人类活动对高寒矮嵩草草甸的碳容管理分析
2013-04-10林丽李以康张法伟郭小伟曹广民
林丽,李以康,张法伟,郭小伟,曹广民
(中国科学院西北高原生物研究所,青海 西宁810001)
青藏高原作为欧亚大陆最高最大的地貌单元,对全球气候变化和人类活动十分敏感,具有独特的研究价值。过去30年里,极度退化的高原草地生态系统释放大约3.02Pg贮存碳[1],其中土地利用格局和强度、土壤侵蚀、人类管理是系统碳源汇转化的重要因素[2,3]。高寒矮嵩草(Kobresiahumilis)草甸是青藏高原广泛分布的重要植被类型,是集生产、生态、生活三生功能的重要自然资源[4]。人类活动对该类草甸最大的影响是放牧,其演替的主流方向是退化[5,6]。随着高寒矮嵩草草甸放牧强度的加大,地上植物有机碳储量显著降低,严重影响了草地的生产服务能力[7];但植物-土壤系统有机碳储量则先增高后减少[6-9],草地生产服务最佳阶段同草地碳储量最高阶段不同步,说明,必须寻找这样一个或多个阶段,可以使得草地生产服务能力和碳储能力双赢(即适宜碳容管理阶段)。对极度退化的高寒矮嵩草草甸(黑土滩-杂类草次生裸地)建植人工草地是实现草地快速恢复的必要手段[4]。建植人工草地是黑土滩-杂类草次生裸地最常用的恢复措施,但该草地建植5年左右会出现优良牧草退化,毒杂草大量繁殖等“退化”现象[10-13],且很难通过灭杂等措施根治。如何对该类型草地进行碳容管理,以增强其碳容功能及生产服务功能,成为目前亟待解决的问题。
草地生态系统碳容管理的目标是通过合理的管理活动来调控草地生态系统碳循环过程及其影响因素,维持草地生态系统碳收支平衡[14]。受到外界不确定因素的影响,草地生态系统是一个动态变化的过程,这一过程直接影响了土壤—植被—大气系统—水循环之间的碳交换过程与机制[15],草地生态系统本身同干扰存在一定的互馈作用[16-19],如何探索天然草地的最佳利用模式,如何对极度退化天然草地进行有效的恢复,是目前高寒矮嵩草草甸适宜碳容管理的重要研究方向。据此选取高寒矮嵩草草甸典型退化演替及人工草地恢复系列为研究对象,分析2种过程碳储能力的变化,探索该类型草地的适宜碳容管理方法、阶段,为草地保护、建设及碳贸易谈判提供理论指导。
1 材料与方法
1.1 样地的选择
前人对高寒矮嵩草草甸演替过程长期定位观测发现:高寒矮嵩草草甸退化演替广泛存在于青藏高原的不同地域,受到人类活动强度或牧民经营策略的影响,各地区草地现今所处的演替阶段出现明显的分异,甚至在局域出现个别阶段的缺失[7,8]。本研究以此为基础,以《1∶100万中国草地资源图集》(1992年出版)初步划分高寒矮嵩草草甸分布区域,以实地调查为基础确定不同的演替阶段,并在该草地周围1km范围内寻找演替系列中其他演替阶段(通常以围栏为界,减弱环境因素对植被类型的影响力),据此选取的采样地分布于青海省海北州、果洛州、玉树州和西藏藏北高原32个县(乡),96个样地,收集整理及测定样地植物群落数量特征(植被盖度、生物量等13