放牧对伊犁绢蒿群落结构的影响
2013-04-25荀其蕾安沙舟孙宗玖董莉莉
荀其蕾,安沙舟,孙宗玖,董莉莉,魏 鹏
(新疆农业大学草业与环境科学学院 新疆草地资源与生态重点实验室,新疆 乌鲁木齐 830052)
伊犁绢蒿(Seriphidiumtransiliense)是隶属于菊科绢蒿属的半灌木,分布中心在我国新疆[1]。广泛分布在海拔500~1 400 m的准噶尔盆地、博乐谷地、伊犁谷地、塔城谷地的山前冲积-洪积平原、山前冲积扇和低山丘陵,是新疆天山北坡荒漠和荒漠草原的重要组成植物[2],常与草原绢蒿(S.schrenkianum)、新疆绢蒿(S.kaschgaricum)、纤细绢蒿(S.gracilescens)和博乐绢蒿(S.borotalense)等组成群落。伊犁绢蒿不仅是新疆荒漠草地中饲用价值较高的一类植物,也是荒漠区天然草地补播和固沙等生态环境建设的优良植物[2],同时还是新疆主要春秋牧场或冬春牧场的主要建群植物之一。
由伊犁绢蒿为建群种组成的蒿属荒漠草地属中亚气候型荒漠,随环境条件的变化,伊犁绢蒿在群落中的比重有所不同。由于长期的严重超载过牧,草地退化十分普遍,局部地区已经极度退化,带来植物群落的逆向演替,造成草地生产性能和质量的下降,这严重制约了新疆草地畜牧业的发展[3-5]。目前,有关伊犁绢蒿的研究,多集中于伊犁绢蒿的种质资源[1]、营养成分[2]及人工模拟放牧条件下的变化[3]等方面,对自然放牧条件下的伊犁绢蒿群落结构的研究未见详细报道。本研究对不同自然放牧压力下的伊犁绢蒿群落结构进行分析,旨在为伊犁绢蒿荒漠草地合理放牧提供科学依据,使新疆伊犁绢蒿荒漠春秋草地受损的生态系统得到恢复并可持续利用。
1 材料与方法
1.1研究区概况 研究区位于中亚荒漠天山北坡中段昌吉市西南部阿什里哈萨克民族乡的阿苇滩,地理位置87°03′-87°05′ E,43°52′-43°53′ N,海拔804~833 m,年平均气温6.4 ℃,年降水量180~190 mm,年蒸发量1 760 mm,干燥度4~10。土层深厚,为黄土覆盖的灰棕色荒漠土。研究区的草地类型为典型的伊犁绢蒿荒漠,建群种为伊犁绢蒿,伴生种有角果藜(Ceratocarpusarenarius)、木地肤(Kochiaprostrata)、叉毛蓬(Petrosimoniasibirica)、小蓬(Nanophytonerinaceum)、伊犁郁金香(Tulipailiensis)、黄芪(Astragalussp.)和四齿芥(Tetracmequadricornis)等。
1.2试验方法 依据试验区放牧点与居民区距离的远近以及群落特征,建立试验样地,且样地均位于牧户所拥有草地的中心地段。放牧家畜主要为细毛羊,放牧时间为每年的4月中下旬至6月下旬(约60 d),9月上中旬至10月中下旬(约40 d)连续放牧。于2012年5月在进行调查和实地考察的基础上,参照任继周[6]对放牧强度和草地演替阶段的划分标准,根据牲畜(新疆细毛羊)数量和草地面积及放牧时间,确定了3个放牧强度梯度,分别为轻度放牧(LG)、中度放牧(MG)和重度放牧(HG),禁牧草地作为对照(CK),选在阿苇滩某机场(表1)。

表1 不同放牧梯度群落结构特征及放牧强度Table 1 Different grazing gradient characteristic of community structure and grazing intensity
在每个放牧梯度上随机设置1 m×1 m的样方3个,记录各样方内的种类组成,分种测定地上部分高度、盖度、密度和生物量。
1.3群落多样性测定 调查样地的“四度一量”,用直接测量法测量植物的高度;直接目测法估测盖度;将样方中的植物分种剪下地上部分,称量,得到生物量(质量),并装入密封袋中带回实验室。得到了生物量、盖度、重要值等多种测度指标,根据这些数据作为α多样性指数[7-8]的测度指标。
重要值(IV)=(相对高度+相对盖度+相对密度+相对生物量)/4;
