杜仲MEP途径系列基因5′端非编码区的顺式作用元件预测
2013-04-03乌云塔娜刘慧敏杜红岩
乌云塔娜,刘慧敏,杜红岩
(1.中国林业科学研究院 经济林研究开发中心,河南 郑州 450003;2.国家林业局杜仲工程技术研究中心,河南郑州 450003;3.中南林业科技大学 a.经济林育种与栽培国家林业局重点实验室;b.林学院,湖南 长沙 410004)
杜仲Eucommia ulmoidesOlive是我国特有的经济树种,属国家二级保护植物,其树叶、树皮、果实均含丰富的杜仲胶,杜仲皮为中药上品,也是我国传统的出口创汇商品之一[1-4]。杜仲胶在国际上被称为古塔波胶,其化学结构式为反式-聚异戊二烯(C5H8)n,是普通天然橡胶巴西橡胶的同分异构体[5]。杜仲胶高弹性体具有“橡胶-塑料”双重特性和形状记忆等特殊功能[6],能广泛应用于交通、通讯、医疗、电力、国防、水利、建筑和人们的日常生活中,其产业化前景十分广阔[7-9]。
植物中用于不同萜类化合物合成的前体物异戊烯基二磷酸(isopentenyl diphosphate,IPP)和二甲烯丙基二磷酸(dimethylallyl diphosphate,DMAPP)在质体中主要是由2-C-甲基-D-赤藓糖醇 -4-磷 酸(2-C-Methyl-D-Erythritol-4-Phosphate,MEP)途径合成的[10-12]。该途径的终产物IPP是许多萜类物质(包括杜仲胶)的前体物,该途径涉及到多种酶的参与,许多基因受到多基因编码和转录水平的调节。基因中的5′段非编码区存在着与转录相关的序列,包括启动子、增强子、应答软件等顺式作用元件及一些常见的组件TATA盒、CAAT盒等调节组件,是调控翻译的一个重要组成部分。目前无论是在人、动物还是在植物、真菌中均有关于5′端非编码区的研究。2012年高震[13]对小麦属和山羊草属Waxy基因5′端非编码区序列的变异进行了分析,采用了PCR克隆、聚类分析、系统进化分析等方法深入了解了小麦属和山羊草属Waxy基因5′端的变异,为进一步了解Waxy基因的功能和调节机制,以及利用小麦属和山羊草属基因资源奠定基础。