FCER1B和STAT6基因单核苷酸多态性与小麦过敏的关联研究
2013-04-09蔡培培
蔡培培,尹 佳
(中国医学科学院 北京协和医学院 北京协和医院变态反应科 协和转化医学中心,北京 100730)
小麦是主要农作物之一,人类加工并消耗,在此过程中小麦过敏也随之出现[1]。根据对过敏原暴露的路径和潜在的免疫机制,小麦过敏可分为经典食物过敏(影响皮肤、胃肠道或呼吸道)、食物依赖运动诱发严重过敏反应(food-dependent exercise-induced anaphylaxis, FDEIA)、职业性哮喘即所谓的面包师哮喘(baker’s asthma,BA)及接触性荨麻疹[2]。瑞典的调查显示,小麦诱发的食物过敏在儿童的发生率为4%[2];美国的研究认为,小麦诱发的食物过敏在成人的发生率为0.4%[2]。针对韩国某大型面包工厂的问卷调查显示,BA的发生率为1.53%[3]。截至目前,小麦诱发的FDEIA和接触性荨麻疹的流行病学数据尚未见有研究发表,小麦诱发的FDEIA罕见,其发生率可能较低。小麦蛋白是小麦过敏的过敏原。小麦蛋白分为水溶性蛋白和非水溶性蛋白,水溶性蛋白包括清蛋白(albumins)和球蛋白(globulins);非水溶性蛋白包括醇溶蛋白(gliadins)和谷蛋白(glutenins),醇溶蛋白和谷蛋白联合称麦胶蛋白(gluten)[4]。小麦醇溶蛋白分为α-、β-、γ-、ω-醇溶蛋白,其中ω-醇溶蛋白被细分为慢-ω-醇溶蛋白(ω-1、ω-2)和快-ω-醇溶蛋白(ω-5)两种类型,ω-5-醇溶蛋白是导致小麦依赖运动诱发的严重过敏反应(wheat-dependent exercise-induced anaphylaxis,WDEIA)发生的主要致敏蛋白[1]。
小麦过敏是一种多因子疾病,即由遗传因素和环境因素相互作用而致其发生。目前,对多因子疾病的遗传因素研究一般通过遗传学关联研究实现。根据所研究的病种,选择具代表性的单核苷酸多态性(single nuclear polymorphism,SNP)位点,在遵循Hardy-Weinberg平衡定律的基础上研究SNP与多因子疾病的相关性。关联研究的经典模式是病例-对照研究,即通过比较等位基因频率或基因型频率在疾病组与对照组之间的统计学差异阐明基因多态性位点与表型的相关性[5]。
本研究采用SNP分型为基础的遗传学关联研究,对小麦过敏的遗传因素进行部分阐述。从小麦过敏的发生机制着手,确定2个细胞因子基因的SNP——FCER1B基因rs569108和信号转导和转录活化因子6(signal transducer and activator of transcription 6,STAT6)基因rs324015位点,分析这2个SNP的基因型频率及等位基因频率在病例组与对照组之间的统计学差异,探讨FCER1B基因rs569108和STAT6基因rs324015位点与小麦过敏的易感相关性。
对象和方法