Shannon-Wiener多样性指数:
H=-∑PilnPi;
Pielou均匀度指数:
J=(-∑PilnPi)/lnS;
物种丰富度:R=S;
Simpson指数:D=1-∑Pi2;
式中,S为样地总物种数;IV为样地各物种总重要值;IVi为第i个物种重要值;Pi为IVi/S的值,即样地中第i个物种的相对重要值。
1.4数据分析 利用Excel 2003和SPSS 17.0对数据进行计算和方差分析。
2 结果与分析
2.1植物群落种类组成及结构特征 草地退化是多种因素综合作用的结果,在伊犁绢蒿荒漠草地上,过度放牧是导致草地退化的主要原因。放牧是放牧家畜对草地的一种压迫行为[9]。放牧家畜在草地上进行采食、践踏和排泄粪尿,对草地产生多种影响,如改变植物种类,改变土壤结构和肥力等。
5月,3种放牧强度下草地的植物种类数均高于未放牧的CK组,出现这种结果是因为放牧增加了群落中的空隙,为其他物种的生长创造了机会,使植物种类增多(表2)。在未放牧的伊犁绢蒿草地中,植物种数为4种,分别是伊犁绢蒿、抱茎独行菜、播娘蒿和田旋花,其中伊犁绢蒿处于绝对优势地位,重要值为0.946,其余3种植物抱茎独行菜、播娘蒿及田旋花重要值依次降低。随着放牧强度的增强,草地中的物种多样性发生了改变,有新的物种侵入,群落结构发生变化。在LG草地中,植物种数为5种,新增加了叉毛蓬、角果藜、刺沙蓬和木地肤,其重要值呈依次下降的趋势。在MG草地中,植物种数有6种,比LG草地新增加了一年生植物假狼紫草及短命植物四齿芥,其重要值表现为伊犁绢蒿(0.499)>角果藜(0.254)>木地肤(0.194)>叉毛蓬(0.032)>四齿芥(0.014)>假狼紫草(0.007)。在HG草地中,物种数上升至8种,角果毛茛的重要值为0.262,而伊犁绢蒿仅为0.218。
7月,CK组仅有伊犁绢蒿、角果藜和田旋花3种植物,重要值依次降低。在LG草地中,植物种数有6种,伊犁绢蒿重要值高于其它3个放牧梯度下的重要值,为0.716;其次是角果藜,为0.125;之后是叉毛蓬、猪毛菜、刺沙蓬和樟味藜。MG草地中,伊犁绢蒿重要值(0.349)为该月4个放牧梯度中的最小值,但仍高于MG草地中其余4种植物重要值。在HG草地,植物种数增加到7种,较MG增加了条叶车前及扁蓄,重要值表现伊犁绢蒿(0.425)>角果藜(0.253)>叉毛蓬(0.177)>樟味藜(0.099)>刺沙蓬(0.027)>条叶车前(0.011)>扁蓄(0.010)(表2)。
9月,CK区因受到人为破坏,不能作为样地代表。LG草地,植物种数为5种,重要值分别为伊犁绢蒿(0.554)>木地肤(0.228)>角果藜(0.193)>猪毛菜(0.021)>小蓬(0.005)。MG草地植物种数也为5种,伊犁绢蒿重要值依然高于其他4种植物。HG区,仅有两种植物,重要值为伊犁绢蒿(0.625)>角果藜(0.375)。

表2 伊犁绢蒿荒漠不同放牧压力下草地植物种类的组成与重要值Table 2 The floristic composition and import value of S. transiliense desert in different grazing intensity
与CK组相比,在3种放牧压力下物种数均呈现增加的趋势。从重要值看,建群种伊犁绢蒿随放牧强度的增强,其重要值呈下降趋势,而角果藜、叉毛蓬的重要值呈增加趋势。并且伊犁绢蒿由建群种退至次要地位,原属伴生或偶见种的叉毛蓬、角果藜、假狼紫草等逐步增多成为主要成分。主要原因可能是群落中的优势物种伊犁绢蒿受放牧的干扰,生长受到抑制,减少了对光资源的竞争,削弱了对群落中其他物种的抑制性,种间竞争力下降。使得竞争处于劣势的角果藜、叉毛蓬等得以快速生长,物种多样性增加。这一点符合中度干扰假说。其次是家畜选择性采食植物的直接影响[10],放牧家畜选择性采食植物,使本来作为优势种的伊犁绢蒿种群竞争力被减弱,释放了草地的竞争压力,这就有利于角果藜、叉毛蓬以及木地肤等竞争力弱但又耐牧的植物生长。