个指标)及土壤理化特征(有机碳储量、容重等6个指标),建立草地生态系统归属性判别指标体系,通过Fisher’s线性判别及欧式聚类划分出草地退化过程几个关键阶段,根据群落类型及地表特征分别命名为禾草-矮嵩草草甸,矮嵩草草甸,正常小嵩草(Kobresiapygmaea)草甸、小嵩草草甸草毡表层开裂期,小嵩草草甸草毡表层剥蚀期,黑土滩杂类草次生裸地,这种划分方式同已有研究结果吻合[8]。人工草地恢复系列根据人工草地建植年限确定,分别为黑土滩-杂类草次生裸地、人工草地建植初期、5年人工草地、14年人工草地。以2种演替系列为研究对象,以空间尺度代替时间尺度,选取欧式聚类值最接近的1~3个样地为研究样地[20]。
1.2 样地概况
禾草-矮嵩草阶段(HA):研究区域位于青海省海北州门源县风匣口,N 38°01.35′,E 100°40.26′,海拔3 170m;果洛州达日县满掌乡,N 33°16′,E 100°28′,海拔3 946m。样地概况:草地类型均属于禾草-莎草-杂类草型草地,草地有明显的分层结构,草地总盖度达到96%以上,禾本科、杂类草和莎草科植物优势度分别为51.3%,48.7%和18.5%。
矮嵩草草甸(AS):研究区域位于青海省海北州皇城乡,N 37°39.88′,E 101°10.75′,海拔3 227.6m;青海省玉树州巴塘滩,N 35°51.210′,E 96°59.656′,海拔3 907m;青海省门源县鄂博乡,N 37°56.342′,E 100°57.8′,海拔3 428m。样地概况:草地较为平坦,无裂缝,地面秃斑少,鼠类活动较少,地表总盖度大于90%,层片结构明显,下层为矮嵩草、小嵩草,上层多为禾本科植物,杂类草种类丰富。草毡层厚度为2.13cm左右。
正常小嵩草草甸(XS-1):研究区域位于青海省海北州皇城乡,N 37°39.876′,E 101°10.748′,海拔3 227.6m;青海省海北州门源县扣门子,N 37°35.018′,E 101°16.578′,海拔3 280m;青海省海北州门源县鄂博乡,N 37°56.342′,E 100°57.88′,海拔3 428m。样地概况:小嵩草草甸,且小嵩草死亡后形成的黑斑面积占13.2%左右,小嵩草斑块周围的塌陷面积达6.6%,斑块以莎草科植物为主,草层高度为2cm左右,地面平坦程度低,草地塌陷区域以禾本科植物为主。
小嵩草草甸草毡表层开裂期(XS-2):研究区域位于青海省海北州皇城乡,N 38°03.87′,E 100°26.30′,海拔2 994m;青海省门源县鄂博乡,N 37°56.34′,E 100°57.88′,海拔3 428m。样地概况:小嵩草草甸,草地斑驳不平,存在大量由小嵩草死亡后形成的黑斑块,其面积达到18.1%,斑块周边草皮塌陷,面积达到24.6%,草地以莎草科植物为优势,没有明显的分层现象,草毡表层厚度大约4.3cm。