2013年Wenjing Wu[14]等人通过将一段2.0 kb的5′非编码区的片段克隆到荧光素酶报告基因载体上,缺失和突变分析表明Ets-2结合位点是尿路上皮癌抗原1(UCA1)基因启动子或活性的关键。Keunsub Lee[15]等对利用转录组序列土壤农杆菌A.tumefaciens C58进行转录起始位点定位分析发现其5′端非编码区的长度超过60 nt,表明有大量的顺式作用元件被编码在土壤农杆菌的基因组中。
1 材料与方法
1.1 序列来源
本研究所用数据资料来自杜仲胶合成时期的转录组数据和杜仲全基因组测序数据。
1.2 分析方法
利 用 PLANTCARE(http://bioinformatics.psb.ugent.be/webtools/plantcare/html/)在线软件预测MEP途径系列基因5′端非编码区中的顺式作用元件,用PLACE(http://www.dna.affrc.go.jp/PLACE/signalscan.html)在线软件做辅助分析。
2 结果与分析
2.1 杜仲MEP途径EuDXS基因5′端非编码区顺式作用元件结构和功能预测
将1-脱氧-D-木酮糖-5-磷酸合成酶基因(EuDXS2)的5′端非编码区序列提交到在线软件PLANTCARE和PLACE中,预测EuDXS2基因5′端可能存在的顺式作用元件。结果表明:EuDXS2基因5′端非编码区共有22种共220个顺式作用元件,不仅包括启动子顺式元件TATA-box、CAAT-box等,还包括光应答、热胁迫、干旱诱导、类黄酮生物合成、胚乳表达、糖抑制、真菌诱导、乙烯应答等多种元件;EuDXS2基因5′端非编码区包含TATA-box可能的位点有145个,占总元件的65.91%;CAAT-box可能的位点有39个,占总元件的17.73%(见表1)。
2.2 杜仲MEP途径EuDXR基因5′端非编码区顺式作用元件结构功能预测
