青藏公路沿线植被多样性特征及其与环境因子的关系
2013-04-25王光鹏斯贵才沈妙根张更新
王 建,王光鹏,斯贵才,沈妙根,张更新
(1.中国科学院青藏高原研究所,北京 100101; 2.中国科学院大学,北京100049;3.中国科学院地质与地球物理研究所兰州油气资源研究中心,甘肃 兰州 730000)
生物多样性与全球变化是目前各国面临的主要生态环境问题[1]。生物多样性的中心是物种多样性[2],物种多样性是一个群落结构和功能复杂程度的量度[3],它包括物种丰富程度、单位面积物种数目(物种密度)、特有物种比例等[4],是当前群落生态学中重要研究内容之一。研究物种多样性对认识及保护生物多样性具有重要意义。物种多样性的研究主要集中在多样性随环境梯度的变化情况[5-7]等方面。大尺度水平上主要表现为纬度梯度变异,小尺度水平上主要表现为海拔梯度的变异。多数理论研究认为,随纬度的增加物种多样性下降[8],形成这一格局的原因主要归结于生产力或其它与能量和环境有关的因素[9]。海拔梯度由于涉及温度、降水和太阳辐射等诸多环境因子而成为生物多样性梯度格局研究的又一个重要方面[10]。此外,近年来,小尺度水平上有关单个环境因子对物种多样性影响的研究也不断增多,在青藏高原主要表现为模拟控制试验尤其是增温和放牧试验[11-15]。但是,有关该区物种多样性中起决定性因素的环境因子的研究并不多见。
草地生态系统是陆地生态系统的重要组成部分。目前,在生物多样性的生态系统功能研究中,草地生态系统备受关注[16]。青藏公路地处世界屋脊,具有独特的自然地理环境和草原草甸群落特征。研究沿线植物物种多样性分布格局及其与环境因子的关系是人们关注的问题。目前,有关青藏公路沿线植被特征调查仅就特定的植被类型区做过一些植被多样性的研究[17-20],而对于完整的青藏公路沿线植物群落多样性的研究还未有系统报道。为了分析青藏公路沿线植物物种多样性分布格局及其与环境因子的关系,本研究对青藏公路沿线两侧植物进行样方调查分析,以期为研究该区生物多样性的保护与可持续利用提供科学参考。
1 研究方法