小嵩草草甸草毡表层剥蚀期(XS-3):研究区域位于果洛州达日县、甘德交界处,N 33°53.34′,E 99°49.050′,海拔4 233m;果洛州玛沁县优云乡,N 34°26.09′,E 99°12.19′,海拔4 321m。样地概况:小嵩草草甸,小嵩草斑块退化呈剥蚀状态,剥蚀度60%~70%,小嵩草死亡黑斑占残留草皮的40%,地表裂缝较多,草毡表层发生破碎,均不连片,且残余草皮以小嵩草为优势,存在大量小嵩草死亡后形成的黑斑,在残余草皮基部四周布满鼠洞。
黑土滩-杂类草次生裸地(人工草地建植前)[HZ(EXD)],研究区域位于青海省果洛藏族自治州大武县军马场,N 37°39.38′,E 101°20.17′,海拔3 344m;果洛州玛沁县大武乡大武河对岸,N 34°28′,E 100°12′,海拔3 751 m。研究区域概况:草地已退化为黑土型次生裸地,植物生长盛期盖度不足50%,且多为杂类草,植物群落均匀度差,有少量小嵩草、矮嵩草残留于黑斑、白斑周围,没有明显的优势植物,土壤疏松,鼠洞多,但罕见鼠类活动。
人工草地建植初期(AP):研究区域位于青海省果洛州大武县军马场,N 37°40.625′,E 101°21.083′,海拔3 437m;研究区域概况:总盖度达83%以上,其中禾本科盖度为61%左右,杂类草盖度不足14.5%,物种丰富度较低,一般不超过15种,草层生殖枝高度为50cm左右。
5年人工草地(APD):研究区域位于青海省大武县窝塞乡,N 34°22.003′,E 100°29.612′,海拔3 739m。研究区域概况:总盖度为68%左右,禾本科盖度15%,杂类草盖度达到50%左右,物种丰富度达到17种,7-8月形成以杂类草[马先蒿(Pedicularisspp.)等]为优势的群落。
14年人工草地(APRN):研究区域位于青海省大武县满掌乡,N 33°16′,E 100°28′,海拔3 946m。研究区域概况:草地类型为禾本科-杂类草型草地,草地盖度达到93%左右,草地具有较为明显的分层结构,禾本科优势度为36.4%,杂类草优势度为37.7%,莎草科植物多居草层下层。
1.3 草地载畜量
载畜量测定:产草量采用地上收获法[7],载畜量测定:根据中国北方草场资源调查办公室对中国北方11片牧区制定的绵羊单位标准,每个羊单位日食牧草鲜重4kg,暖季放牧时间为150d,牧草利用率为70%[21]。据此计算禾草-矮嵩草草甸(围栏禁牧草地、打草场草地或病畜产羔畜用地),理论载畜量为10.05羊单位/hm2;矮嵩草草甸(冬场),理论载畜量为8.35羊单位/hm2;小嵩草草甸(公共草场),理论载畜量为5.10羊单位/hm2;黑土滩-杂类草次生裸地(无法利用草场),理论载畜量为1.26羊单位/hm2。
1.4 采样及测定方法
采样时间为2009年7-8月。采样方法:地上植物采用样方法(样方面积为25cm×25cm),每样地6个重复;地下植物和土壤均采用土钻法(直径为5cm),以10cm为一层分层取样,共4层,每6钻为一个重复,每样地3~6个重复,地下植物采用水洗法,土壤、植物风干后粉碎,过0.25mm土壤分析筛,聚乙烯自封袋封存。有机碳含量测定采用日本岛津总有机碳分析仪(TOC-5000A)。有机碳储量采用同类草地平均值,计算方法见参考文献[9]。
