和CPUV之间的整体趋势基本上保持一致,但CPUV较CH更为稳定。此外,还有一点需要注意,本研究中所使用的CPUE指标 (CH和CPUA)均未进行标准化,这会给计算结果带来一定的不确定性。由于这些指标与许多影响因素之间呈现非线性关系,故需要利用一些非线性模型,如采用广义可加模型 (GAM)对其进行标准化,从而降低用CPUE指标表征渔业资源丰度时所带来的外来干扰[21-23]。
3.2 影响南极磷虾渔业指标的因素
南极磷虾是一种喜冷水环境的浮游生物,成体栖息于200 m以浅水域,幼体则在表层水域中栖息的时间较长[24],其对海洋温度和海冰密度极为敏感[25]。磷虾栖息地分布范围的极限温差约为7℃, 1~2℃的水温变化对南极磷虾生理功能、分布及行为会产生显著的影响。因此,水温是影响南极磷虾分布的一个重要因素。Priddle等[13]曾详细地讨论了影响南极磷虾渔业指标的相关机制,并认为捕获率变动与表温有关。Kawaguchi等[26]认为,通过科考和渔业调查获取的表温信息可作为预测48.3区南极磷虾产量的指标之一,夏季初渔期的表温状况可用于预测整年的磷虾捕捞情况,且这种预测的可靠性已通过比较某些年的预测产量和实际产量予以证实。朱国平等[3]量化了夏季南极磷虾渔业指标 (CH)与表温之间的关系,并得出起放网表温在0.5~1.0℃及1.0~1.5℃时平均CH均较高。本研究结果显示,46.2%的作业时间内的表温均在-0.4~-0.2℃。尽管在温度范围上与之前的结果有明显差异,但考虑本研究中的调查时间为秋冬季节,而之前的研究多在夏季,同时考虑到本研究中的主要作业水温范围 (-0.4~-0.2℃)相对来说是该季节偏高 (但不是最高)的水温段,这点与之前的其他研究结果保持一致。
日本渔船曾于1980年初期通过底拖网调查试验 (拖曳水层依200 m划分)得知,瞄准捕捞南极磷虾集群的拖网渔业,其捕捞水层通常从表层至海面以下几百米,有时甚至达到800 m[23]。朱国平等[3]认为,夏季南极磷虾渔业指标 (CH)随着拖曳水深的增加而减小,当拖曳水层在25 m以浅时CH最大。Taki等[14]认为,冬季商业性磷虾拖网的拖曳水层通常较深,且不同渔场的拖曳深度范围也各不一致 (南设得兰群岛水域为90~110 m,南奥克尼群岛水域为160~240 m,南乔治亚群岛水域为120~160 m),而夏季的拖曳深度则在40~70 m。冬季南极磷虾渔业声学映像显示,一些密度较高的集群出现在商业性拖网瞄准捕捞的近底层(170~250 m)[26],但一些集群也出现在近表层。之所以存在差异,一个原因可能是各研究的调查时间不同,瞄准捕捞的集群结构和密度也可能影响到拖曳深度,且拖曳时段 (白天或夜间)也会对拖曳水深产生一定的影响。通过比较可知,本研究中的平均拖曳深度 (55 m,n=106)较朱国平等[3]的平均拖曳深度 (52 m,n=107)稍深,且优势拖曳深度分布范围也较后者高,这就佐证了Kawaguchi等[26]的研究结果。还应该关注的是,白天拖曳水层与南极磷虾个体有着明显的关系,因为大个体磷虾白天会下潜到更深的水层。
致谢:感谢上海开创远洋渔业有限公司及“开利”轮船长和船员在取样过程中给予的大力配合!感谢中国南极海洋生物资源开发利用项目的支持及中国南极磷虾渔业科学观察员在海上数据收集过程中所付出的辛勤劳动!
[1] Siegel V.A concept of seasonal variation of krill Euphausia superba distribution and abundance in west of the Antarctic Peninsula [C]//Sahrhage D.Antarctic Ocean and Resources Variability. Berlin:Springer Verlag,1988:219-230.
[2] Nicol S,Endo Y.Krill fisheries:development,management and ecosystem implications[J].Aquat Living Resour,1999,12:105-120.
[3] 朱国平,冯春雷,吴强,等.南极磷虾调查CPUE指数变动的影响因素初步分析[J].海洋渔业,2010,32(4):368-373.
[4] 朱国平.南极磷虾种群生物学研究进展Ⅰ——年龄、生长与死亡[J].水生生物学报,2011,35(5):862-868.
[5] Everson I.Krill:Biology,Ecology and Fisheries[M].Oxford: Blackwell Science,2000.
[6] 孙松,严小军.南极大磷虾的生物活性物质及其用途研究进展[J].极地研究,2001,13(3):213-216.
[7] Litvinov F F,Sundakov A Z,Arkhipov V I.CPUE indices used in Soviet krill fishery statistics from 1977 to 1992 and their possible utility for evaluation of relative changes in biomass[J].CCAMLR Sci,2004,11:165-177.
[8] 陈雪忠,徐兆礼,黄洪亮.南极磷虾资源利用现状与中国的开发策略分析[J].中国水产科学,2009,16(3):451-458.
[9] CCAMLR.Report of the working group on ecosystem monitoring and management[R].Busan:CCAMLR,2011:92.