将1-脱氧-D-木酮糖-5-磷酸还原异构酶基因(EuDXR)的5′端非编码区序列提交到在线软件PLANTCARE和PLACE中,预测EuDXR基因5′端可能存在的顺式作用元件。结果表明:EuDXR基因5′端非编码区共有21种共127个顺式作用元件,包括CAAT-box、TATA-box等元件,还涉及到光应答、厌氧感应、乙烯响应、赤霉素响应、热胁迫、干旱诱导、胚乳表达、水杨酸响应等多种方面。顺式元件中包含27个CAAT-box,占总元件的21.26%;68个TATA-box,占总元件的53.54%(见表2)。
2.3 杜仲MEP途径EuCMK 基因5′端非编码区顺式作用元件结构功能预测
将4-(5′-焦磷酸胞苷)-2-C-甲基 -D-赤藓醇激酶基因(EuCMK)的5′端非编码区序列提交到在线软件PLANTCARE和PLACE中,预测EuCMK基因5′端可能存在的顺式作用元件。结果表明:EuCMK基因5′端非编码区共有24种共134个顺式作用元件,包括CAAT-box和TATA-box等顺势元件,此外还涉及到光应答、胚乳表达、热胁迫、防御和应激、乙烯响应、干旱诱导、茎特异表达、昼夜调节等多种方面。预测出32个CAAT-box,占总元件的23.88%;68个TATA-box,占总元件的50.75%(见表3)。
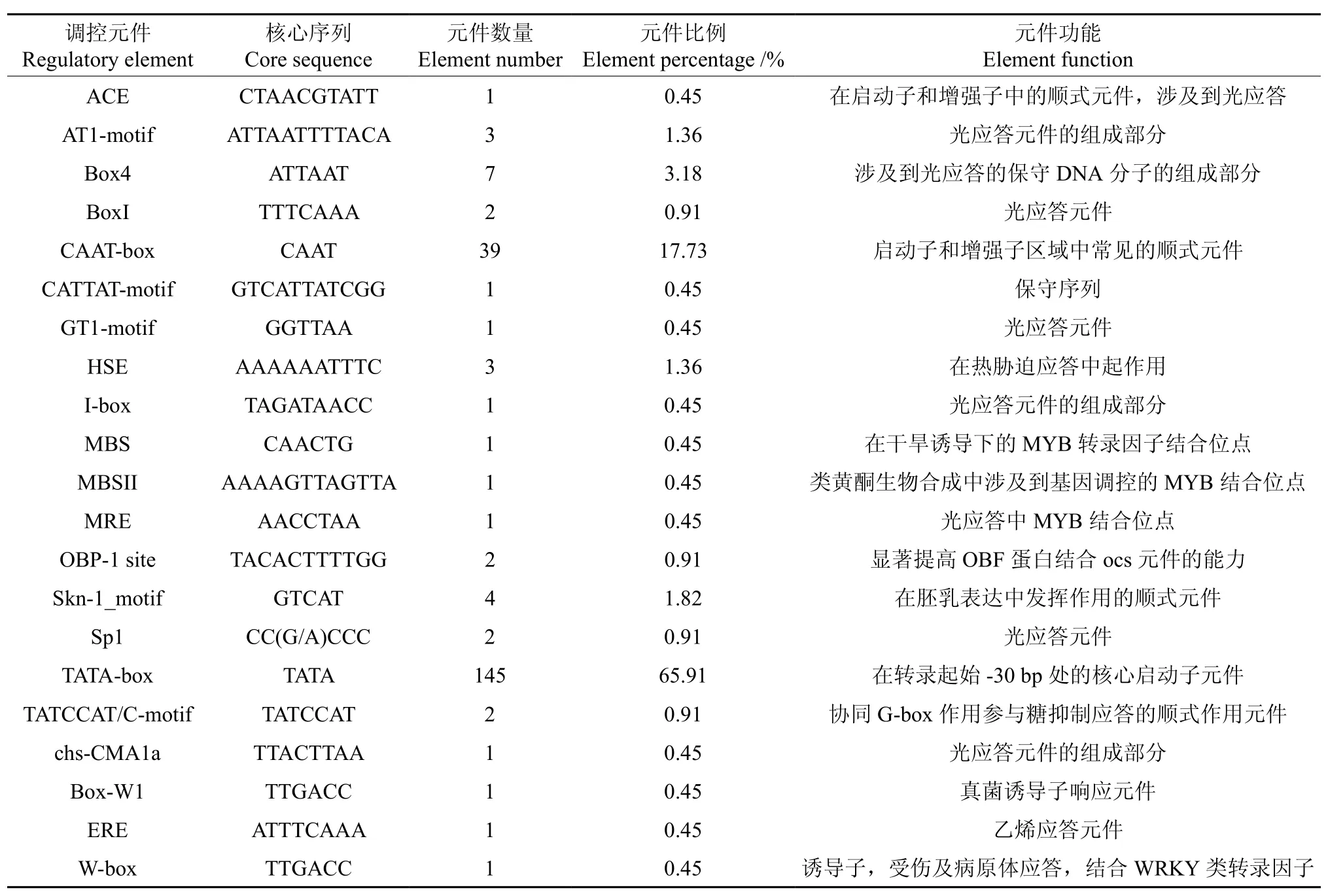
表1 EuDXS2基因5′端顺式作用元件分布Table 1 Distribution of the cis-acting elements in non-coding region of EuDXS2 5′end

表2 EuDXS2基因5′端顺式作用元件分布Table 2 Distribution of the cis-acting elements in non-coding region of EuDXR 5′end
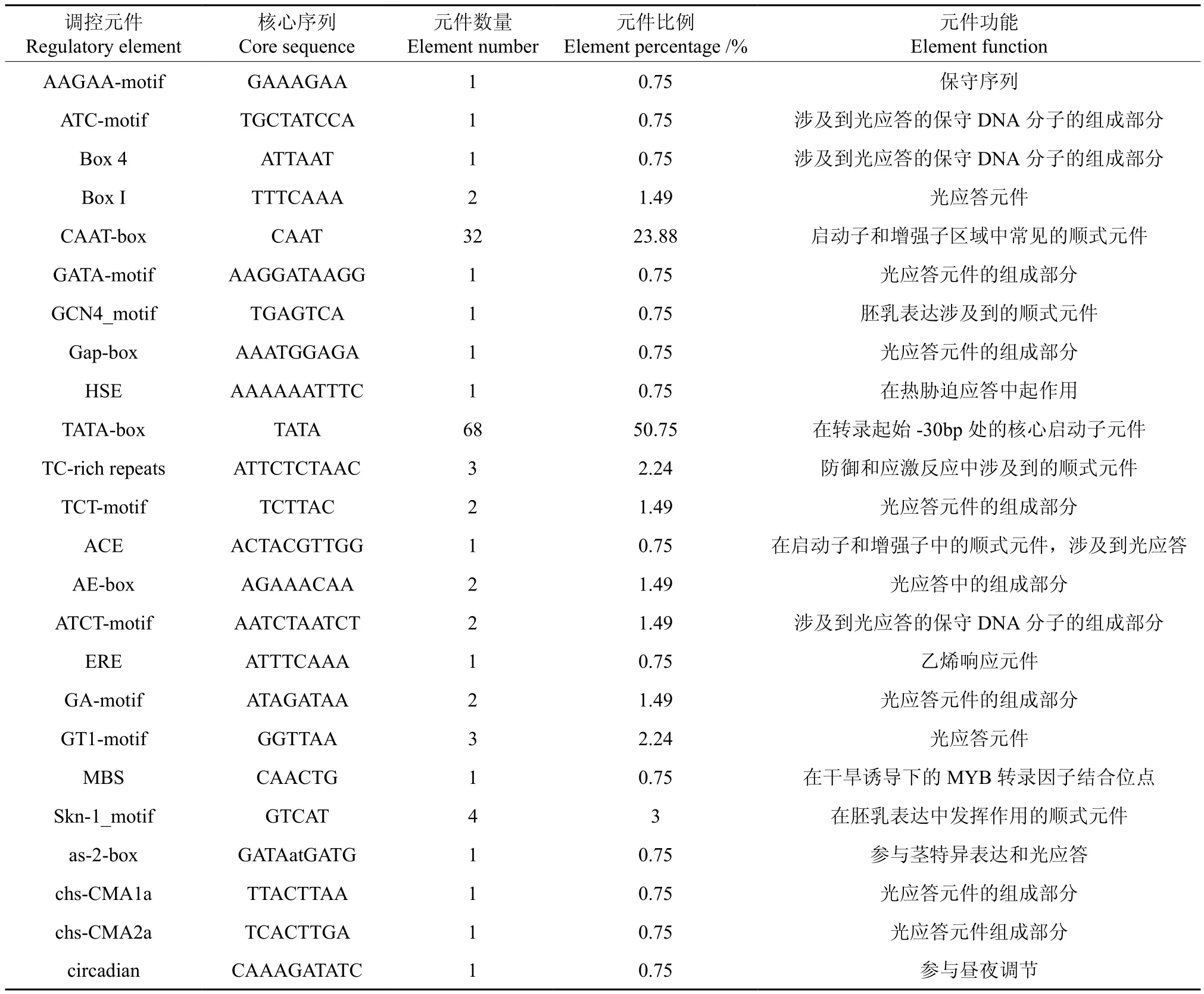
表3 EuCMK基因5′端顺式作用元件分布Table 3 Distribution of the cis-acting elements in non-coding region of EuCMK 5′end
2.4 杜仲MEP途径EuMDS基因5′端非编码区顺式作用元件分析
