干旱时期放养紫胶虫对马鹿花光合生理生态特征影响研究
2013-04-11黄小波李福强杨贵成王鲜艳
黄小波,李福强,杨贵成,谷 勇 ,王鲜艳,吴 昊
(1.中国林业科学研究院资源昆虫研究所,云南 昆明 650224;2.西南林业大学 环境科学与工程学院,云南 昆明650224;3.怒江州林业局,云南 怒江 673100;)
马鹿花(Pueraria wallichii)属于蝶形花科(Papilionaceae),葛藤属(Pueraria)。原产于热带、南亚热带的疏林中,具有喜温、耐旱、耐贫瘠、生长快、萌发力强、繁殖栽培容易、种子蛋白质含量高、病虫害少等特点,可作为干热河谷地区绿化荒山、保持生态环境的造林先锋树种;又可作为紫胶虫的优良灌木寄主植物,其产胶性能优良,投产早,产胶量高,胶被厚,可多次放养利用;加之其生长快,萌发力强,可作为优良的薪炭材树种;而种子蛋白质含量高达33%,又可作为优良的木本蛋白饲料[1]。1999年云南省林业厅把马鹿花列为澜沧江、珠江生态防护林工程推广树种之一;随后也列为金沙江、怒江等防护林的造林树种之一[2],并大面积植造。本文拟通过对放养紫胶的马鹿花与未放胶的马鹿花的光合生理生态特征比较,为干热河谷地区植被恢复和紫胶产业的经营提供基础理论依据。
1 试验地概况
马鹿花光合测定样地为人工造林地,林龄 12 年,位于东经 98°50'34.4″,北纬 25°53'12.1″,海拔897.0 m,属南亚热带季风气候,≥10℃活动积温7374℃,年平均日照时数1889.2 h,日照百分率43%,年均温20.1℃,最冷月月均温9.1℃,最热月月均温24.8℃,年极端最高温度39.8℃,年极端最低温度3.7℃,全年无霜,年平均降雨量1001.6mm;土壤为山地红壤,母质岩为石灰岩;群落结构为干热河谷稀树灌草丛,原生植物种类有云南松(Pinus yunnanensis Franch.)、番木瓜(Carica papaya L.)、大叶榕(Ficus altissima B.)、攀枝花(Bombax malabaricum DC.)、余甘子(Phyllanthus emblica L.)、羊蹄甲(Bauhinia claviflora L.)、大叶千斤拔[Flemingia macrophylla(Wiild.)Prain]、旱芦苇(Eremopogon Stapf L.)、扭黄茅[Heteropogon contortus(L.)Beauv.]、白茅[Imperata cylindrical var.major(Nees)C.E.Hubb.]、黄背草[Themeda triandra var.japonica(Willd.)Makino]、飞机草(Eupatorium odoratum L.)、紫茎泽兰(Eupatorium adenophorum Spreng.)等。
2 材料与方法
2011年12月下旬,选择晴朗无风的天气,使用美国Li-COR公司生产的Li-6400便携式光合测定系统,对净光合速率[Pn,μmol/(m2·s)、蒸腾速率[Tr,mmol/(m2·s)]、气孔导度[Gs,mmol/(m2·s)]、胞间 CO2浓度(Ci,μmol/mol)、大气 CO2浓度,以及气温(Tair)、叶温(Tleaf)、相对湿度(RH)、光合有效辐射(PAR)等相关大气因子进行测定。测定时,分别选取放养紫胶虫和未放养紫胶虫长势良好的马鹿花植株各15株作为测定株,每株选择中部不同方向具有代表性的马鹿花叶片5片,做好标记。每次测定部位相同,3次重复。于12月24—26日连续测定3 d,每天自08:00开始,每2 h测定1次,至18:00结束。测定所记录的数据导入计算机后,用Microsoft Excel 2003与SPSS 17.0软件分析并绘图。