2.2植物群落物种多样性分析 群落的物种丰富度及多样性是群落的重要特征,任何一种干扰因子对群落结构影响的研究都离不开物种多样性问题[11]。利用多样性指数不仅可以反映出物种水平上群落结构、组成多样性、异质性程度[12-13],还可以表现出种间个体分布的均匀性程度以及群落物种多样性。从调查结果看(表3),本研究区草地植物群落物种多样性的分布规律因放牧强度而异。
5月,各放牧梯度的多样性指数随放牧压力的不断增强呈逐渐递增趋势。Shannon-Wiener指数在HG组最大,CK组最小(表3)。表明在HG试验区物种数最多,复杂程度最高;CK组由于伊犁绢蒿处于绝对优势地位,争取到更多的资源,而影响其他植物生长,Pielou指数物种单一。这一结果与物种丰富度变化结果相符。Pielou指数显示,CK组与LG和CK组均有显著差异(P<0.05),MG组与LG、HG组无显著差异(P>0.05)。由于物种丰富度与物种均匀度密切相关,故与Pielou指数变化相同,HG显著高于CK组(P<0.05),与LG和MG组无显著差异(P>0.05)。
7月,α多样性指数变化不同,除LG组的Pielou指数外,所有放牧草地的多样性指数均高于CK组,放牧使多样性指数增加(表3)。其中Shannon-Wiener指数表现为LG、MG与HG组无显著差异(P>0.05),CK组显著低于MG、HG组(P<0.05)。物种丰富度差异情况与Shannon-Wiener指数相同,这是因为未放牧的伊犁绢蒿草地中,群落建群种伊犁绢蒿的长势发育优良,抑制了杂类草的生长,植物多样性低。Simpson指数在不同放牧强度下差异不显著(P>0.05)。两种多样性指数之所以有差异,是因为这两种指数对于物种丰富度和均匀度的敏感性不同,Simpson指数对于均匀度的敏感性高于Shannon-Wiener指数,而Shannon-Wiener指数对于物种丰富度的敏感性高于Simpson指数。这与试验的物种丰富度与Pielou指数的结果相符。4种放牧梯度下的Pielou指数无显著差异(P>0.05),但MG组物种的均匀度最大(表3)。通常认为,具有单一优势种和寡优势种的群落均匀度较低,而具有多优势种和优势种不明显的群落均匀度较高。本研究中CK组中伊犁绢蒿处于绝对优势地位,故均匀度较低。当放牧条件出现后,伊犁绢蒿的优势中地位被其他新增物种所分配,使整个群落的优势不明显,所以MG组均匀度最大。4种处理下的物种丰富度表现为CK 表3 不同放牧梯度多样性指数Table 3 The diversity index of grasslands with grazing intensities 9月,CK组遭到人为破坏,不能作为样地代表。MG组的Shannon-Wiener指数、Simpson指数均显著大于HG组(P<0.05),表明MG组物种较丰富(表3)。这一点符合中度干扰假说[14]。LG组的Pielou指数显著小于MG组和HG组(P<0.05),MG组与HG组差异不显著。HG组的物种丰富度显著小于MG组和LG组,LG组与MG组差异不显著,表明LG组的物种较其它两个放牧强度分布均匀。 3.1群落种类组分发生变化 据研究,放牧后天然草地植物对光的竞争力变化较大[12],直立且高大类型的植物较低矮植物在获取更多光资源或光能时有较多优势[14-17];草地物种的丰富度和多度随植被高度的增加受限[18],这可能是光照的有效利用降低导致的[19]。放牧对物种丰富度的不同影响可以结合干扰水平和放牧时间长短加以解释[20],放牧使多年生和匍匐生长的植物频度增加,冠层高度降低,杂类草(包括毒草,害草)和豆科植物的频度增加,禾草频度减小[21]。本研究中4个放牧强度组,放牧使群落中优势种的重要值显著减小,这是因为家畜对伊犁绢蒿种群优先采食,使得整个群落的郁闭度降低,光照不再是其他物种的限制因子,一些曾在光竞争中被排斥的物种又得以重新占据生态位,导致物种均匀度和丰富度增加,主要是一些杂类草的增加。本研究结果同样符合中度干扰假说。