1.1研究区概况 青藏公路位于号称地球“第三极”的青藏高原,全长1 917 km,全线平均海拔在4 000 m以上,是世界上海拔最高的公路。沿线降水量较少,年均温较低,属于干旱寒冷的典型生态敏感区。该区干湿两季分明,冬长夏短,季节变化缓慢,太阳辐射强烈,日照时间长,年温差小,日温差大,受局部地形及小环境影响显著,不同地域温度差异很大[21]。青藏公路所跨范围可以分为6个生态分区[18,22-23],即西宁-天俊县山地草原区,天俊县-南山口山地荒漠区,南山口-昆仑山山地荒漠、山地草原区,昆仑山-唐古拉山高寒草甸、草原、荒漠区,唐古拉山-当雄高寒草甸区,当雄-拉萨灌丛草原区。草原草甸优势种主要有高山嵩草、藏嵩草、矮嵩草、紫花针茅和青藏苔草等,亚优势种主要有早熟禾、黄芪和委陵菜等,伴生种主要有冷地早熟禾、弱小火绒草、风毛菊和棘豆等[24-27]。
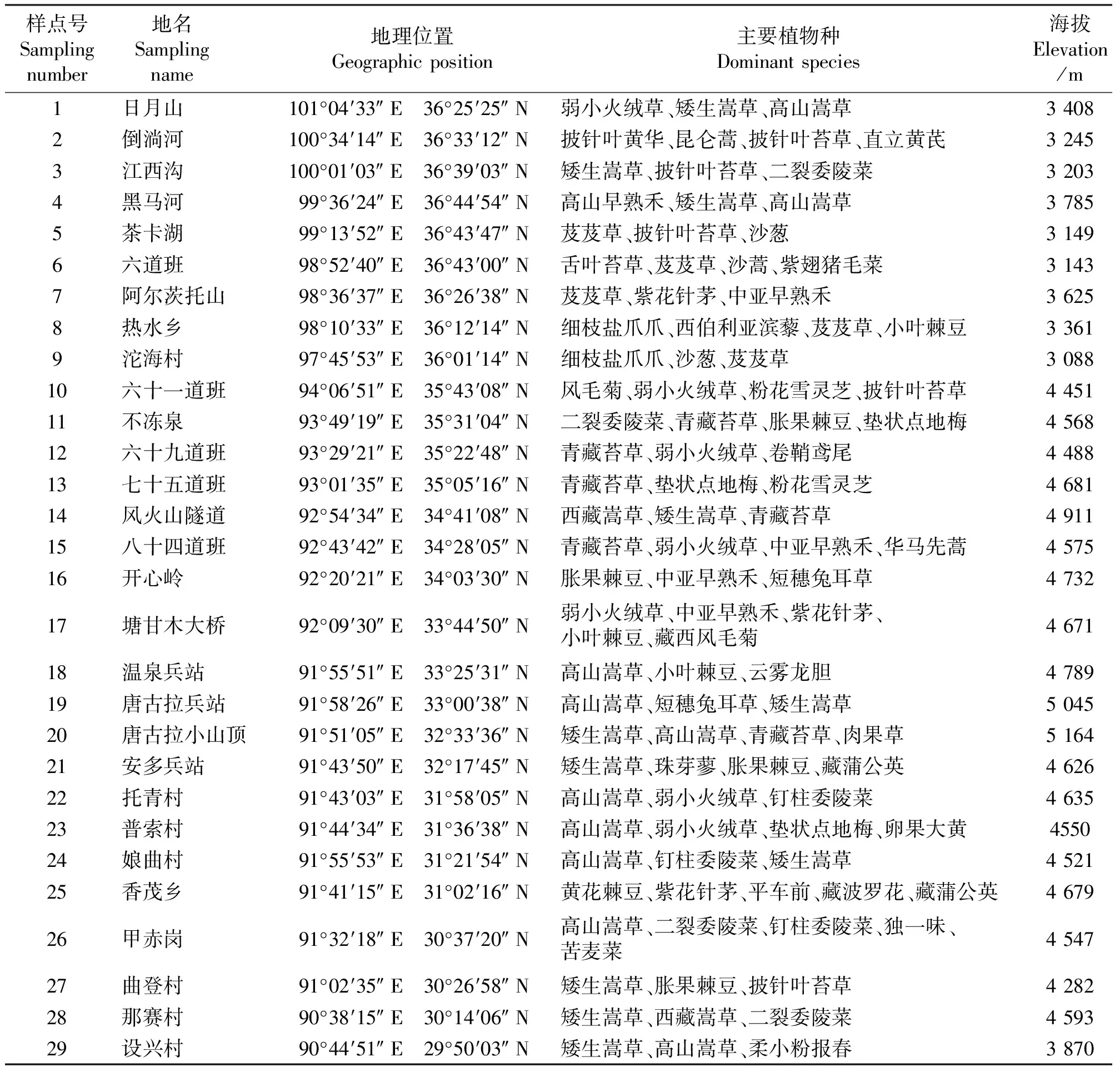
表1 样点概况Table 1 Generalization of transect in different latitude gradients of Tibetan Plateau
1.2样地设置 在植物生长最旺盛的8月,从西宁市出发,沿青藏公路进行植物群落调查,大约每隔50 km选择一处样地,一直到拉萨市共调查29处样地(图1)。在每处样地选取一个有代表性的10 m×10 m的大样方,在大样方中随机设置3个1 m×1 m的小样方,进行样方调查。样方调查采用针刺法,调查记录每个小样方内出现的物种数及每个物种的盖度、高度、密度和频度,同时,采集植物标本并记录每个样地所在位置的经纬度、海拔等信息,然后在室内准确鉴定植物标本,统计科属种及其组成。各样点经纬度、海拔高度及主要植物种详见表1。
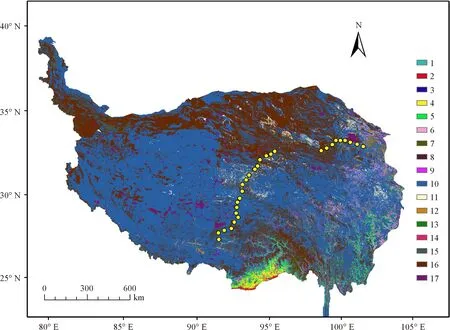
图1 样地分布图Fig.1 Location of the sample sites
1.3样地多样性计算 有关物种多样性的计算方法很多,本研究根据野外盖度、物种数、株数和频数等基础样方数据,运用Excel进行数据的初步处理,同时采用Sigma Plot 10.0、ArcGIS 9.3软件制图及SPSS 19.0统计分析软件进行相关分析和逐步回归等统计分析。物种多样性用丰富度指数、多样性指数和均匀度指数3类指数表征[28-32],其计算方法如下:
重要值(IV)=(相对频度+相对盖度)/2。
丰富度指数:R=S.