3 结果与分析
3.1 环境因子的日变化
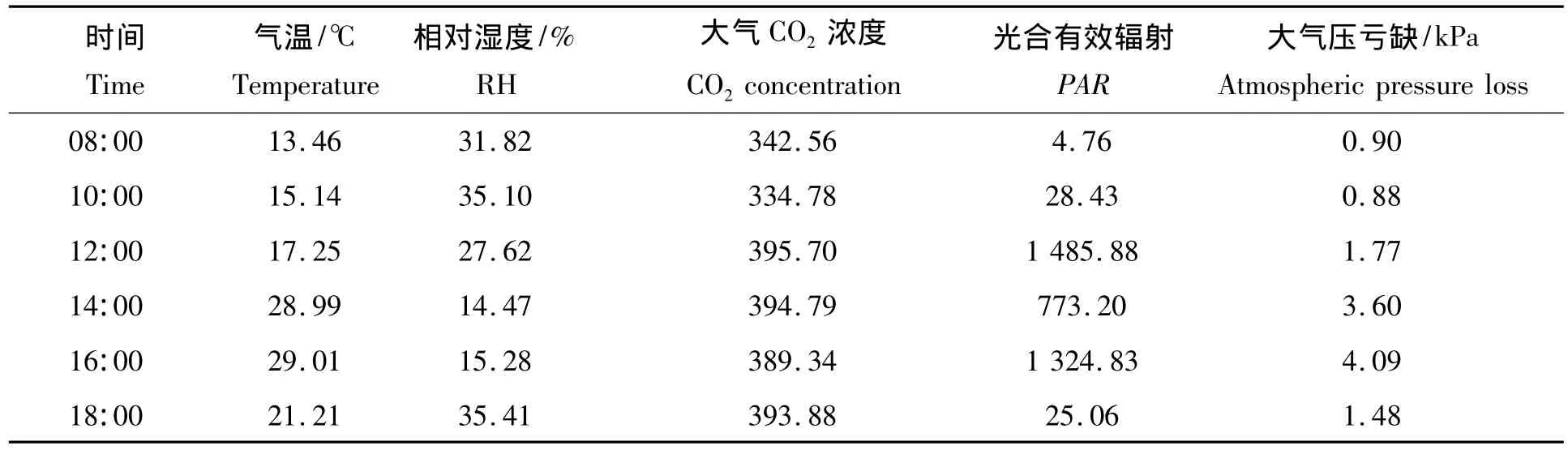
表1 环境因子的日变化Tab.1 The diurnal variation of environmental factors
环境因子的日变化如表1所示。由表1可以看出,测定期间温度逐渐升高,在16:00达到最大,随后开始降低。空气相对湿度则呈先升高后降低后升高的趋势。大气CO2浓度日变化呈“W”型趋势,即CO2浓度先降低再升高,在12:00达到最大,在10:00与12:00之间浓度上升最快,14:00到16:00有所下降,16:00到18:00再次上升,下降与上升变化比较平缓。在08:00开始光合有效辐射持续上升,在12:00左右达到最大,为1485.88 μmol/(m2·s),然后逐渐下降,在14:00 左右出现1 个波谷,为773.20 μmol/(m2·s),随之急剧上升,在16:00又出现1个峰值,为1324.83 μmol/(m2·s),然后又急剧下降,基本呈“M”型变化趋势。
3.2 放养紫胶虫与未放养紫胶虫马鹿花净光合速率与气孔导度的日变化比较
在自然条件下,植物光合作用的日变化大体有2种类型,单峰和双峰型[3],双峰型两峰之间形成一个中午的低谷,呈现“午休”现象。如图1a所示,马鹿花光合速率的日变化为双峰型,两峰值分别出现在12:00左右和16:00左右,分别为4.43 μmol/(m2·s)和2.13 μmol/(m2·s),其中第1 个峰值明显大于第二个峰值,在14:00左右达到最低值,为1.88 μmol/(m2·s)。有研究表明,光合作用出现“午休”现象主要是因为大气湿度变低的缘故[4]。而放养紫胶虫的马鹿花净光合速率(Pn)却区别于未放胶马鹿花,在8:00至10:00之间,略显下降趋势,10:00到12:00迅速上升并达到最大值1.78 μmol/(m2·s),之后平缓下降至观测结束。整个观测期间,放养紫胶虫的马鹿花净光合速率(Pn)明显小于未放胶的马鹿花净光合速率(Pn)。
植物叶片的气孔导度(Gs)是植物指示遭受水分和光热胁迫的敏感因子,也是植物自我调节的一种生理机制[4]。从图1a和图1b可以看出,马鹿花的净光合速率(Pn)与气孔导度(Gs)变化趋势基本吻合,这正好拟合Ball等提出的Ball-Berry模型[5]。该模型认为气孔导度和光合速率之间存在着线性关系。随着光照的加强,刺激气孔开放,引起净光合速率(Pn)的迅速上升,随着太阳辐射的进一步加强,气孔关闭,气孔导度(Gs)迅速下降。相比之下,放养紫胶虫的马鹿花各个时刻的气孔导度(Gs)比较小,变化较为平缓,且各个时刻的气孔导度(Gs)均小于未放养紫胶虫的马鹿花。

图1 放养紫胶虫与未放养紫胶虫马鹿花叶片净光合速率和气孔导度的日变化Fig.1 The diurnal variation of Pn,Gsof Pueraria wallichii leaves for stocking lac and no stocking lac

图2 放养紫胶虫与未放养紫胶虫马鹿花叶片胞间CO2浓度与气孔限制值的日变化Fig.2 The diurnal variation of Ci,Lsof Pueraria wallichii leaves for stocking lac and no stocking lac
3.3 放养紫胶虫与未放养紫胶虫马鹿花叶片胞间CO2浓度与气孔限制值的日变化比较
