深点食螨瓢虫对土耳其斯坦叶螨和截形叶螨捕食作用的比较
2013-04-26杨帅贺亚峰何欢李婷卫秋阳冯盼张建萍
杨帅,贺亚峰,何欢,李婷,卫秋阳,冯盼,张建萍
(石河子大学农学院,石河子 832003)
土耳其斯坦叶螨Tetranychus turkestani(Ugarov et Nikolski)和截形叶螨Tetranychus truncatus Ehara是新疆重要的害螨种类,危害棉花、玉米、果树、蔬菜等多种作物,对新疆的农业生产造成了严重的损失[1-3]。由于2种叶螨早期危害具有隐蔽性,所以田间一旦暴发,药剂防治效果不理想,人们就随意加大施药量,结果造成害螨抗性发展迅速[4],化学防治的效果进一步降低。因此,开展害螨的生物防治成为解决这一问题的关键,天敌昆虫的保护和利用是其中一项很重要的举措。据前人报道,2种叶螨的天敌种类非常丰富,优势种天敌有深点食螨瓢虫Stethorus punctillum Weise和塔六点蓟马Scolothrips takahashii Priesener等[5-6]。有关深点食螨瓢虫对叶螨的捕食作用的研究已有不少,如沈妙青等[7]和刘永生[8]研究了深点食螨瓢虫对柑桔全爪螨的捕食功能反应,王登元等[9]研究了深点食螨瓢虫对土耳其斯坦叶螨的捕食功能反应,Kandasamy等[10]研究了深点食螨瓢虫对茶树茶红螨的捕食功能。鲁素玲等[11]研究了深点食螨瓢虫、小花蝽和中华草蛉对土耳其斯坦叶螨的捕食功能反应。杨群芳等[12]研究了深点食螨瓢虫和塔六点蓟马对朱砂叶螨的捕食作用。
前人只研究了深点食螨瓢虫对单一叶螨的捕食作用,但对于2种叶螨捕食作用的差异性还没有相关研究。本研究以田间常见天敌深点食螨瓢虫为捕食者,研究了其对土耳其斯坦叶螨和截形叶螨的捕食作用,通过比较深点食螨瓢虫对2种叶螨的捕食效能及选择性,旨在进一步探讨天敌因子对土耳其斯坦叶螨和截形叶螨种间竞争的影响。
1 材料与方法
1.1 材料
土耳其斯坦叶螨和截形叶螨采自石河子大学农学院试验站和石河子新疆兵团第八师143团一场,在RXZ-260B型智能人工气候箱内使用刀豆Semen Canavaliae Gladiatae饲养。
深点食螨瓢虫成虫采自石河子大学农学院试验站,采回后先使其饱食,然后饥饿24h,待用。培养皿直径为9cm,中间放一直径为6cm的棉花叶碟,叶碟下依次垫有同样大小的湿滤纸和海绵。
人工气候箱设置条件为:温度(28±0.5)℃,相对湿度70%±10%,光周期16L∶8D。
1.2 方法
1.2.1 深点食螨瓢虫对土耳其斯坦叶螨和截形叶螨不同密度的功能反应
土耳其斯坦叶螨和截形叶螨选用刚羽化的且个体大小相同的雌成螨,投放量设20、30、40、50和100头5个密度,每个培养皿中接入一定数量的叶螨后,投入1头饥饿处理24h后的深点食螨瓢虫,用保鲜膜封口,并用昆虫针扎许多小孔,放置在人工气候箱中培养。另设以不接天敌且有相同密度的叶螨成螨的处理为对照,校正成螨死亡数量。每个处理重复3次,24h后观察记载各处理培养皿中剩余的和自然死亡的成螨数。
1.2.2 深点食螨瓢虫自身密度对土耳其斯坦叶螨和截形叶螨的干扰反应
2种叶螨设置密度相同,均为每皿100头,饥饿处理24h后的深点食螨瓢虫成虫按1、2、3、4头分别接入培养皿中,置人工气候箱中培养。另设以不接天敌且相同密度的叶螨成螨的处理为对照,校正成螨死亡数量。试验重复数及记载内容同1.2.1。