1.5 数据处理
数据采用SPSS 18.0处理,采用单因素方差分析,显著系数为0.05。
2 结果与分析
2.1 高寒矮嵩草草甸退化演替系列植物-土壤有机碳储量分异特征
高寒矮嵩草草甸在放牧干扰下发生不同程度的退化,但不同界面有机碳储量变化明显不同步。即随退化演替的进行,地上植物有机碳储量降低,地下植物、土壤及土壤-植物系统有机碳储量先增高后降低。
地上植物有机碳储量最高点出现在禾草-矮嵩草草甸[(145.9±6.7)g/m2],同矮嵩草草甸[(131.8±9.6)g/m2]差异不显著,但显著高于其他演替阶段(P<0.05);最低值出现在黑土滩-杂类草次生裸地[(18.9±6.0 g/m2)]。
地下植物有机碳储量最高值出现在小嵩草草甸草毡表层开裂期[(3 011.6±114.3)g/m2],最低值出现在黑土滩-杂类草次生裸地[(121.6±14.9)g/m2],其中禾草-矮嵩草草甸、矮嵩草草甸、正常小嵩草草甸地下植物(0~40cm)有机碳储量处于(1 354.8±143.5)~(1 617.9±272.5)g/m2,三者差异不显著,但均显著低于小嵩草草甸草毡表层开裂期(P<0.05)(图1)。
土壤有机碳储量最高值出现在矮嵩草草甸[(16 805.94±856.5)g/m2],高于禾草-矮嵩草草甸[(14 023.1±289.5)g/m2],最低值出现在黑土滩-杂类草次生裸地,其有机碳储量比矮嵩草草甸低(8 255.152±671.5)g/m2。
土壤-植物系统碳储量最高值出现在矮嵩草草甸[(18 555.7±879.7)g/m2],同正常小嵩草草甸[(17 281.7±320.6)g/m2]和小嵩草草甸草毡表层开裂期[(17 587.9±626.2)g/m2]差异不显著,但显著高于禾草-矮嵩草草甸[(15 523.8±278.9)g/m2](P<0.05);最低值出现在黑土滩-杂类草次生裸地[(8 550.8±169.7)g/m2](图2)。
2.2 人工草地恢复演替系列植物-土壤有机碳储量分异特征
人工草地建植初期虽然能够明显的改善草地的景观特征,但对植物-土壤系统有机碳储量的增汇作用贡献不大,黑土滩-杂类草次生裸地(人工草地建植前),植物-土壤系统固碳能力为(8 550.8±169.7)g/m2,人工草地建植初期固碳能力为(8 797.6±113.8)g/m2,同建植前差异不显著(P>0.05);5年人工草地,植物-土壤系统固碳能力为(9 726.2±29.8)g/m2,与前两阶段差异显著,14年人工草地植物—土壤固碳能力达到(13 648.8±28.7)g/m2,显著高于前3个阶段(P<0.05)(图3)。在黑土滩-杂类草次生裸地——人工草地建植初期——5年人工草地变化过程中,草地地上植物群落发生了明显更替,由禾本科群落演替为杂类草群落,但植物总体固碳能力没有达到显著差异(P>0.05),人工草地建植14年后草地地上、地下及整体植物固碳能力依次是黑土滩-杂类草退化草地的3.3,2.7和2.9倍,是建植初期人工草地的2.4,1.9和2.0倍,差异均达到显著水平(P<0.05)(图4)。