多样性指数:
Simpson指数:D=1-∑Pi2;Pi=Ni/N.
Shannon-Wiener指数:
H=-∑Pi·lnPi.
均匀度指数:
Pielou指数:P=H/lnS.
Alatalo指数:A=[1/(D-1)]/(eH-1).
式中,S为调查样方的物种数,由于个体数对于草本植物统计较为困难,本文采用各物种的重要值(IV)代替个体数进行计算[33-34],Ni为样方中第i种植物的重要值,N为样方中所有物种的重要值之和。
1.4气候数据来源和处理 对全国700多个标准气象台站1979-2010年间每月平均温度和降水数据进行克里格插值分析,然后根据样地的经纬度从插值结果中提取样地每月平均温度、降水量、空气比湿、短波辐射、长波辐射等数据,以各样点32年的气象数据平均值作为样地的温度和降水等数据。数据来源详见Yang等[35]和Chen等[36]。
2 结果与分析
2.1青藏公路沿线各样带植物群落物种多样性分析 由于青藏公路沿线气候、地形等条件的差异,沿线29个样点的物种多样性之间存在一定的差异。
2.1.1各样点总盖度和丰富度特征 沿线各样点1 m2样方平均植被总盖度和物种丰富度沿青藏公路呈先降低后升高然后趋于平稳的趋势(图2),而植被总盖度这种变化趋势更加明显。
青海湖南岸1~4号样点植被盖度为90%左右,5号样点由于靠近柴达木盆地东南缘,植被总盖度降至青海湖的1/3,而物种丰富度较1~4号样点有所减少。10~21号样点植被总盖度开始呈增加趋势,10~12号3个样点增加缓慢,14~16号3个样点增加迅速,17~21号5个样点增加缓慢,而物种丰富度最低点出现在海拔较高的风火山隧道(14号)。22号样点植被总盖度较17~21号5个样点降低较明显,物种丰富度略低于17~21号5个样点。那曲地区3个样点(23~25号)植被总盖度较17~21号5个样点降低较明显,较22号样点没有明显差异,物种丰富度没有明显变化趋势。进入当雄灌丛草原区,样点(26~29号)物种丰富度呈下降趋势,而草原植被总盖度并没有受到灌丛的影响,反而较那曲地区有所增加。其中那赛村样点(28号)植被总盖度偏低,可能由于该样点海拔(4 593 m)较其它3个样点高所致(图2)。
2.1.2各样点多样性指数特征 除个别样点波动幅度较大外,物种多样性辛普森指数和香农指数总体从1号样点到29号样点呈降低趋势(图3)。
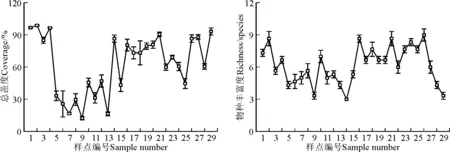
图2 青藏公路沿线植被总盖度和物种丰富度的变化Fig.2 Vegetation coverage and species richness along Qinghai-Tibet Highway
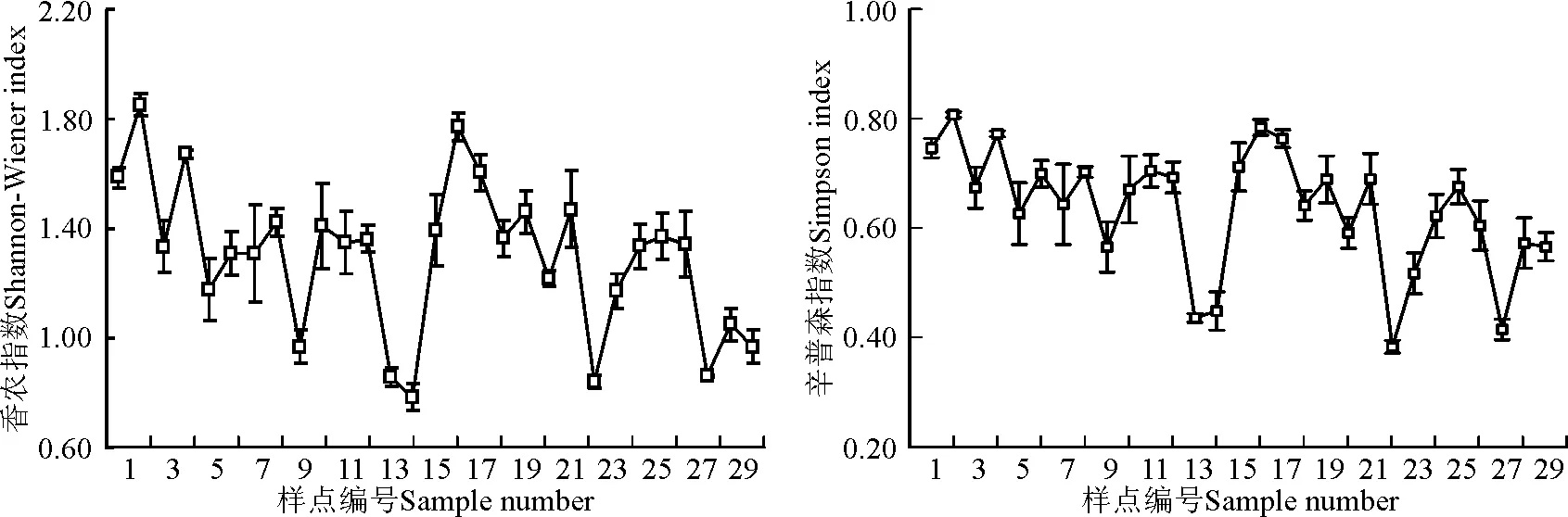
图3 青藏公路沿线物种多样性指数的变化Fig.3 Species diversity index along Qinghai-Tibet Highway