1.2.3 深点食螨瓢虫对土耳其斯坦叶螨和截形叶螨的捕食选择性
培养皿中先接入50头土耳其斯坦叶螨成螨和50头截形叶螨成螨,再接入1头饥饿处理24h后的深点食螨瓢虫成虫,置人工气候箱中培养。另设以不接天敌且相同密度的叶螨成螨的处理为对照,校正成螨死亡数量。试验重复数及记载内容同1.2.1。
1.2.4 数据处理
1.2.4.1 功能反应
采用 Holling-Ⅱ型圆盘方程模拟(Holling-Ⅱ,1959)[13]:Nɑ'=ɑ'NT/(1+ɑ'ThN),
寻找效应:S=ɑ'/(1+ɑ'ThN)。
其中:Nɑ'为猎物被捕食量,N 为供试猎物密度,Th为处置时间,ɑ'为捕食者对猎物的瞬间攻击率,T为试验时间(1d)。
1.2.4.2 干扰反应
采用Hassel等[14]提出的干扰反应模型模拟:
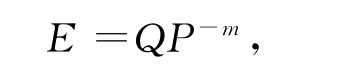
式中:E为捕食率,Q为寻找系数,P为捕食者密度,m为干扰参数。
1.2.4.3 捕食选择性
用选择系数E比较深点食螨瓢虫对两种叶螨的捕食选择性,E可用Jacobs[15]提出的方程表示为:
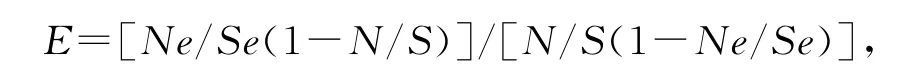
其中:E为选择系数,Ne为第1种猎物被捕食的数量,Se为2种猎物被食数量之和,N为第1种猎物的初始数量,S为2种猎物初始数量之和。当E<1,表现为负喜好,E>1,表现为正喜好。
2 结果与分析
2.1 深点食螨瓢虫对土耳其斯坦叶螨和截形叶螨不同密度的功能反应
由表1可见,同密度条件下,深点食螨瓢虫对截形叶螨的平均捕食量均大于土耳其斯坦叶螨。深点食螨瓢虫的日捕食量随叶螨密度的增加而增加,当叶螨密度达40头后,深点食螨瓢虫的捕食量增加较缓慢,即捕食量与叶螨密度为逆密度制约关系,呈负加速型曲线。因此,可用Holling圆盘方程拟合。拟合结果(表2)表明,深点食螨瓢虫成虫对2 种叶螨成螨的捕食功能反应均为Holling-Ⅱ型。

表1 深点食螨瓢虫在2种叶螨不同密度下的日捕食量Tab.1 Daily preying capacity of Stethorus punctillumon two species of mites at different density
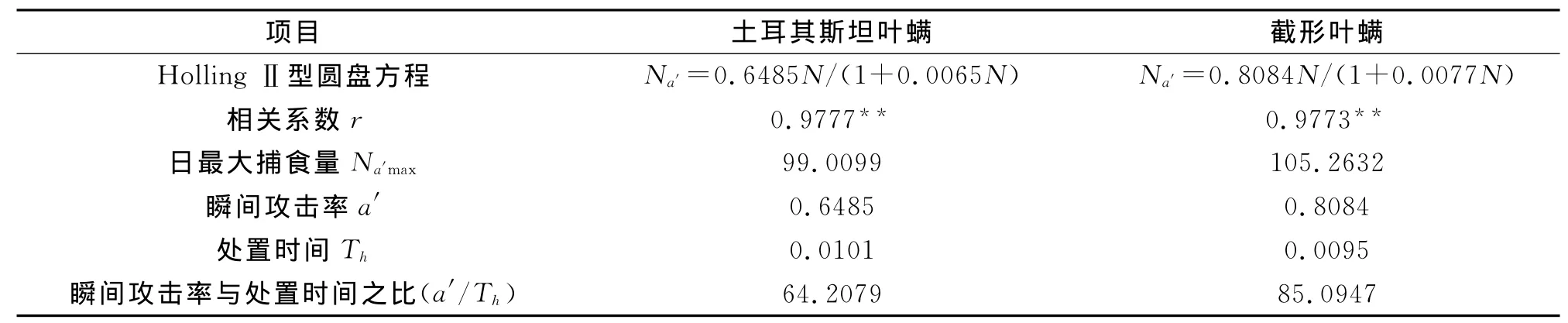
表2 深点食螨瓢虫捕食2种叶螨的圆盘方程及其参数Tab.2 Disc equation and parameters of Stethorus punctillumto two species of mites
由表2可知,用Holli-ng-Ⅱ型圆盘方程拟合效果较为理想。深点食螨瓢虫成虫对截形叶螨的日最大捕食量和瞬间攻击率(105.2632、0.8084)均大于土耳其斯坦叶螨 (99.0099、0.6485),处 置时间(0.0095)小于土耳其斯坦叶螨(0.0101)。根据瞬时攻击率(ɑ′)和处理时间(Th)之比可以衡量天敌对害虫的控制能力,ɑ′/Th越大,则表示天敌对害虫的控制能力越强。深点食螨瓢虫成虫对截形叶螨的瞬间攻击率与处置时间之比(85.0947)明显大于土耳其斯坦叶螨(66.2079),说明深点食螨瓢虫对截形叶螨的控制能力要大于土耳其斯坦叶螨。
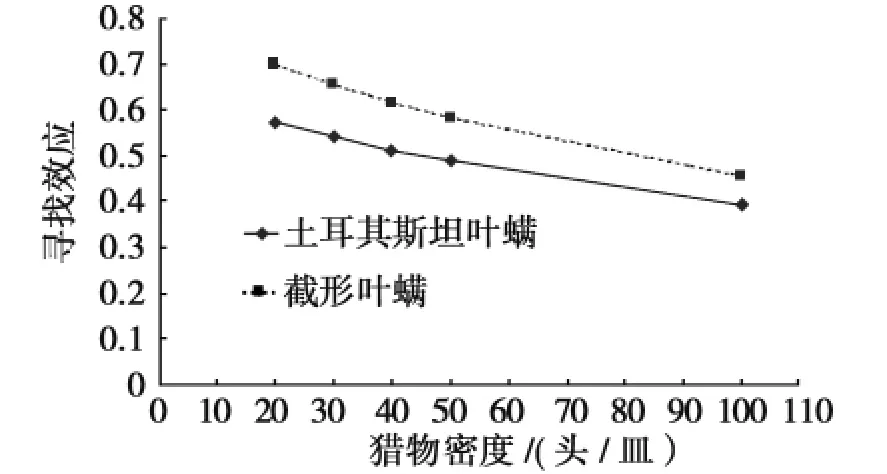
