杉木林分内优势木与平均木的遗传效应分析
2013-05-16游昌顺程小凯张建章肖纪军
游昌顺,程小凯,张建章,肖纪军
(1. 浙江省龙泉市林业局,浙江 龙泉 323700;2. 浙江省龙泉市林业科学研究院,浙江 龙泉 323700)
杉木林分内优势木与平均木的遗传效应分析
游昌顺1,程小凯1,张建章2,肖纪军2
(1. 浙江省龙泉市林业局,浙江 龙泉 323700;2. 浙江省龙泉市林业科学研究院,浙江 龙泉 323700)
在3个杉木产地分别随机选择当地起源的4个林分,各林分采集1株优势木种子(C1)和5株平均木(平均木之间相距50 m以上)的混合种子(C2),共计12对24份种子,进行育苗、造林对比试验。结果表明,12个林分的优势木生长速度总平均数大于平均木总平均数;10年生胸径生长量多重比较,C1 > C2达到差异显著水平;在杉木林分中按一定标准选出的林分优势木和该林分的平均木之间的生长速度差异基本上是能够遗传给 F1代的;产地间的遗传方差占总遗传方差的73.9%以上,占近3/4的绝对优势,而林分间与林分内单株间的遗传方差合计只占近1/4。
杉木;林分;优势木;平均木;遗传差异
20世纪60年代以来,我国开展了杉木遗传育种工作,杉木种子园建设也从1代(初级)发展到高世代,目前已在生产上发挥了作用。本研究分别采集杉木不同产地不同林分内优势木与平均木的自由授粉种子进行成对比较,试图验证它们的遗传效应。
1 材料和方法
1.1 材料
1982年秋冬,分别在广西融水(以A1表示)、贵州锦屏(A2)和浙江临安(A3)3个产地,每个产地随机选择当地起源的4个林分,林分年龄≥25 a,即3个种源内各4个林分(B1、B2、B3、B4),共计12个林分,然后采集各林分1株优势木种子(C1)和5株平均木(平均木之间相距50 m以上)的混合种子(C2),共计12对24份种子,于1983年育苗,1984年春造林。
1.2 试验方法
为重点考察优势木与平均木之差异,采用三裂式裂区试验设计。种源为主区处理(A),林分为裂区处理(B),优势木与平均木为小裂区处理(C),即优势木与平均木放在相邻小区。首先主区处理均随机排列,然后裂区处理均随机排列,最后小裂区处理均随机排列。5株直行小区,重复10次。
1.3 造林情况
造林地点,浙江省龙泉市查田镇下堡岭脚,开旷的山间丘陵,普通红壤,土层厚度1 m以上。全垦整地,株行距2 m×2 m。当年造林成活率达99.9%。
1.4 分析方法
1988年l月和1992年12月对试验林进行生长量调查,测定数据按三裂式裂区试验设计进行统计分析,采用DPS(date processing system,数据处理系统)进行数据分析。
2 结果与分析
5年生(包括苗期1年)试验林总平均树高3.69 m,平均胸径5.0 cm;10年生(包括苗期1年)平均胸径9.68 cm,平均单株材积0.031 97 m3。林木生长均良好。
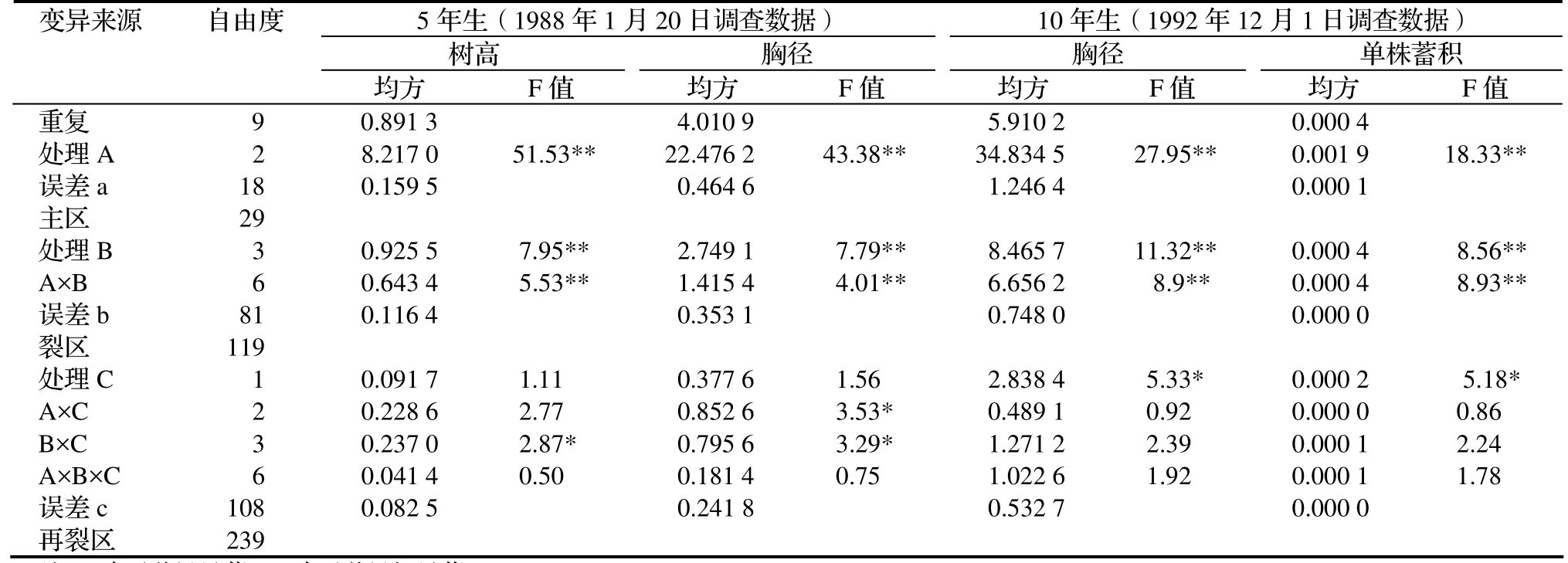
表1 杉木林分内优势木与平均木成对对比试验林方差分析Table 1 ANOVA on comparison test of dominant and mean sample tree in C. lanceolata stands
2.1 结果分析