气孔限制值(Ls)反映植物叶片对大气CO2相对利用率的大小[6]。如图2b所示,两类马鹿花气孔限制值(Ls)日变化起始值都很低,未放养紫胶虫的马鹿花气孔限制值(Ls)从08:00到12:00急剧上升至最大值,放养紫胶虫的马鹿花气孔限制值(Ls)从08:00到10:00之间稍微下降,10:00开始急剧上升至12:00达到最大值,之后二者的气孔限制值(Ls)在一个高点值上平缓降低,都呈单峰的变化趋势。根据Farquhar和Sharkey[6]的报告,当光合速率和胞间CO2浓度变化方向相同,且气孔限制值增大,才可以认为光合速率的下降主要是由气孔因素导致,当光合速率和胞间CO2浓度变化方向相反,气孔限制值减小,则净光合速率下降归因于叶肉细胞同化能力的降低。结合图1a和图2可以看出,当胞间CO2浓度和净光合速率变化方向相反,Ci增高,且Ls呈下降趋势,这就表明Pn午间降低并不是由于气孔限制因素引起叶肉细胞间CO2不足造成的,而是主要由非气孔因素引起的叶片光合能力降低造成的。观测期间,放养紫胶虫的马鹿花叶片胞间CO2浓度高于未放养马鹿花的胞间CO2浓度值。
3.4 放养紫胶虫与未放养紫胶虫马鹿花叶片蒸腾速率与水分利用效率的日变化比较
蒸腾速率(Tr)的强弱是表明植物水分代谢的一个重要的生理指标[7]。蒸腾速率(Tr)既受外界因子的影响,也受植物体内部结构和生理状况的调节[8]。气孔导度对于土壤水分的变化及其敏感,气孔导度的变化进而影响蒸腾速率[4]。如图3a所示,放胶与未放胶的马鹿花蒸腾速率(Tr)变化趋势基本保持一致,呈单峰型。但未放养紫胶虫马鹿花蒸腾速率(Tr)各时刻的值均大于放养紫胶后马鹿花蒸腾速率(Tr)值,且二者出现的峰值时刻不同,未放胶马鹿花蒸腾速率(Tr)峰值出现在16:00前后,为1.29mmol/(m2·s),而放养紫胶的马鹿花蒸腾速率(Tr)峰值出现在14:00前后,为0.88mmol/(m2·s),二者的蒸腾速率(Tr)在16:00过后急剧下降。放养紫胶虫后马鹿花表现较低的蒸腾速率主要因其具有较低的气孔导度(图1b)。
植物叶片净光合速率是影响植物叶片水份利用效率WUE的一个重要生理因子[9]。当植物受到外界环境胁迫时,叶片光合机构受到伤害,净光合速率降低,从而使WUE下降[10]。高WUE常常被作为植物生存和分布能力强弱的表征[11]。结合图1a和图3b所示,马鹿花净光合速率由于气孔关闭出现午休现象急剧下降时,其WUE在12:00至14:00之间也急剧下降。放养紫胶虫的马鹿花WUE值除08:00与18:00前后略大于未放养紫胶虫的马鹿花WUE外,其他均小于未放养紫胶虫马鹿花WUE。
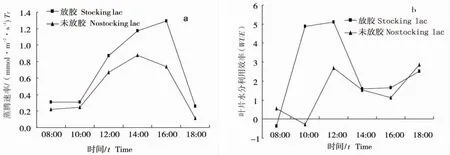
图3 放养紫胶虫与未放养紫胶虫马鹿花叶片蒸腾速率与水分利用效率的日变化Fig.3 The diurnal variation of Tr,WUE of Pueraria wallichii leaves for stocking lac and no stocking lac
3.5 放养紫胶虫与未放养紫胶虫马鹿花叶片温度与光能利用效率的日变化比较
由图4a可以看出,放养紫胶虫与未放养紫胶虫的马鹿花叶片温度基本随着光照强度的变化而变化,未放养紫胶虫马鹿花叶片温度持续升高,在16:00左右达到最高值。之后,随着光照强度下降,叶温也随之下降,放养紫胶虫马鹿花叶片温度从观测开始到10:00前后有所下降,然后上升迅速,在14:00前后达到最高值,之后,叶温逐渐下降。整个过程,二者的叶片温度曲线互有交叉。
由图4b可以看出,放养紫胶虫与未放养紫胶虫的马鹿花光能利用率在08:00至12:00之间的变化趋势正好相反,放养紫胶虫马鹿花光能利用率在08:00至12:00左右呈现先下降后上升的变化趋势,而未放养紫胶虫的马鹿花光能利用率却在这段时间呈现先上升后下降的变化趋势。在12:00至18:00,二者变化趋势基本保持一致,但未放养紫胶虫的马鹿花光能利用率大于放养紫胶虫的马鹿花光能利用率的值。