分组及入组标准
本研究共设小麦过敏组、非小麦过敏组和健康对照组3组。
小麦过敏组入组标准:(1)患者食入小麦制品后发生食物过敏,或食入小麦制品4~6 h内运动发生WDEIA,或吸入面粉后发生BA;(2)小麦特异性IgE(specific IgE,sIgE)>0.35 kUA/L,或麦胶蛋白sIgE>0.35 kUA/L,或ω-5-醇溶蛋白sIgE>0.35 kUA/L。同时符合以上2个条件的患者可入组。
非小麦过敏组入组标准:(1)患有过敏性疾病,但与小麦不相关;(2)小麦sIgE<0.35 kUA/L,且麦胶蛋白sIgE<0.35 kUA/L,且ω-5-醇溶蛋白sIgE<0.35 kUA/L。同时符合以上2个条件的过敏患者可入组。
健康对照组入组标准:(1)无过敏性鼻炎;(2)无过敏性哮喘及咳嗽;(3)无皮肤过敏症;(4)无食物过敏;(5)小麦sIgE<0.35 kUA/L,且麦胶蛋白sIgE<0.35 kUA/L,且ω-5-醇溶蛋白sIgE<0.35 kUA/L。同时满足以上5条标准,即可入组。
对象
小麦过敏组:根据小麦过敏组入组标准的2个条件,对2004年3月至2010年3月就诊于北京协和医院的疑似小麦过敏的患者进行详细的病史和基本信息的采集,最终入组63例确诊小麦过敏患者,小麦过敏包括食物过敏、WDEIA及BA。63例小麦过敏组患者,首次发病的年龄范围为12~69岁,首次发病的平均年龄为(36.02±13.79)岁。其中,男31例,女32例,男女比例为0.97∶1。
非小麦过敏组:2004年3月至2010年3月就诊于北京协和医院的75例其他因素诱发过敏性疾病的患者,过敏性疾病包括其他食物诱发的食物过敏、花粉症、药物过敏等。75例非小麦过敏组患者,发病年龄范围为5~71岁,平均年龄为(31.93±13.88)岁。其中,男30例,女45例,男女比例为0.67∶1。
健康对照组:2010年3月至2010年8月北京协和医院体检中心197例非过敏健康人。年龄为21~77岁,平均年龄为(37.03±11.21)岁。其中,男92例,女105例,男女比例为0.88∶1。
方法
基于聚合酶链式反应-限制性片段长度多态性(Polymerase Chain Reaction-Restriction Fragment Length Polymorphism,PCR-RFLP)进行SNP分型。实验分为DNA提取及SNP分型两步骤进行。DNA提取:用全血基因组DNA提取试剂盒提取本实验所有样本DNA(人类全血基因组DNA提取试剂盒,大连宝生物工程有限公司)。SNP分型:PCR反应(PCR Mix,北京全式金生物技术有限公司);酶切(限制性内切酶,NEB);PCR扩增反应引物、扩增片段长度、限制性内切酶以及酶切条件见表1。
sIgE检测应用Phadia 250检测系统,ImmunoCAP检测试剂,ThermoFisher Scientific。
统计学分析