从表1可以看出,杉木A(产地)间和B(林分)间的生长差异无论是5年生树高、胸径,还是10年生的胸径、单株蓄积都表现出极显著的差异,均达到1%的水平;而C(优势木与平均木)间的5年生树高、胸径生长差异不显著,只有10年生的胸径、单株蓄积表现出显著的差异,达到5%的水平。这可能与杉木的生长规律中“胸径生长在6 ~ 12 a最快”[1]有关,也就是说,C间也是有生长差异的,但是5年生时差异不明显,而经过第6至第10年共5 a的胸径快速生长,10年生时才生长差异明显,达到显著水平。这说明,假如把C间看成是林分内单株间,在杉木速生丰产这个遗传改良指标中,无论在产地间、产地内林分间或林分内单株间三个层次中进行选择都能取得良好的选择效果[2]。但是,选择效果是A间最好。因为其遗传方差分量(见表2)占绝对优势[2]。
2.1.1 胸径遗传方差分量年份间变动趋势 从表2还可以看出,杉木胸径生长三个层次的遗传方差分量占比会因生长时间长短不同而出现变化,A间方差分量占方差分量的百分比从 5年生时的 86.61%降到 10年生时的73.86%,降幅14.7%;B间从5年生时的13.04%升到10年生时的22.82%,升幅75%;C间从5年生时的0.35%升到10年生时的3.32%,升幅841%。也就是说,胸径遗传方差分量年份间变动趋势是:从5年生到10年生,A间方差分量占方差分量的百分比呈下降趋势;B间、C间呈上升趋势,C间的升幅最大。
2.1.2 ABC之间的互作效应分析 5年生树高和胸径,以及10年生的胸径、材积的生长差异中,产地与林分(A ×B)的互作效应表现出差异都极显著,均达到1%的水平;说明林分间的生长差异是因产地不同而不同;同时产地本身的差别会因林分不同而受到影响。
4项生长指标的(A×B×C)的互作效应差异均不显著。

表2 杉木种内遗传变异的相对数量组成Table 2 Intra-specific genetic variation of tested C. lanceolata
5年生树高生长差异中,产地与林分内优势木、平均木(A×C)的互作效应差异不显著;而5年生胸径的(A×C)差异显著,说明林分内优势木与平均木间的生长差异是因产地不同而不同;同时产地本身的差别会受林分内优势木与平均木不同的影响。但是,10年生时的胸径和材积的互作效应(A×C)差异不显著。
5年生树高和胸径生长差异中,林分间与林分内优势木、平均木的互作效应(B×C)都表现为差异显著,说明林分内优势木与平均木间的生长差异也因林分不同而不同;同时林分本身的差别会受林分内优势木与平均木不同的影响。但是,10年生时的胸径和材积的互作效应(B×C)差异不显著。
2.2 处理间生长量多重比较结果
2.2.1 A处理间多重比较 从表3可以看出,5年生树高和胸径,以及10年生胸径3个调查项目生长量都是A1 > A2 > A3。经多重比较,A1 > A2、A3和A2 > A3的生长差异都达到了1%极显著水平。但是,10年生单株蓄积(根据“龙泉市立木一元材积表”把每木胸径换算成单株蓄积)略有差别,虽然A1 > A2 > A3的趋势不变,经多重比较,A1 > A2、A3的生长差异仍然达到了1%极显著水平;A2>A3却只达到了5%的显著水平。

表3 A处理间生长量多重比较结果Table 3 Multiple comparisons on increment of treatment A
2.2.2 C处理间多重比较 从表4可以看出,5年生树高和胸径的生长量都是C1 >C2,但经多重比较,它们的生长差异没有达到显著水平;10年生胸径和单株蓄积的生长量也是C1 > C2,经多重比较,C1 > C2的胸径生长差异达到了5%的显著水平,而单株蓄积差异达到了1%极显著水平。