图4 放养紫胶虫与未放养紫胶虫马鹿花叶片温度和光能利用率的日变化Fig.4 The diurnal variation of Tleaf,LUE of Pueraria wallichii leaves for stocking lac and no stocking lac
3.6 Pn与各因子之间的相关性分析
为了比较放养紫胶虫与未放养紫胶虫马鹿花植株的光合作用受哪些主要因子的影响,将测定的Pn与同步测定的各因子之间进行相关性分析(表2)。结果表明:放养紫胶虫的马鹿花Pn与Tr、PAR呈显著正相关,与Ci呈显著负相关,而未放养紫胶虫的马鹿花Pn只与Ci呈显著负相关,这与孙启忠[12]等的“光合作用速率高,蒸腾速率也较高”的结论有很大差异,这或许与冬季太阳辐射不强,温度不高有关,而放养紫胶虫后,随着马鹿花植株营养物质的消耗,使放养紫胶虫后的马鹿花生理生态极为敏感。无论放养紫胶虫还是未放养紫胶虫的马鹿花Pn与Gs无显著相关,而与Ci呈显著负相关,这也证明了观测期间,影响马鹿花Pn下降的是非气孔因素。

表2 放养紫胶与未放养紫胶马鹿花Pn与各因子之间的相关性分析Tab.2 Correlation coefficients between Pnand factors affecting Pnfor stocking lac and no stocking lac
4 结论和讨论
“光合午休”现象比较普遍,并不是某种植物特有[13]。实际上是植物在长期进化过程中适应干旱环境而产生的一种生理现象[8]。“光合午休”按其影响因子大体可以分为两类。外部因素主要有低湿高温引起的高饱和差[14],空气CO2浓度的降低[14]等。内部因素主要来自植物本身的生理特性,如气孔关闭[15]、叶温升高[16]、光合产物的积累[17]以及水分亏损[18]等。云南怒江峡谷地区有明显的旱季和雨季之分,在旱季,未放养紫胶虫的马鹿花的光合速率的日变化基本为双峰型(图1a),两峰值分别出现在12:00和16:00前后,且第1个峰值远大于第2个峰值,在14:00前后出现最低值,有较明显的“午休”现象,但“午休”现象出现较晚,这可能与怒江峡谷地区的区域环境有关,呈南北走向的怒江大峡谷在观测期间日出时间和日落时间分别为11:34和17:12。14:00时太阳照射差不多与西坡呈垂直角度,此时的大气湿度最低,这与许大全[4]的研究正好吻合,再者,我国所用时间为北京时间,怒江地处滇西,时差导致最高温度滞后也是“午休”现象出现较晚的一个重要原因。而放养紫胶的马鹿花光合速率却没有光合“午休”,这可能与放胶后,紫胶虫在整个生活过程中会从寄主树上获得全部营养物质所致,根据林秀兰等[19]的研究,挂胶后的寄主树树皮冷、热水抽取物、碱抽取物、戊聚糖、全氮含量均略有下降。营养物质的丢失或许导致了放养紫胶后马鹿花生理的紊乱。
综上所述,与未放养紫胶虫马鹿花相比,放养紫胶虫马鹿花具有较低的净光合速率,较低的蒸腾速率、气孔导度、气孔限制值和较低的水分利用效率,较高的胞间CO2浓度。另外,相关性研究表明,Pn与Gs无显著相关,而与Ci显著负相关,说明影响马鹿花Pn下降的是非气孔因素,这可能是冬季暗反应中的许多酶活性和酶促反应速率降低,导致还原CO2的暗反应速度下降所致。同一环境因子对于放养紫胶虫和未放养紫胶虫马鹿花Pn的影响不同,冬季未放养紫胶虫与胞间CO2浓度关系最为密切,而放养紫胶虫后,Pn除受胞间CO2浓度影响外,还受蒸腾速率Tr和光合有效辐射PAR的影响较大。
[1]谷勇,邹恒芳,周榕,等.施肥放养紫胶对马鹿花结籽的影响[J].西南林学院学报,2002(12):38-40.
[2]张建云,安银岭,李正红,等.马鹿花化学成分研究初探[J].林产化学与工业,2001(6):67-70.