沿线样点辛普森指数介于0.38~0.81,总体呈降低趋势。1号样点辛普森指数为0.75,一直到12号样点辛普森指数缓慢降低,中间样点辛普森指数略有波动。在13号和14号样点辛普森指数急速下降至0.40左右,之后在15号样点约增加至14号样点的两倍,然后一直到22号样点呈现出递减的趋势。其中,15号到21号样点辛普森指数缓慢降低,22号样点降低幅度最大,约降至21号样点的1/2。到25号样点辛普森指数迅速增加至0.70左右,26号样点缓慢降低,到27号样点又迅速降低,最后两个样点又增加至26号样点水平。辛普森指数对于物种丰富度的敏感性较低,而对于物种均匀度的敏感性较高[37]。总体来说,13号、14号、22号和27号4个样点辛普森指数偏低,主要是由这些样点物种分布不均匀所致[38]。
香农指数与物种丰富度和分配均匀性有关,物种数目越多,多样性越大,物种间个体分配的均匀性越大,多样性越大。沿线样点香农指数所呈现的规律和辛普森指数基本一致,不同的是13号、14号、22号、23号和27号5个样点波动幅度较辛普森指数小,而16号和17号两个样点波动幅度较辛普森指数大。
2.1.3各样点均匀度指数特征 Pielou指数越大,物种分布越均匀,优势种越不明显,Alatalo指数则相反。Pielou指数在1~12号样点先保持稳定趋势,维持在0.73~0.88,快到风火山山顶时(13号)急速下降至0.60左右,之后增至风火山以前水平。过17号样点后开始下降,直到安多山(22号)降至最低,之后呈增加趋势,到25号样点增至最大。翻越念青唐古拉山(27号)时又出现明显降低趋势,之后两个样点又呈快速增加趋势,29号样点约增至27号的两倍。Alatalo指数变化趋势与Pielou指数相反,不同的是这种相反趋势较Pielou指数更加明显,主要体现在个别Pielou指数偏低的样点其Alatalo指数偏高,且波动很大,能达到其它样点的3~4倍(图4)。
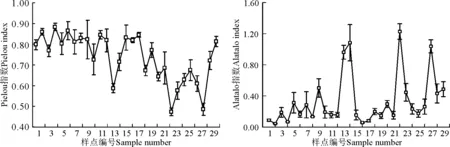
图4 青藏公路沿线物种均匀度指数的变化Fig.4 Species evenness index along Qinghai-Tibet Highway
青海湖南岸到柴达木盆地东南缘再到昆仑山区六十九道班(12号),均匀度指数保持稳定,波动幅度不大。可能原因是青海湖地区水热条件好,植被生长茂盛,而柴达木盆地东南缘地区气候干旱,只有少数耐旱植物生长,表现为植被覆盖度低,长势差等特点,两个地区植被优势种均不明显,植被分布较均匀。此外,高海拔地区物种均匀度指数波动较大。在风火山调查的14号样点、安多山调查的22号样点和念青唐古拉山调查的27号样点,都表现出植被Pielou指数随海拔升高而降低、Alatalo指数随海拔升高而升高的趋势。
2.2植被物种多样性与环境因子的关系