表4 C处理间生长量多重比较结果Table 4 Multiple comparisons on increment of treatment C
2.2.2 同产地同林分的C处理间胸径生长多重比较 从表5可以看出,同产地同林分的C处理间共有12对,经多重比较,胸径生长中C1 > C2的共有9对。其中,5对(A1B2、A2B2、A2B3、A3B3、A3B4)表现为差异极显著,2对(A1B4、A2B1)表现为差异显著,2对(A2B4、A3B2)表现为差异不显著。胸径生长中C1 < C2的共有3对(A1B1、A1B3、A3B1)。

表5 各产地各林分的优势木与平均木胸径生长量差异Table 5 Variation of DBH growth of dominant and mean sample trees from different provenances cm
从表5还可以看出,同产地同林分的C1(优势木)10年生胸径平均生长量为9.79 cm,> C2(平均木9.57 cm)0.22 cm,达到差异显著水平。其中:产地A1平均差异不显著;产地A2平均差异极显著;产地A3平均差异也不显著。
3 结论和讨论
上述结果表明,在杉木林分中按一定标准选出的林分优势木和该林分的平均木之间的生长速度差异基本上是能够遗传给F1代的。本试验林属于12个林分优势木与平均木的F1代,调查分析结果是12个林分的优势木生长速度总平均数大于平均木总平均数。以10年生胸径为例,经多重比较,C1 > C2达到差异显著水平。其中:有9个林分为C1 > C2,占林分总数的3/4;另有3个林分为C2 > C1;占林分总数的1/4。这就说明,林分优势木的选择标准虽然低于优树的选择标准,但是选出的林分优势木生长量仍然大于林分的平均木生长量,也就是说,林分优势木比林分平均木带有更多的速生基因,以致在F1代中,表现为优势木的生长速度快于平均木。研究结果进一步验证了陈益泰提出的杉木“在未经改良的人工林中,按照一定标准选出的优树,一般带有较多的速生基因。虽然直接在林分中从优树上采种造林,也有一定的遗传增益”的结论[3]。
上述结果还表明,以胸径生长为例,林分优势木和该林分的平均木之间的生长速度差异受产地(A×C)与林分(B×C)的互作效应影响是:5年生时差异显著,10年生时差异不显著。也就是说,杉木单株间的生长速度差异受产地与林分互作效应影响是存在的,但是,这种影响会随着林龄的增长而减少。进一步验证了陈岳武提出的“有关杉木早期选择年限的结论(优良家系或个体在造林后6 ~ 7 a作第2次决选),引证到杉木产地子代测定试验是不恰当的”的结论[4]。
上述结果再表明,杉木生长的3个层次遗传方差分量中,5年生树高、胸径与10年生胸径、单株蓄积其产地间遗传方差占总遗传方差的 87.8%、86.6%、73.9%和 76.3%;产地内林分间与林分内单株间的遗传方差相加占总遗传方差的12.2%、13.4%、26.1%和23.7%。也就是说,产地间的遗传方差占总遗传方差的73.9%以上,占近3/4的绝对优势;而林分间与林分内单株间的遗传方差合计只占近1/4[2]。进一步验证了赖特等研究欧洲赤松(Piuns silvestris)得出的“在生长速度等性状上,总遗传变异的70% ~ 85%是变种间(指地理变种)的差异,15% ~ 30%则很多是属于同一变种不同林分内的差异”结论[5]。
[1] 俞新妥. 杉木[M]. 福州:福建科学技术出版社,1982:37-58