应用SPSS 16.0软件,行χ2检验分析SNP基因型在各对照组中的Hardy-Weinberg平衡;在遵循Hardy-Weinberg平衡定律的基础上,利用Fisher’s检验分析SNP的基因型频率及等位基因频率在小麦过敏组、非小麦过敏组与健康对照组之间的统计学差异,P<0.05为差异有统计学意义。
伦理审核
本研究已得到北京协和医院伦理委员会批准。
结 果
SNP分型
FCER1B基因rs569108多态位点的酶切图谱,纯合子CC 103 bp,纯合子TT 80 bp,杂合子CT 103、80 bp(图1)。STAT6基因rs324015多态位点的酶切图谱,纯合子AA 93 bp,纯合子GG 74 bp,杂合子AG 93、74 bp(图2)。
SNP在各组的基因型和等位基因频率分布
rs569108和rs324015位点的三种基因型和两种等位基因在各组的频率分布见表2。其中,rs569108位点的基因型CC在小麦过敏组中的统计值为0。
Hardy-Weinberg遗传平衡检验
FCER1B基因rs569108和STAT6基因rs324015位点的基因型频率在各组的分布均符合Hardy-Weinderg遗传平衡定律(P>0.05)(表3),说明所选择的样本具有群体代表性。

表1 PCR扩增及其酶切反应条件Table 1 Conditions of PCR and restriction enzyme digestion
PCR:聚合酶链式反应
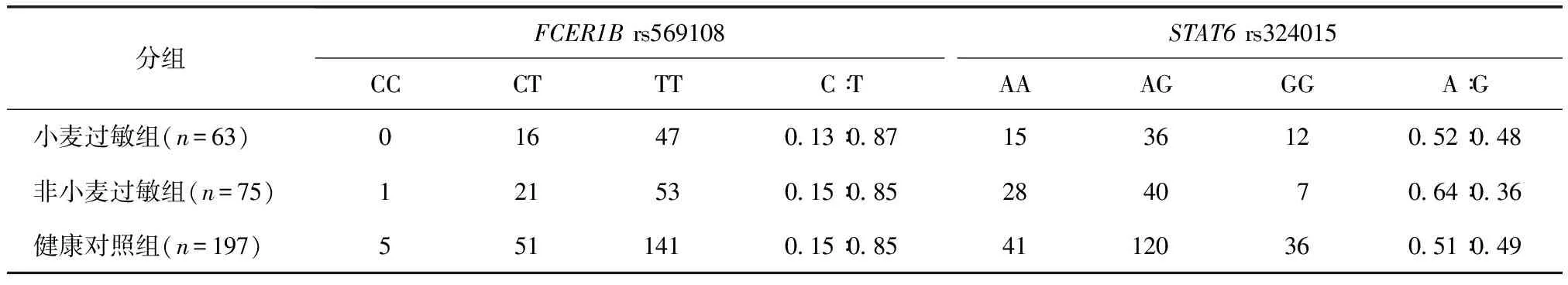
表2 FCER1B基因rs569108和STAT6基因rs324015位点的基因型和等位基因频率分布Table 2 Genotype frequencies distribution and allele frequencies distribution of loci of rs569108 in FCER1B gene and rs324015 in STAT6 gene
FCER1B 基因rs569108或STAT6基因rs324015与小麦过敏的关联性
差异统计结果显示,两个位点的基因型频率分布和等位基因频率分布在小麦过敏患者与非小麦过敏患者间比较差异无统计学意义(P>0.05)(表4),在小麦过敏患者与健康对照者间比较差异亦无统计学意义(P>0.05)(表5)。
FCER1B基因rs569108或STAT6基因rs324015与小麦过敏患者的sIgE水平关联性
对于FCER1B基因rs569108,因小麦过敏组无CC基因型,只比较CT基因型患者的sIgE水平与TT基因型患者sIgE水平间的差异,结果显示,两者差异无统计学意义(P>0.05)(图3)。
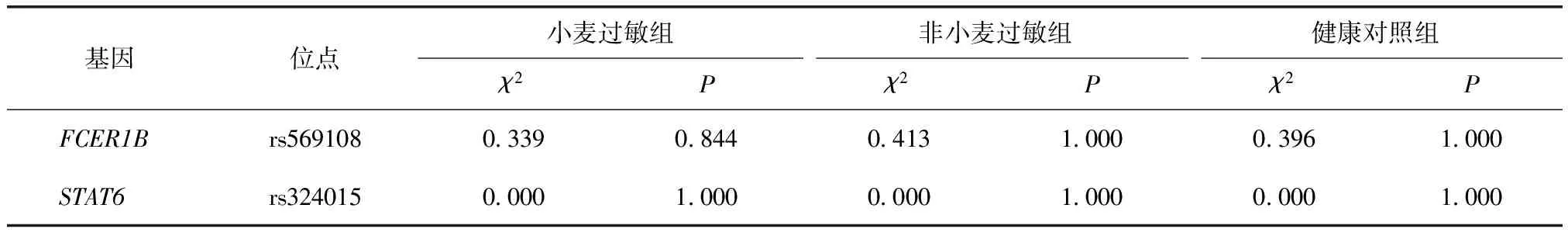