2.2.1物种盖度和丰富度与环境因子的关系 马涛等[23]研究青藏铁路格尔木至拉萨段植被样方时发现,物种丰富度与海拔高度之间呈现极显著线性正相关关系,说明海拔高度对物种多样性有着重要影响[39]。植被多样性不仅受海拔的影响,而且受其它环境因子的影响。青藏高原属高海拔、空气稀薄、气压低、气候寒冷的高原气候,其植被多样性受高寒气候条件下多种环境因子的影响。本研究分析了海拔高度、年降水量、生长季降水量、年均温、生长季均温、空气比湿、生长季空气比湿、短波辐射、生长季短波辐射、长波辐射、生长季长波辐射、压强和风速这些环境因子与青藏公路沿线物种多样性变化特征之间的关系及影响物种多样性变化特征的关键性环境因子(表2)。物种总盖度和物种丰富度受水分影响较大(图5)。相关性分析表明,物种盖度与年降水量、生长季降水量分别呈极显著正相关关系(P<0.01),与空气比湿呈显著正相关关系(P<0.05)。物种盖度与年降水量、生长季降水量分别呈显著正相关关系(P<0.05),而温度和其它环境因子与物种盖度和物种丰富度之间均未达到显著相关水平。
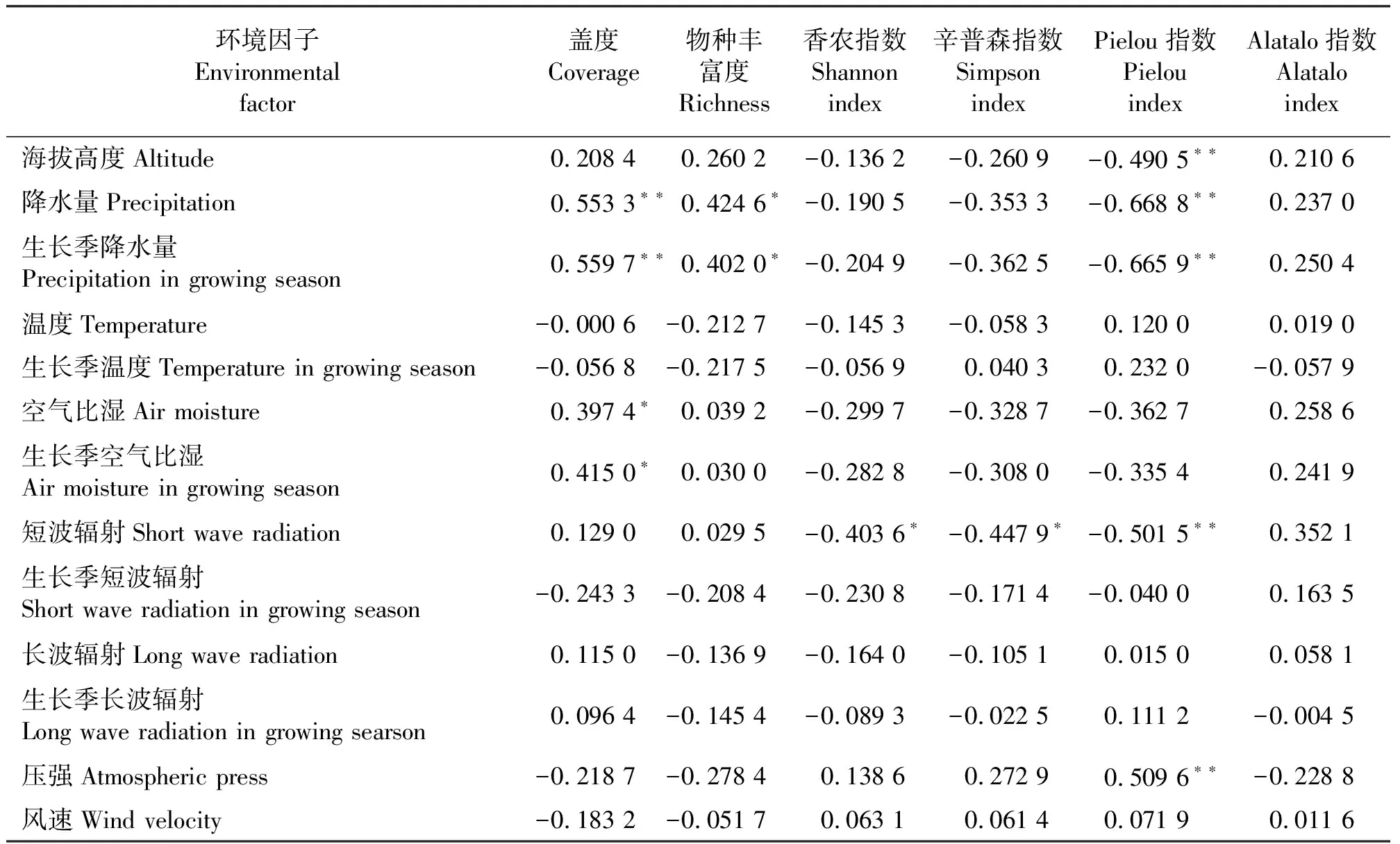
表2 青藏公路沿线环境因子与物种多样性指数间的相关系数Table 2 Coefficients of correlation between species diversity and environmental factors

图5 物种多样性与部分环境因子之间的相关关系Fig.5 Relationships between species diversity and some environmental factors
对物种丰富度、盖度分别与海拔高度、年降水量、生长季降水量、年均温、生长季均温、空气比湿、生长季空气比湿、短波辐射、生长季短波辐射、长波辐射、生长季长波辐射、压强和风速进行逐步回归分析,回归方程为:
C=349.19+0.25gp-1.60s;
其中gpr=0.89,sr=-0.48,P<0.05;
R=7.55+0.09p-0.08gp-1.073gh;
其中pr=5.22,gpr=-4.41,ghr= -0.58,P<0.05。
式中,C、gp、S、gpr、sr、R、p、gh和ghr分别为总盖度、生长季降水量、短波副射、生长季降水量标准偏回归系数、短波辐射准偏回归系数、物种丰富度、降水量、生长季比湿和生长季比湿标准偏回归系数。
逐步回归方程说明植被总盖度随生长季降水量增加而增加,随短波辐射增加而减少。标准偏回归系数绝对值大小表明生长季降水量对植被总盖度的贡献大于短波辐射。物种丰富度随降水量的增加而增加,随生长季降水量和生长季比湿的增加而减少,标准偏回归系数绝对值大小表明降水量对植被总盖度的贡献大于生长季比湿。
2.2.2物种多样性指数与环境因子的关系 物种多样性指数与短波辐射关系密切(图5)。香农指数和辛普森指数均与短波辐射呈显著负相关关系(P<0.05)。物种均匀度指数受多种环境因子共同作用。Pielou指数与海拔、降水量、短波辐射呈极显著负相关关系(P<0.01),与压强呈极显著正相关关系(P<0.05)。Alatalo指数与各环境因子相关性均没有达到显著水平。