[10] Kawaguchi S,Ichii T,Naganobu M.Catch per unit effort and proportional recruitment indices from Japanese krill fishery data in subarea 48.1[J].CCAMLR Sci,1997,4:47-63.
[11] Sushin V A.Distribution of the Soviet krill fishing fleet in the South Orkneys area(Subarea 48.2)during 1989/90[J].CCAMLR Sci,1998,5:51-62.
[12] SC-CAMLR.WG-FSA Standard Assessment Methods[R].Hobart:CCAMLR,2002.
[13] Priddle J,Croxall J P,Everson I,et al.Large-scale fluctuations in distribution and abundance of krill—a discussion of possible causes[C]//Sahrhage D.Antarctic Ocean and Resources Variability. Berlin:Springer-Verlag,1988:169-182.
[14] Taki K,Hayashi T,Naganobu M.Characteristics of seasonal variation in diurnal vertical migration and aggregation of Antarctic krill Euphausia superba in the Scotia Sea,using Japanese fishery data [J].CCAMLR Sci,2005,12:163-172.
[15] Fedoulov P P,Murphy E,Shulgovsky K E.Environment-krill relations in the South Georgia marine ecosystem[J].CCAMLR Sci, 1996,3:13-30.
[16] Siegel V,Damm U,Sushin V A.Catch per unit effort(CPUE) data from the early years of commercial krill fishing operations in the Atlantic Sector of the Antarctic[J].CCAMLR Sci,1998,5: 31-50.
[17] Kalinowski J,Witek Z.Scheme for classifying Antarctic krill[J].BIOMASS Handbook Series,1985,27:1-12.
[18] Cooke J G.On the relationship between catch per unit effort and whale abundance[J].Rep Int Whal Commn,1985,35:511-519.
[19] Butterworth D S,Miller D G M.A note on relating Antarctic krill catch-per-unit-effort measures to abundance trends[J].S Afr J Antarct Res,1985,17(2):112-116.
[20] Everson I.Can we satisfactorily estimate variation in krill abundance?[C]//Sahrhage D.Antarctic Ocean and Resources Variability.Berlin:Springer Verlag,1988:199-208.
[21] 朱国平.基于广义可加模型研究时间和环境因子对南极半岛北部南极磷虾渔场的影响[J].水产学报,2012,36(10):162-171.
[22] 朱国平,朱小艳,徐怡瑛,等.基于GAM模型分析夏秋季南奥克尼群岛南极磷虾渔场时空分布及与环境因子之间的关系[J].极地研究,2012,24(3):62-69.
[23] Santora J A,Sydeman W J,Schroeder I D,et al.Krill space:a comparative assessment of mesoscale structuring in polar and temperate marine ecosystems[J].ICES J Mar Sci,2012,69(7): 1317-1327.
[24] Ban S,Ohi N,Shee S,et al.Effect of solar ultraviolet radiation on survival of krill larvae and copepods in Antarctic Ocean[J].Polar Biol,2007,30:1295-1302.
[25] Huang T,Sun L,Stark J,et al.Relative changes in krill abundance inferred from Antarctic fur seal[J].Plos One,2011,6(11): e27331.
[26] Kawaguchi S,Nicol S.Learning about Antarctic krill from the fishery[J].Antarctic Sci,2007,19(2):219-230.
The variation and impacting factors in CPUE index of Antarctic krill fishery around Livingston Island during fall-winter of 2012
ZHU Guo-ping1,2,3,ZHU Xiao-yan2,MENG Tao2,ZHAO Yong4, XU Yi-ying2,WU Ji-kui4,XU Liu-xiong1,2,3
(1.College of Marine Sciences,Shanghai Ocean University,Shanghai 201306,China;2.National Engineering Research Center for Oceanic Fisheries, Shanghai 201306,China;3.Key Laboratory of Sustainable Exploitation of Oceanic Fisheries Resources,Ministry of Education,Shanghai 201306, China;4.College of Food Science and Technology,Shanghai Ocean University,Shanghai 201306,China)
The catch per unit effort(CPUE)including catch per hour(CH)and catch per unit sweeping area (CPUA)and their impacting factors were evaluated in Antarctic krill Euphausia superba fishery in Livingston Island during fall-winter of 2012 based on the catch data collected by the Antarctic krill fishery observers.The results showed that the two fishing indicators were found to range from 0 to 84.0 t/h in CH and from 0 to 15.0×103t/km2in CPUA,with average CH of 10.0 t/h and average CPUA of 4.7×103t/km2.The main fishing ground was found in the western Livingston Island,and the higher abundance of Antarctic krill was observed from 8:00 to 16:00, while the other time was not the optimal fishing time.About half of fishing operation(46.2%)was found at sea surface temperature from-0.4 to-0.2℃ and the trawling(43.4%)was carried out at depth from 40 m to 60 m, the predominant fishing ground in the shallow area at depth of below 100 m.
Antarctic krill;Livingston Island;catch per unit effort(CPUE);abundance
S931.41;S931.42
:A
2095-1388(2013)04-0399-07
2012-10-25
国家科技支撑计划项目 (2013BAD13B03);国家海洋局极地科学重点实验室开放基金资助项目 (KP201210);上海海洋大学“海鸥计划”项目 (B-5003-11-0023)
朱国平 (1976-),男,副教授。E-mail:gpzhu@shou.edu.cn