[3]Schulze E D,Hall A E.Encyclopedia of plant physiology[M].Springer-Verlag Berlin,1982:181.
[4]许大全.光合作用“午睡”现象的生态生理与生化[J].植物生理学通讯,1990,14(6):5-10.
[5]Ball J T,Woodrow I E,Berry J A.A model predicting stomatal conductance and its contribution to the control of photosynthesis under different environmental conditions[M].Biggens J.Progress in photosynthesis research[J].Dordrecht:Martinus Nijhoff Publishers,1987:221 -224.
[6]Fqrquhar G D,Sharkey T D.Stomata conductance and photosynthesis[J].Ann Rev Plant Physiologic,1982,33:317 -345.
[7]王旭军,吴际友,唐水红,等.红榉光合生理特性日变化规律[J].湖南林业科技,2012,39(1):10-13.
[8]周秋平,程积民,万惠娥.本氏针茅与柳枝稷光合生理生态特征比较研究[J].水土保持通报,2009,29(2):129-133.
[9]李机密,黄儒珠,王健,等.陆生植物水分利用效率[J].生态学杂志,2009,28(8):1655 -1663.
[10]Gimenez C,Mitchell V J,Lawlor D W.Regulation of photosythetic rate of two sunflow hybrids under water stress[J].Plant Physiol,1992 ,98(2):516 -524.
[11]Kristina A,Schierenbeck G,John D M.Seasonal and diurnal patterns of photosynthetic gas exchange for Lonicera serrrerurens and L.japonica(Caprifoliaceae)[J].American Journal of Botany,1993,80:1292 -1299.
[12]孙启忠,桂荣,那日苏,等.赤峰地区不同生长年限沙打旺生产力的研究[J].中国草地,1999(5):29-34.
[13]Neil C T,Paul J K.Adaptation of plants to water and higher temperature stress[M].Wiley-Interscience,1980:139.
[14]廖建雄,王根轩.谷子叶片光合速率日变化及水分利用效率[J].植物生理学报,1999,25(4):362-368.
[15]丁小球,胡玉佳,王榕楷.3种草坪草净光合速率和蒸腾速率的日变化特点研究[J].植物生理学报,1997,23(1):62-65.
[16]孟庆伟,邹琦,许长成,等.小麦和大豆叶片的气孔不均匀关闭现象[J].草业科学,2003,23(4):17-20.
[17]张永亮,王建丽.单播与混播下的杂花苜蓿与无芒雀麦光合生理生态特征分析[J].草业科学,2007,24(1):53-60.
[18]姜华,毕玉芬.燕麦叶片光合变化初探[J].草原与草坪,2002(1):34-36.
[19]林秀兰,苏小青.放养紫胶虫对大叶相思树皮提取物质量的影响[J].福建林学院学报,2001,21(1):65-67.