图1 深点食螨瓢虫对2种叶螨的寻找效应Fig.1 The searching efficiency of Stethorus punctillum to two species of mites
由寻找效应估计公式及表2中的各参数值,可以计算出深点食螨瓢虫成虫对土耳其斯坦叶螨和截形叶螨的寻找效应,结果见图1。
由图1可知:深点食螨瓢虫成虫对土耳其斯坦叶螨和截形叶螨的寻找效应均随着猎物密度的增加而降低;当密度相同时,深点食螨瓢虫成虫对截形叶螨的寻找效应始终高于土耳其斯坦叶螨。
2.2 深点食螨瓢虫自身密度对土耳其斯坦叶螨和截形叶螨的干扰反应
不同密度深点食螨瓢虫成虫对2种叶螨成螨的捕食率及干扰模型(表3)表明:随着深点食螨瓢虫自身密度增大,其捕食率下降。可见由于天敌密度的增大,反而增加了其个体之间的相互干扰,造成捕食率下降。相同密度下,深点食螨瓢虫成虫对截形叶螨的捕食率均大于对土耳其斯坦叶螨的捕食率。深点食螨瓢虫成虫以土耳其斯坦叶螨为食料的干扰反应模型为:

以截形叶螨为食料的干扰反应模型为:


表3 深点食螨瓢虫成虫对2种叶螨的捕食率及干扰模型Tab.3 Predation rate and interference of Stethorus punctillumto two species of mites
2.3 深点食螨瓢虫对土耳其斯坦叶螨和截形叶螨的捕食选择性
深点食螨瓢虫对2种叶螨的选择系数分别为0.7106和1.4082。其中,深点食螨瓢虫成虫对截形叶螨的选择系数大于1,表现为正喜好;对土耳其斯坦叶螨的选择系数小于1,表现为负喜好,即深点食螨瓢虫成虫偏好于取食截形叶螨。
3 讨论
本试验结果表明,深点食螨瓢虫对土耳其斯坦叶螨和截形叶螨的功能反应均符合Holli-ng-Ⅱ型,这与王登元等[9]和杨德松[16]的研究深点食螨瓢虫对土耳其斯坦叶螨的功能反应结果相同。本研究侧重研究了深点食螨瓢虫对2种叶螨捕食作用的差异,表明深点食螨瓢虫不论在何种密度下均表现为对截形叶螨的捕食量大于土耳其斯坦叶螨。深点食螨瓢虫对于2种叶螨的寻找效应均随猎物的密度的增加而降低。当密度相同时,深点食螨瓢虫成虫对截形叶螨的寻找效应始终高于土耳其斯坦叶螨。
捕食性天敌常以增加局部扩散的倾向来对同种个体的相遇行为起反应,亦为相互干扰效应。本实验的干扰作用研究证实,随着深点食螨瓢虫自身密度增加,由于种内个体间的相互干扰,深点食螨瓢虫对2种叶螨的捕食率捕食率均下降,这与王登元等[9]的研究结论相同。与功能反应的结果相同,干扰反应实验也表明同密度下深点食螨瓢虫对截形叶螨的捕食作用强于土耳其斯坦叶螨。另外,在2种叶螨共存的条件下,深点食螨瓢虫对2种叶螨有明显的选择性,深点食螨瓢虫对截形叶螨的偏好性强。
本文研究结果代表了深点食螨瓢虫成虫对2种叶螨成螨捕食能力的趋势,较为客观的评价了深点食螨瓢虫对2种叶螨的捕食能力,阐明了深点食螨瓢虫对截形叶螨的控制能力要强于土耳其斯坦叶螨。但是,本研究是在室内一个简单的封闭系统内进行的,其生存条件与田间自然环境存在较大的差异,另外田间多种猎物并存,再加上深点食螨瓢虫本身也有天敌。因此,本文试验结果可能与田间实际情况存在一定差距,在自然条件下深点食螨瓢虫对2种叶螨的捕食作用的差异、控制效果的差异等还需进一步研究。
[1]邓慧君,南更明.库尔勒香梨主要病虫害及其防治技术[J].中国果树,2001(5):37-40.