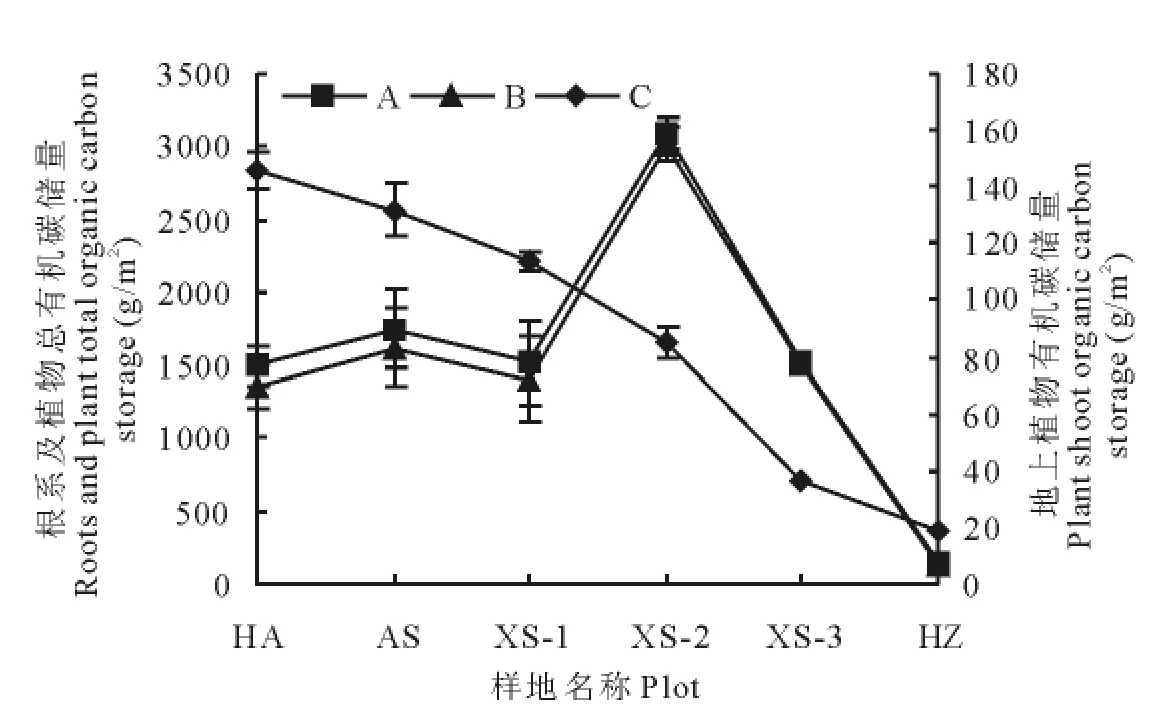
图1 植物有机碳储量特征Fig.1 Organic carbon storge in plant system
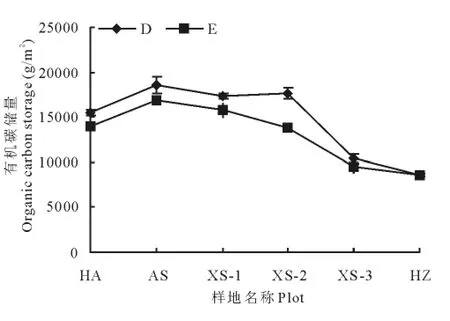
图2 土壤及植物-土壤有机碳储量特征Fig.2 Organic carbon storage in different succession storage
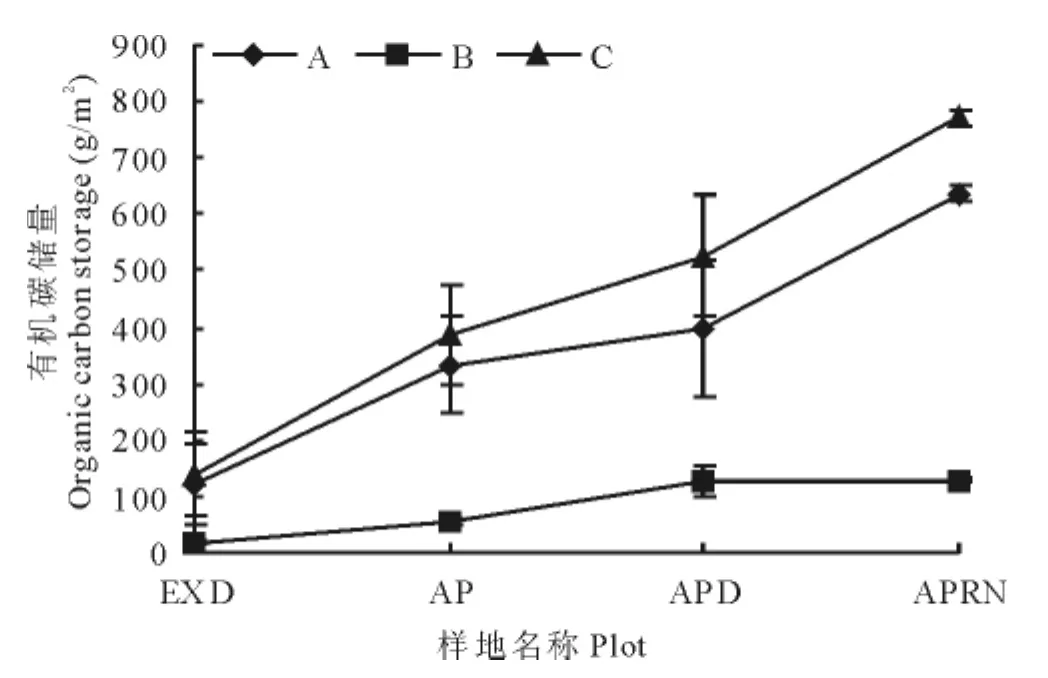
图3 人工草地不同演替阶段植物-土壤系统有机碳量特征Fig.3 Plant-soil system and soil system organic carbon storage in different plant succession stages
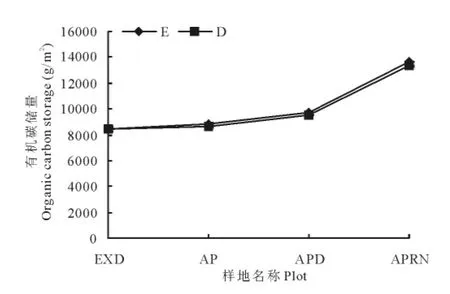
图4 人工草地不同演替阶段植物有机碳储量特征Fig.4 Plant organic carbon storage in different succession stages in artificial grassland
3 讨论
从草地的生产服务能力看,随草地利用强度加大,退化程度加深,草地的载畜能力逐渐减弱,其中禾草-矮嵩草草甸多为围栏封育草地,常作为牧民的病畜、产羔畜用地或冬季打草场,夏季很少利用,虽然该群落维持了较高的理论载畜量,但利用率低;矮嵩草草甸是继禾草-矮嵩草草甸后草地利用效率最高、经济生产服务能力最强的草场。黑土滩-杂类草次生裸地则在丧失了草地的生态服务能力的同时也失去了生产服务能力[7,9,21]。从草地碳储能力看,高寒矮嵩草草甸退化演替系列碳储能力先增加后降低,其最高值出现在矮嵩草草甸,但它与正常小嵩草草甸和小嵩草草甸草毡表层开裂期差异不显著,说明草地退化后改变了其碳储量在地上植物群落和根系中的分配比例,增加了土壤中有机碳的储量,使得草地植物-土壤系统对有机碳固定能力大幅度增加,但从小嵩草草甸草毡表层剥蚀期开始,土壤-植物系统有机碳储量开始降低,表明该系列小嵩草草甸草毡表层开裂期是该系列碳源/汇转换的拐点,草地开始由碳汇转变为碳源。但将黑土滩-杂类草次生裸地建植人工草地后,草地的碳储能力较建植前升高,草地由碳源转化为碳汇,且这种碳汇能力在一定时间范围内随草地建植年限增加而增加。从草地稳定性看,高寒矮嵩草草甸退化演替过程中,正常小嵩草草甸前,草地退化是由于过度放牧干扰引起,放牧干扰去除,草地可以在较短的时间内达到自我恢复,而从正常小嵩草草甸后,草地土壤养分供求关系、植物群落优势种生活型、野生动物群系(啮齿动物等)等内部因素发生改变[4,6-8],加之当地恶劣的自然条件(如冻溶交替),即使去除过度放牧干扰,草地仍然会发生退化,这种草地由于生态系统内因变化引起的退化,严重影响了草地的稳定性[4]。