表3 FCER1B基因rs569108和STAT6基因rs324015位点在各组的Hardy-Weinberg遗传平衡检验Table 3 Hardy-Weinberg genetic equilibrium test of rs569108 in FCER1B gene and rs324015 in STAT6 gene in each group
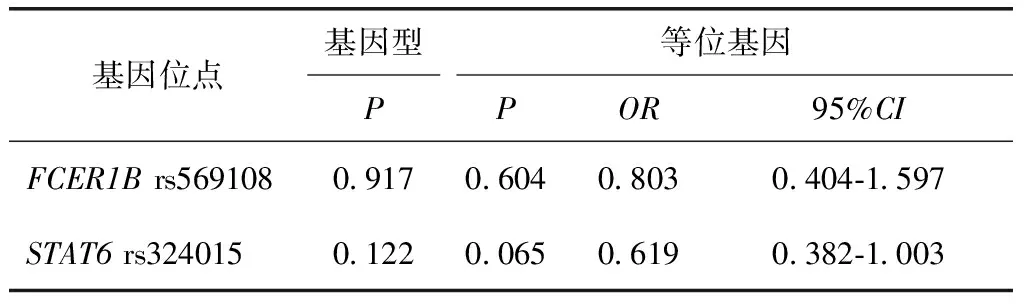
表4 FCER1B基因rs569108和STAT6基因rs324015位点的基因型频率及等位基因频率在小麦过敏患者组与非小麦过敏患者组之间的差异统计Table 4 Difference statistical results of the genotype frequencies and allele frequencies between patients with wheat allergy and patients with non-wheat allergyfor loci of rs569108 FCER1B gene and rs324015 in STAT6 gene
OR:比值比;CI:可信区间
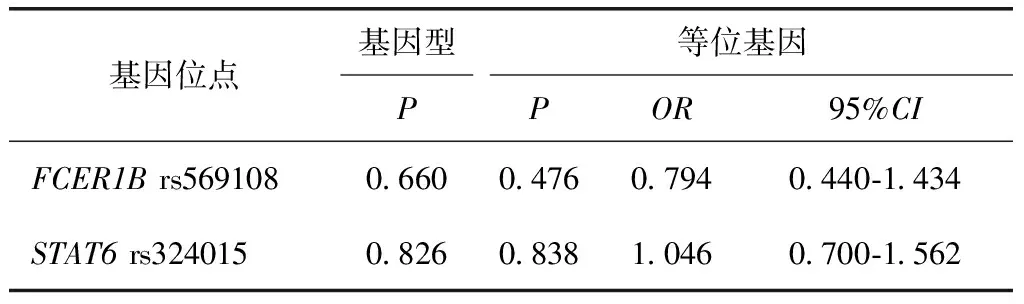
表5 FCER1B基因rs569108和STAT6基因rs324015位点的基因型频率及等位基因频率在小麦过敏患者组与健康对照组之间的差异统计Table 5 Difference statistical results of the genotype frequencies and allele frequencies between patients with wheat allergy and healthy controls for loci of rs569108 FCER1B gene and rs324015 in STAT6 gene
OR:比值比;CI:可信区间
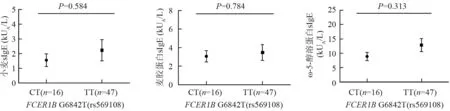
图3FCER1B基因rs569108多态位点CT基因型小麦过敏患者sIgE与TT基因型小麦过敏患者sIgE的比较
Fig3Comparison of sIgE levels between the group with CT and the group with TT for rs569108 polymorphic locus inFCER1Bgene
对于STAT6基因rs324015,比较次要等位基因纯合子AA患者的sIgE水平与含主要等位基因患者(AG+GG)sIgE水平间的差异,以及AG基因型患者sIgE水平与GG基因型患者sIgE水平间的差异,结果显示均无统计学意义(均P>0.05)(图4、5)。
讨 论
在以往过敏性疾病遗传学研究中,STAT6基因和FCER1B基因的SNPs被证明与多种过敏性疾病相关。