低矮的一年生植物叉毛蓬、角果藜等数量明显增加。总之,放牧导致的直接采食和由此造成的竞争释放改变了群落的整体结构,使整个伊犁绢蒿群落的高度降低,群落中优势种被削弱,物种多样性增加;草地建群种伊犁绢蒿的优势度逐渐下降,角果藜和叉毛蓬呈现向群落主要成分演变的趋势,并且随着放牧强度的增强,这种趋势更加明显。 3.2群落多样性变化 放牧对植物群落既有积极作用,也有消极作用,是一种高度复杂的干扰方式[22]。随着放牧强度的增强,伊犁绢蒿群落Shannon-Wiener指数、Pielou指数、物种丰富度和Simpson指数整体呈上升趋势。由此可看出,适度的放牧有助于增加草地生物量和植物多样性,维持群落的稳定性。放牧干扰造成草地群落植物多样性发生变化,但植物多样性的影响程度随放牧强度的不同而异。自然干扰或人为干扰均容易在群落结构上形成断层,而这些断层就成为其他物种的侵入机会。放牧干扰一方面使群落在结构上出现断层,生境异质性增加;另一方面也使得优势种的竞争受到一些阻碍,为物种的侵入,特别是为一些原有物种种群的扩大提供了良好的媒介[22]。 过于频繁和剧烈的干扰(包括自然和人为)会破坏植物的生长环境,降低物种多样性。相反,在干扰很少时,群落由少数优势种占据,多样性也会降低。关于放牧对草地群落物种多样性的影响,本研究结果也支持中度干扰假说。中度放牧草地物种的Shannon-Wiener指数、Pielou指数、Simpson指数较高,这是由于适当放牧干扰下,牲畜的啃食抑制了优势种伊犁绢蒿的生长;同时一些较耐牧的物种(如角果藜、叉毛蓬和刺沙蓬等)的数量增加,抑制了优良牧草的竞争优势,使一些牲畜不喜食的杂类草如叉毛蓬、黄花软紫草的数量也随之增加,这样就提高了资源的利用率,增加了群落结构的复杂性。为了保持伊犁绢蒿在其群落中的优势地位,应该在其优势地位被取代的阶段采取有效保护措施。因此,建议对尚未因放牧过度而退化的草地进行合理利用,不能超出其草地载畜力阈值。 本研究从群落结构和群落多样性两个方面探讨了伊犁绢蒿群落对放牧干扰的响应。结果表明,放牧不仅使伊犁绢蒿在群落中的重要值降低,而且使群落中不同功能群所占比例也有所变化,杂类草和害草的比例上升,即适口性好、产量高的牧草数量减少,适口性差、较为耐牧的杂类草数量增多。这主要是由于这些自身具有较强的耐牧性的杂类草和害草是竞争释放的受益者,这就导致了物种多样性增加,群落结构和生物多样性的变化,群落结构改变,同时导致了放牧和禁牧草地的生态系统功能有所差异。结果反映出放牧不仅使伊犁绢蒿荒漠草地的生态功能发生改变,而且降低了草地的载畜量,并且使草地植被有所退化,但是如果在合理放牧强度下,伊犁绢蒿荒漠草地并不会进一步退化,这也得益于该地区分区轮牧和控制家畜数量的一些举措。 [1] 孙会忠,贺学礼.中国绢蒿属植物种质资源及其开发利用现状[J].西南林学院学报,2006,26(3):67-70,78. [2] 崔乃然.新疆主要饲用植物志(第二册)[M].乌鲁木齐:新疆科技卫生出版社,1994:295. [3] 朱进忠,吴咏梅.伊犁绢蒿荒漠不同退化阶段草地经济性状演变的分析[J].草业科学,2005,22(10):1-5. [4] 朱进忠.对新疆草业发展的思考(一)[J].新疆农业科学,2006,43(3):195-199. [5] 朱进忠.对新疆草业发展的思考(二)[J].新疆农业科学,2006,43(4):281-285. [6] 任继周.草业科学研究方法[M].北京:中国农业出版社,1998:42-48,207-211. [7] 马克平.生物群落多样性的测度方法[M].北京:中国科学技术出版社,1994. [8] Tilman D.Competition and biodiversity in spatially structured habitats [J].Ecology,1994,75(1):2-16. [9] 林慧龙,侯扶江,任继周.放牧家畜的践踏强度指标探讨[J].草业学报,2008,17(1):85-92. [10] 滕星,王德利,程志茹,等.