此外,温度、长波辐射、风速和物种多样性指标之间关系均未达到显著水平。该结果表明在青藏高原地区,物种多样性受温度、长波辐射、风速这3个环境因子影响较小。
对香农指数、辛普森指数、Pielou均匀度指数和Alatalo均匀度指数分别和海拔高度、年降水量、生长季降水量、年均温、生长季均温、空气比湿、生长季空气比湿、短波辐射、生长季短波辐射、长波辐射、生长季长波辐射、压强和风速进行逐步回归分析,回归方程为:
H=4.33-0.01s;
其中sr=-0.41,P<0.05;
D=1.99-0.006s;
其中sr=-0.45,P<0.05;
E=0.98-0.001p;
其中pr=-0.67,P<0.05;
A=-2.81+0.01s;
其中sr=-0.35,P<0.10。
式中,H、s、sr、D、E、p、pr和A分别为香农指数、短波辐射、短波辐射标准偏回归系数、辛普森指数、Pielou均匀度指数、降水量、降水量标准偏回归系数和Alatalo均匀度指数。
回归方程说明植被物种多样性香农和辛普森指数随短波辐射增加而减少,物种Pielou均匀度指数随降水量的增加而减少,物种Alatalo均匀度指数随短波辐射的增加而增加。
3 讨论
从物种多样性和环境因子的相关性分析及逐步回归分析发现,水分因子是影响该区物种总盖度、物种丰富度和物种Pielou均匀度指数最主要的因子,而短波辐射是影响该区物种多样性指数香农指数和辛普森指数最主要的因子。这主要是由于青藏高原地区特殊的高海拔环境特征导致该区干旱缺水和太阳短波辐射强烈,其对植物的影响大于包括温度在内的其它环境因子,成为影响植物生长最主要的环境因子,进而决定物种多样性沿青藏公路的变化趋势。
沈振西等[39]通过野外控制实验,探讨矮嵩草草甸的群落特征对模拟降水的响应,发现增加降水可以显著增加物种多样性。杨元合等[40]通过对青海海北地区、唐古拉地区以及藏北地区的草原草甸物种丰富度研究和环境因子的相关性和逐步回归分析也表明,降水是影响物种丰富度最主要的环境因子。这些研究结果与本研究结果一致,能较好地说明水分对干旱区物种多样性的重要影响。有关太阳短波辐射对植物的影响的研究主要集中在对其形态结构以及生理特征的影响方面[41],而对物种多样性的影响方面还较少。但是,研究区强烈的太阳短波辐射使植物形态、结构和生理特征发生变化的同时,必定会改变物种的重要值,进而引起物种多样性的改变。
4 小结
除个别样点波动较大外,总体上看,随纬度的增加,青藏高原植被盖度呈先降低后升高的趋势,物种丰富度呈先降低后升高再降低的趋势,香农指数和辛普森指数随样点呈降低趋势,Pielou指数呈先降低后升高的趋势,Alatalo指数变化趋势与Pielou指数相反。
从物种多样性和环境因子的相关性分析及逐步回归分析发现,物种多样性的变化趋势主要受水分因子和太阳短波辐射因子控制。此外,一个地区的植被物种多样性是多种环境因子综合作用的产物,除了降水和短波辐射外,其它环境因子也对研究区物种多样性也有一定程度的影响。