综上所述,矮嵩草草甸不但可以维持较高的地上植物生产能力,还可以维持较高的植物-土壤系统的固碳能力,一旦草地放牧压力增加,草地植物群落即便发生明显变化,但植物-土壤系统的固碳能力并没有明显变弱,草地具有较高的稳定性,可以通过围栏禁牧或减牧在较短的时间内使其生态和生产能力得到改善,而正常小嵩草草甸成为高寒矮嵩草草甸牧压增大后的预警阶段[9]。
对已经极度退化的矮嵩草草甸(黑土滩-杂类草次生裸地)进行人工草地恢复后可以明显改善植物、土壤及植物-土壤系统的有机碳储能力,是一个较为明显的碳增汇过程。值得一提的是人工草地建植5年后,普遍会出现杂类草(马先蒿等)成为优势种的生产功能退化现象,目前针对该类退化人工草地采用除杂或重新翻耕等措施,一是反复的耕作措施会使草地越来越贫瘠[12];二是浪费了大量的人力和物力,人工草地建植5年左右仍然面临着生产功能退化的覆辙;三是灭杂采用的化学药剂对环境具有潜在的安全隐患。而人工草地退化后继续围栏禁牧,草地即会向原生植物群落演替[6],经过14年左右的封育,草地的群落结构已经由原来较为单一的禾本科优势功能群转变为具有一定分层结构,物种多样性较为丰富的群落类型,草地稳定性明显加强,草地的固碳能力不断的增高[11,12]。当然该类草地植物-土壤系统有机碳固定能力仍然显著低于原生群落[9],说明通过该草地群落有机碳固定能力较原生群落还存在较大的缺口,具有巨大的有机碳增汇潜力。
高寒矮嵩草草甸退化演替系列是一个伴随着载畜量降低,植物-土壤系统碳储能力先增高后降低的变化过程,草地演替系列中正常小嵩草草甸之前较为稳定,之后则稳定性明显降低,矮嵩草草甸是该退化演替系列中协调经济生产服务能力和碳容管理的适宜阶段,正常小嵩草草甸是该退化演替系列碳容管理的预警阶段。对于极度退化的黑土滩-杂类草次生裸地进行人工草地恢复,可以明显改善植物、土壤和植物-土壤系统有机碳储量、系统稳定性及草地载畜能力。人工草地建植5年后形式上的人工草地“退化”是草地生态系统自我调节的重要阶段,它并没有减少草地的碳储能力,而是由简单的人工草地群落向复杂稳定的原生群落转变的关键阶段,因此对黑土滩-杂类草次生裸地建植人工草地并围栏禁牧可以明显提高草地的生态和生产服务能力,这种管理方式是该类型退化草地的适宜碳容管理方式。
[1] Wang G X,Qian J,Cheng G D,etal.Soil organic carbon pool of grassland soils on the Qinghai-Tibetan Plateau and its global implication[J].The Science of the Total Environment,2002,291:207-217.