[2]鲁素玲,张建萍.新疆棉花害螨种类研究初报[J].新疆农业大学学报,1997(20):135-137.
[3]庞保平,周晓榕,史丽,等.不同寄主植物对截形叶螨生长发育及繁殖的影响[J].昆虫学报,2004,47(1):55-58.
[4]徐遥,杨秀荣,芮昌辉,等.新疆棉花主要害虫对几种杀虫剂的抗药性测定[J].西北农业学报,2004,13(2):74-78.
[5]鲁素玲,丁胜利.玛河流域棉田叶螨天敌研究初报[J].石河子农学院学报,1989,11(1):59-64.
[6]郭文超,吐尔逊,许建军,等.新疆玉米害螨天敌种类及其优势种天敌的控害效应研究[J].新疆农业科学,2003,40(2):81-83.
[7]沈妙青,郭振中,熊继文.深点食螨瓢虫对桔全爪螨的捕食作用[J].贵州农学院学报,1996,15(4):35-38.
[8]刘永生.深点食螨瓢虫生物学特性及捕食功能反应的初步研究[J].中国南方果树,2002,31(1):15-16.
[9]王登元,于江南,袁仙歌,等.深点食螨瓢虫对土耳其斯坦叶螨捕食效应的研究[A].走向21世纪的中国昆虫学[C].李典谟.北京:中国科学技术出版社,2000,1166-1169.
[10]Kandasamy P,Rajagopal S,Amsalingam R,et al.Life table and predatory efficiency of Stethorus gilvifrons(Coleoptera:Coccinellidae),an important predator of the red spider mite,Oligonychus coffeae(Acari:Tetranychidae),infesting tea[J].Experimental and Applied Acarology,2010,50:141-150.
[11]鲁素玲,刘小宁,张建萍.3种天敌对土耳其斯坦叶螨的捕食功能反应的初步研究[J].石河子大学学报:自然科学版,2001,5(3):194-196.
[12]杨群芳,卢永宏.深点食螨瓢虫和塔六点蓟马对朱砂叶螨的捕食作用[J].应用昆虫学报,2011,48(3):622-625.
[13]Holling C S.Some characteristics of simple type of predation and parasitism[J].Canadian Entomologist,1959,91:385-398.
[14]Hassell M P,Varley G C.New inductive population mode-l for insect parasite and its bearing on biological control[J].Nature,1969,223:1113-1117.
[15]Jacobs J.Quantitative measurement of food selection:a modification of the forage ratio and lvlev's selectivity index[J].O-ecologia,1974,14:413-417.
[16]杨德松.棉花-土耳其斯坦叶螨-三种天敌的关系研究[D].石河子:石河子大学,2005.