[1] 常学向,赵文智,赵爱芬.祁连山区不同海拔草地群落的物种多样性[J].应用生态学报,2004,15(9):1599-1603.
[2] Tracy C R,Brussard P F.Preserving biodiversity-species in landscapes[J].Ecological Applications,1994,4(2):206-207.
[3] 黄忠良,孔国辉,何道泉.鼎湖山植物群落多样性的研究[J].生态学报,2000,20(2):193-198.
[4] 周红章.物种与物种多样性[J].生物多样性,2000,8(2):215-226.
[5] 高贤明,陈灵芝,黄建辉,等.秦岭太白山弃耕地植物群落演替的生态学研究.I.演替系列的群落基本特征及其生活型谱分析[J].生态学报,1997,27(3):253-259.
[6] 沈泽昊,方精云,刘增力,等.贡嘎山东坡植被垂直带谱的物种多样性格局分析[J].植物生态学报,2001,25(6):721-732.
[7] 温远光,元昌安,李信贤,等.大明山中山植被恢复过程植物物种多样性的变化[J].植物生态学报,1998,22(1):33-40.
[8] Brown J H,Lomolino M V.Biogeography,second edition[M].Sunderland,MA:Sinauer,1998.
[9] Currie D J.Energy and large-scale patterns of animal- and plant-species richness[J].American Naturalist,1991,137(1):27-49.
[10] Gaston K J.Global patterns in biodiversity[J].Nature,2000,405:220-226.
[11] 李英年,赵亮,赵新全,等.5年模拟增温后矮嵩草草甸群落结构及生物量的变化[J].草地学报,2004,12(3):236-239.
[12] 汪诗平,李永宏,王艳芬,等.不同放牧率对内蒙古冷嵩草原植物多样性的影响[J].植物学报,2001,43(1):89-96.
[13] Klein J A,Harte J,Zhao X Q.Experimental warming causes large and rapid species loss,dampened by simulated grazing,on the Tibetan Plateau[J].Ecology Letters,2004,7(12):1170-1179.
[14] Kudo G,Suzuki S.Warming effects on growth,production,and vegetation structure of alpine shrub:A five-year experiment in northern Japan[J].Oecologia,2003,135:280-287.