FCER1B基因位于11q12.1,全长10 kb,7个外显子,6个内含子。研究证明FCER1B基因多态性与过敏性疾病相关[6]。位于FCER1B基因启动子区域的rs569108位点与免疫受体酪氨酸活化基序位点相近,这个酪氨酸活化基序与γ链相互作用,最终影响IgE细胞信号通路效率[7]。也有研究指出,FCER1B基因的rs569108位点与rs1441585、rs574700位点紧密连锁,影响过敏性疾病的发展;其位点的等位基因C是日本儿童过敏性疾病的危险因素[8]。
STAT6是过敏反应诱导及调节的重要因子。STAT6定位于12q13.3-q14.1,全长19 kb,有23个外显子,22个内含子。位于3’UTR区内的SNP rs324015可影响mRNA的稳定性及蛋白质的表达量[9]。英国高加索人群中,STAT6 基因的rs324015位点与坚果过敏的易感性及严重性相关,且等位基因G、基因型GG是严重过敏反应的危险因素[10-11]。日本人群中,STAT6基因的rs324015位点与过敏性哮喘相关[12]。然而,在高加索人的同胞对研究中则显示STAT6基因的rs324015位点与哮喘无关联[13]。日本学者研究发现STAT6基因的rs324015位点与STAT6第一外显子13-GT突变存在连锁不平衡,联合评价这两个突变位点是预测过敏疾病的有效方法[14]。
虽然以往多项研究证明FCER1B基因rs569108位点和STAT6基因rs324015位点与过敏性疾病相关,但本研究中FCER1B基因SNP rs569108位点和STAT6基因SNP rs324015位点的基因型频率和等位基因频率在小麦过敏组与非小麦过敏组之间无统计学差异,在小麦过敏组与健康对照组之间亦无统计学差异。且FCER1B基因rs569108或STAT6基因rs324015与小麦过敏患者的sIgE水平无关联。由此提示FCER1B基因rs569108位点和STAT6基因rs324015位点与小麦过敏的易感性不相关。
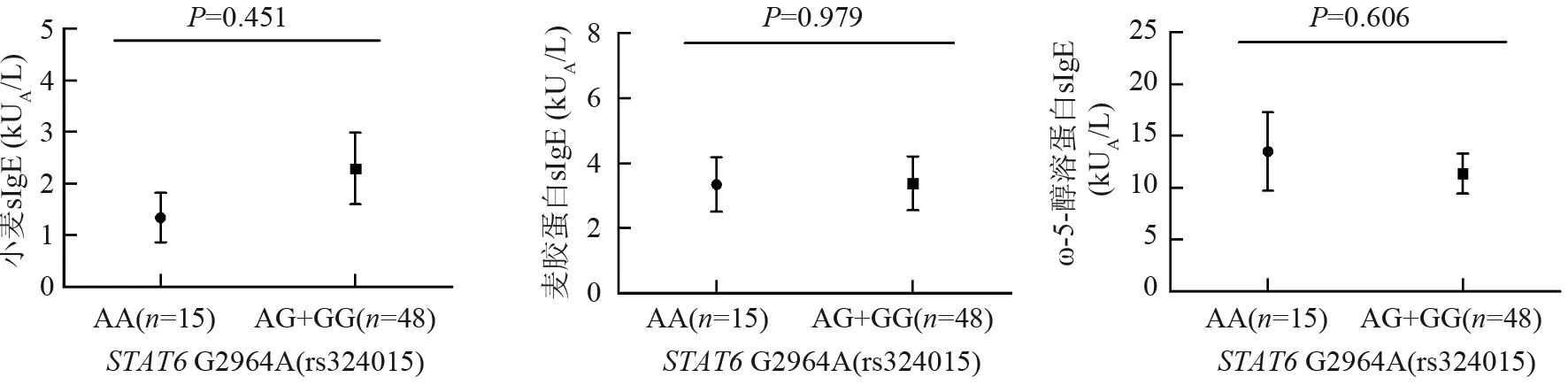
图4STAT6基因rs324015多态位点AA基因型小麦过敏患者sIgE与(AG+GG)基因型小麦过敏患者sIgE的比较
Fig4Comparison of sIgE levels between the group with AA and the group with (AG+GG)for rs324015 polymorphic locus inSTAT6 gene.
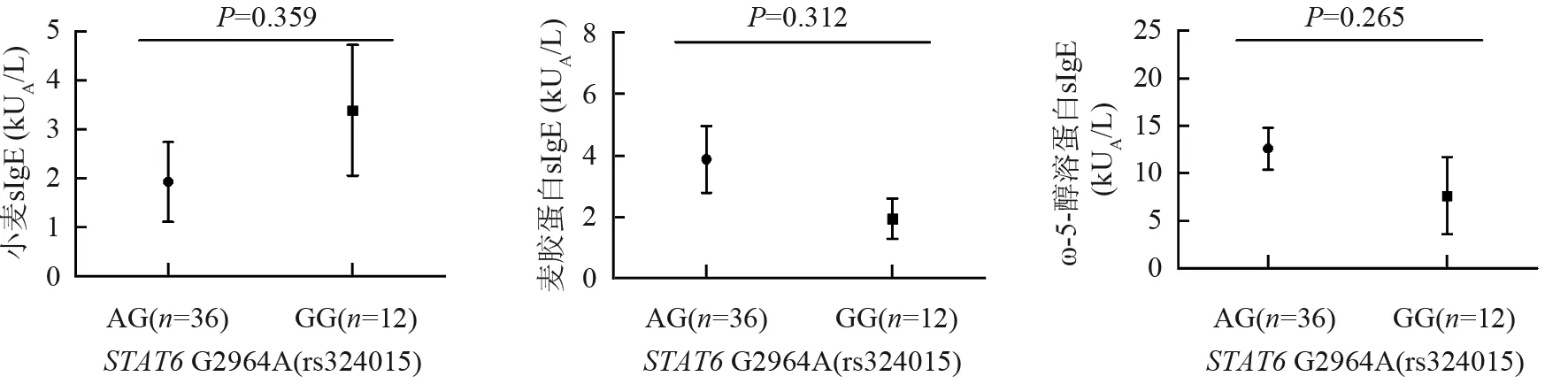
图5STAT6基因rs324015多态位点,AG基因型小麦过敏患者sIgE与GG基因型小麦过敏患者sIgE的比较
Fig5Comparison of sIgE levels between the group with AG and the group with GG for rs324015 polymorphic locus inSTAT6 gene.