不同放牧强度下绵羊采食方式的变化特征[J].草业学报,2004,13(2):67-72. [11] 王正文,邢福,祝廷成,等.松嫩平原羊草草地植物功能群组成及多样性特征对水淹干扰的响应[J].植物生态学报,2002,26(6):708-716. [12] 郝占庆,杨晓明.长白山北坡植物群落α多样性及其随海拔梯度的变化[J].应用生态学报,2002,13(7):786-789. [13] Whittaker R H,Niering W A.Vegetation of the Santa Catalina Mountains Arizona:A gradient analysis of the south slope[J].Ecology,1965,46:429-452. [14] Grime J P.Control of species diversity in herbaceous vegetation[J].Journal of Environmental Management,1973,1:151-167. [15] Tilman D,Wedin D,Knops J.Productivity and sustainability influenced by biodiversity in grassland ecosystems[J].Nature,1996,379:718-720. [16] Oksanen J.Predation,herbivory and plant strategies along gradients of primary productivity[A].In:Grace J B,Tilman D.Perspectives on Plant Competition[M].San Diego,CA:Academic Press,1990:445-474. [17] Grime J P.Benefits of plant diversity to ecosystems:Immediate,filter and founder effects[J].Journal of Ecology,1998,86:902-910. [18] Weiss S B.Cars,cows,and checker spot butterflies:Nitrogen deposition and management of nutrient-poor grassland for a threatened species[J].Conservation Biology,1999,13:1476-1486. [19] Tilman D.Species richness of experimental productivity gradients:How important is colonization limitation[J].Ecology,1993,74:91-98. [20] Oy-Meir I,Guttmann M,Kaplan Y.Responses of mediterranean grassland plants to grazing and protection[J].Journal of Ecology,1989,77:290-310. [21] Diaz S,Lavorel S,Ntyres S,etal.Plant trait responses to grazing——A global synthesis[J].Global Change Biology,2007,13:313-341. [22] Conell J H.Diversity in tropical rain forests and coral reefs[J].Science,1978,199(4335):1302-1310.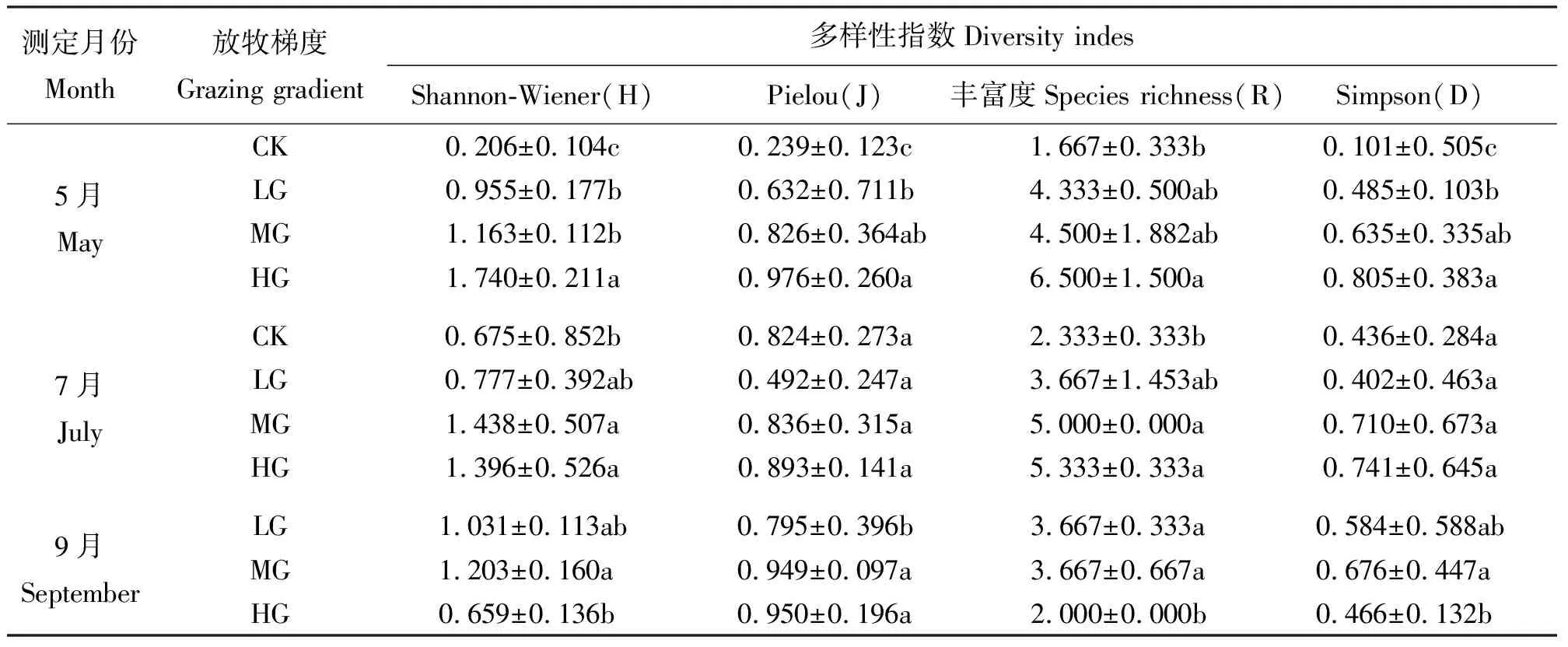
3 讨论
4 结论