[2] 张建章,陈益泰,祝春敏,等. 杉木生长的遗传变异研究:三水平后代试验[J]. 浙江林业科技,1997,17(4):1-5.
[3] 陈益泰,张建章,游昌顺,等. 杉木初级种子园遗传效应的一个验证[J]. 林业科学研究,1990,3(2):151-153.
[4] 陈岳武,施季森. 杉木遗传改良中的若干问题(续)[J]. 南京林学院学报,1984(1):10-11.
[5](美)乔纳森 W 赖特(著),郭锡昌,胡承海(译). 森林遗传学[M]. 北京:中国林业出版社,1981. 228-230.
Genetic Analysis on Dominant and Mean Sample Tree in Cunninghamia lanceolata Stands
YOU Chang-shun1,CHENG Xiao-kai1,ZHANG Jian-zhang2,XIAO Ji-jun2
(1. Longquan Forestry Bureau of Zhejiang, Longquan 323700, China; 2. Longquan Forestry Institute of Zhejiang, Longquan 323700, China)
Four Cunninghamia lanceolata stands were randomly selected at producing areas of Guangxi, Guizhou and Zhejiang province. Seeds were collected from 1 dominant and 5 mean sample trees of each selected stands. Comparison tests were conducted on seedling cultivation and afforestation of collected seeds. The result demonstrated that mean growth rate of dominant trees was higher than that of mean sample trees of 12 tested stands. Multi-comparisons on DBH growth of 10-year trees resulted evident difference between seeds of dominant and mean sample tree. Growth rate of dominant and mean sample tree could be hereditary to their F1. Genetic variance among provenance occupied 73.9% of the total one, the left was variance among stands and with stand.
Cunninghamia lanceolata; stands; dominant tree; mean sample tree; genetic variance
S722.3
A
1001-3776(2013)06-0037-04
2013-07-11;
2013-09-27
游昌顺(1959-),男,浙江龙泉人,工程师,从事森林生态建设和林木遗传育种工作。