本研究结果提示,rs569108和rs324015位点可能不参与小麦过敏的发生,这与以往诸多研究结果不一致。造成这种不一致的因素可能是:(1)基于小麦过敏的流行病学数据,本研究样本量小,未达统计学要求。(2)病例-对照组中研究对象存在各种因素的分层现象,本研究3个组别的研究对象平均年龄及年龄范围均不一致,可能存在年龄分层;本研究4个组别的男女比有差异,可能存在性别分层。因此,人群分层在基因多态性位点的关联研究中值得关注。(3)本 研究的研究对象是汉族,与既往相关研究对象的民族不同,造成遗传背景不同。(4)研究对象入选标准不一致,也会影响研究结果。
小麦过敏是一种多因子复杂疾病。进行基因-基因相互作用、基因-环境相关作用与小麦过敏相关性的研究,可能比研究单个SNP位点与复杂疾病的关联性更有效。同时,外显子组测序和全基因组测序也许是揭示复杂疾病的人类遗传学背景的更有效工具[15-16]。
(致谢:中国医学科学院基础医学研究所医学遗传系刘彦山为本实验提供技术支持)
(本文图1、2见插页第Ⅰ页)
[1]Tatham AS, Shewry PR. Allergens to wheat and related cereals[J]. Clin Exp Allergy, 2008, 38:1712-1726.
[2]Naoko Inomata. Wheat allergy[J]. Curr Opin Allergy Clin Immunol, 2009, 9:238-243.
[3]Gyu-Young Hur, Dong-Hee Koh, Hyoun-Ah Kim, et al. Prevalence of work-related symptoms and serum-specific antibodies to wheat flour in exposed workers in the bakery industry[J]. Res Med, 2008, 102:548-555.
[4]Shewry PR. Wheat[J]. J Exp Bot, 2009, 60:1537-1553.
[5]Lunetta KL. Genetic association studies[J]. Circulation, 2008, 118:96-101.
[6]Shirakawa T, Li A, Dubowitz M, et al. Association between atopy and variants of the beta subunit of the high-affinity immunoglobulin E receptor[J]. Nat Genet, 1994, 7:125-129.
[7]Nishiyama C, Akizawa Y, Nishiyama M, et al. Polymorphisms in theFCER1Bpromoter region affecting transcription activity: a possible promoter-dependent mechanism for association betweenFCER1Band Atopy[J]. J Immunol, 2004, 173:6458-6464.
[8]Thomas NS, Wilkinson J, Holgate ST. The candidate region approach to the genetics of asthma and allergy[J]. Am J Respir Crit Care Med, 1997, 156:144-151.
[9]Jacob CO, Lee Sk, Strassmann G. Mutational analysis of TNF-alpha gene reveals a regulatory role for the 3’untranslated region in the genetic predisposition to lupus-like autoimmune disease[J]. J Immunol, 1996, 156:3043-3050.
[10] Amoli M, Ollier WE, Hajeer AH. A novel PCR-RFLP assay for the detection of a polymorphism in the 3’ ofSTAT6 gene[J]. Genes Immun, 2000, 1:349-350.
[11] Amoli MM, Hand S, Hajeer AH, et al. Polymorphism in the STAT6 gene encodes risk for nut allergy[J]. Genes Immun, 2002, 3:220-224.
[12] Gao PS, Mao XQ, Roberts MH, et al. Variants ofSTAT6 (signal transducer and activator of transcription 6) in atopic asthma[J]. J Med Genet, 2000, 37:380-382.
[13] Duetsch G IT, and Wjst M.STAT6 as an asthma candidate gene: polymorphism-screening, association and haplotype analysis in a Caucasian sib-pair study[J]. Hum Mol Genet, 2002, 11:613-621.
[14] Tamura K, Suzuki M, Arakawa H, et al. Linkage and association studies ofSTAT6 gene polymorphisms and allergic diseases[J]. Int Arch Allergy Immunol, 2003, 131:33-38.
[15] Bamshad MJ, Ng SB, Bigham AW, et al. Exome sequencing as a tool for Mendelian disease gene discovery[J]. Nat Rev Genet, 2011, 12:745-755.
[16] Cirulli ET, Goldstein DB. Uncovering the roles of rare variants in common disease through whole-genome sequencing[J]. Nat Rev Genet, 2010, 11:415-425.