[2] Batjes N H.Management options for reducing CO2-concentrations in the atmosphere by increasing carbon sequestration in the soil[M].Wageningen:International Soil Reference and Information Centre,1999.
[3] Lal R.Soil carbon sequestration impacts on global climate change and foodsecurity[J].Science,2004,304:1623-1627.
[4] 曹广民,龙瑞军.三江源区“黑土滩”型退化草地自然恢复的瓶颈及解决途径[J].草地学报,2009,17(1):4-9.
[5] 李以康,林丽,张法伟,等.小嵩草群落——高寒草甸地带性植被压力下的偏途顶极群落[J].山地学报,2010,28(3):257-265.
[6] 曹广民,龙瑞军.放牧高寒嵩草草甸的稳定性及自我维持机制[J].中国农业气象,2009,30(4):553-559.
[7] 曹广民.青藏高原高寒草甸对放牧干扰的响应与适应[D].兰州:甘肃农业大学,2011.
[8] 曹广民,杜岩功,梁东营,等.高寒嵩草草甸的被动与主动退化分异特征及其发生机理[J].山地学报,2007,25(6):641-648.
[9] 林丽,曹广民,李以康,等.人类活动对青藏高原高寒矮嵩草草甸碳过程的影响[J].生态学报,2010,30(15):4012-4018.
[10] 马玉寿,张自和,董全民,等.恢复生态学在“黑土型”退化草地植被改建中的应用[J].甘肃农业大学学报,2007,4(2):91-97.
[11] 王文颖,王启基,王刚,等.高寒草甸土地退化及其恢复重建对植被碳、氮含量的影响[J].植物生态学报,2007,31(6):1073-1078.
[12] 王启基,李世雄,王文颖,等.江河源区高山嵩草(Kobresiapygmaea)草甸植物和土壤碳、氮储量对覆被变化的响应[J].生态学报,2008,28(3):885-894.
[13] 李月梅,王跃思,曹广民,等.开垦对高寒草甸土壤有机碳影响的初步研究[J].地理科学进展,2005,24(6):59-65.
[14] Grumbine R E.What is ecosystem management[J].Conservation Biology,1994,8(1):27-38.
[15] He N P,Yu Q,Wu L,etal.Carbon and nitrogen store and storage potential as affected by land-use inaLeymuschinensisgrassland of northern China[J].Soil Biology & Biochemistry,2008,40:2952-2959.
[16] Zhang C S,McGrath D.Geostatistical and GIS analyses on soil organic carbon concentrations in grassland of southeastern Ireland from two different periods[J].Geoderma,2004,119:61-275.
[17] 杨渺,李贤伟,张健,等.植被覆盖变化过程中土壤有机碳库动态及其影响因素研究进展[J].草业学报,2007,16(4):126-138.
[18] 张金霞,曹广民,周党卫,等.高寒矮嵩草草甸大气-土壤-植被-动物系统碳素储量及碳素循环[J].生态学报,2003,23(4):627-634.
[19] 任继周,梁天刚,林慧龙,等.草地对全球气候变化的响应及其碳汇潜势研究[J].草业学报,2011,20(2):1-22.
[20] 林丽,李以康,张法伟,等.青藏高原高寒矮嵩草草甸退化演替主成分分析[J].中国草地学报,2012,34(1):24-29.
[21] 许鹏.草地调查规划[M].北京:中国农业出版社,1999.