[15] 周华坤,赵新全,唐艳鸿,等.长期放牧对青藏高原高寒灌丛植被的影响[J].中国草地,2004,26(6):1-11.
[16] Schläpfer F,Schmid B.Ecosystem effects of biodiversity:A classification of hypotheses and exploration of empirical results[J].Ecological Applications,1999,9(3):893-912.
[17] 张镱锂,阎建忠.青藏公路对区域土地利用和景观格局的影响——以格尔木至唐古拉山段为例[J].地理学报,2002,57(3):253-266.
[18] 陈辉,李双成,郑度.青藏公路铁路沿线生态系统特征及道路修建对其影响[J].山地学报,2003,21(5):559-567.
[19] 鄢燕,张锦华,张建国.青藏公路沿线高寒草甸次生群落特征及生态修复[J].草地学报,2006,14(2):156-159.
[20] 丁明军,沈振西,张镱锂,等.青藏公路与铁路沿1981年-2001年植被覆盖变化[J].资源科学,2005,27(5):128-133.
[21] 周金星,Yang J,董林水,等.青藏铁路唐古拉山南段沿线植被多样性及盖度特征分析[J].北京林业大学学报,2008,30(3):24-30.
[22] 孙士云.青藏铁路沿线的生态环境特点及保护对策[J].铁道劳动安全卫生与环保,2002,29(3):121-124.
[23] 马涛,周金星.青藏铁路沿线植物群落物种多样性研究[J].水土保持学报,2012,32(2):261-266.
[24] 陈辉,李双成,郑度.基于人工神经网络的青藏公路铁路沿线生态系统风险研究[J].北京大学学报(自然科学版),2005,41(4):586-593.
[25] 徐宪立,张科利,庞玲,等.青藏公路路堤边坡产流产沙规律及影响因素分析[J].地理科学,2006,26(2):211-216.
[26] 汪双杰,周文锦.青藏公路沿线环境演化及环境保护对策[J].公路,2005(1):37-41.
[27] 杜小冰,陈建业,周剑.青藏公路建设对高寒草原生态的影响及治理对策[J].交通标准化,2011(10):91-94.
[28] 王文颖,王启基.高寒嵩草草甸退化生态系统植物群落结构特征及物种多样性分析[J].草业学报,2001,10(3):8-14.
[29] 孙儒泳,李庆芬,牛翠娟.基础生态学[M].北京:高等教育出版社,2002:136-137.
[30] Margalef R.Perspectives in Ecological Theory[M].Chicago:University of Chicago,1968.
[31] Hill M,Bunce R,Shaw M.Indicator species analysis,a divisive polythetic method of classification,and its application to a survey of native pinewoods in Scotland[J].The Journal of Ecology,1975,63(2):597-613.
[32] Wang C T,Long R J,Wang Q J,etal.Effects of altitude on plant species diversity and productivity in an alpine meadow,Qinghai-Tibetan plateau[J].Australian Journal of Botany,2007,55(2):110-117.
[33] Peet R K.The measurement of species diversity[J].Annual Review of Ecology and Systematics,1974,5:285-307.
[34] 马克平.生物多样性的原理和方法[M].北京:中国科学技术出版社,1994:141-165.
[35] Yang K,He J,Tang W J,etal.On downward shortwave and longwave radiations over high altitude regions——Observation and modeling in the Tibetan Plateau[J].Agricultural and Forest Meteorology,2010,150:38-46.
[36] Chen Y Y,Yang K,He J,etal.Improving land surface temperature modeling for dry land of China[J].Journal of Geophysical Research,2011,116,D20104,doi-10.1029/2011JD015921.
[37] 董林水,张旭东,周金星,等.青藏铁路沿线北段植被物种丰富度及盖度的动态变化[J].长江流域资源与环境,2008,17(4):551-556.
[38] 许晴,张放,许中旗,等.Simpson指数和Shannon-Wiener 指数若干特征的分析及“稀释效应”[J].草业科学,2011,28(4):527-531.
[39] 沈振西,周兴民,陈佐忠,等.高寒矮嵩草草甸植物类群对模拟降水和施氮的响应[J].植物生态学报,2002,26(3):288-294.
[40] 杨元合,饶胜,胡会峰,等.青藏高原高寒草地植物物种丰富度及其与环境因子和生物量的关系[J].生物多样性,2004,12(1):200-205.
[41] 赵新全.高寒草甸生态系统与全球变化[M].北京:科学出版社,2009:53.