将2-甲基-D-赤藓醇-2,4-环焦磷酸合酶基因(EuMDS)的5′端非编码区序列提交到在线软件PLANTCARE和PLACE中,预测EuMDS基因5′端可能存在的顺式作用元件。结果表明:EuMDS基因5′端非编码区共有30种共104个顺式作用元件,包括CAAT-box和TATA-box等顺式元件,此外还涉及到光应答、脱落酸响应、厌氧感应、真菌诱导、分生组织、胚乳表达、热胁迫、低温响应、糖抑制、昼夜调节等多种方面。预测出43个CAAT-box,占总元件的41.35%;23个TATA-box,占总元件的23.08%(见表4)。
2.5 杜仲MEP途径EuHDS基因5′端非编码区顺式作用元件分析
将1-羟基-2-甲基-2-E-丁烯基-4-焦磷酸合酶基因(EuHDS)的5′端非编码区序列提交到在线软件PLANTCARE和PLACE中,预测EuHDS基因5′端可能存在的顺式作用元件。结果表明:EuHDS基因5′端非编码区共有35种共126个顺式作用元件,包括TATA-box和CAAT-box等顺式元件,此外还涉及到光应答、脱落酸响应、木质部表达、厌氧感应、真菌诱导、茉莉酸响应、缺氧反应、热胁迫、胚乳表达、分生组织活性、防御和应激、昼夜调节等多种方面。预测出29个CAAT-box,占总元件的23.02%;34个TATA-box,占总元件的26.98%(见表5)。
2.6 杜仲MEP途径EuHDR基因5′端非编码区顺式作用元件分析
将1-羟基-2-甲基-2-E-丁烯基-4-焦磷酸还原酶基因(EuHDR)的5′端非编码区序列提交到在线软件PLANTCARE和PLACE中,预测EuHDR基因5′端可能存在的顺式作用元件。结果表明:EuHDR基因5′端非编码区共有24种共115个顺式作用元件,包括TATA-box和CAAT-box等顺式元件,此外还涉及到光应答、真菌诱导、分生组织活性、茉莉酸响应、胚乳表达、热胁迫、干旱诱导、防御和应激反应、水杨酸响应、生长素响应等多种方面。预测出19个CAAT-box,占总元件的16.52%;67个TATA-box,占总元件的58.26%(见表6)。
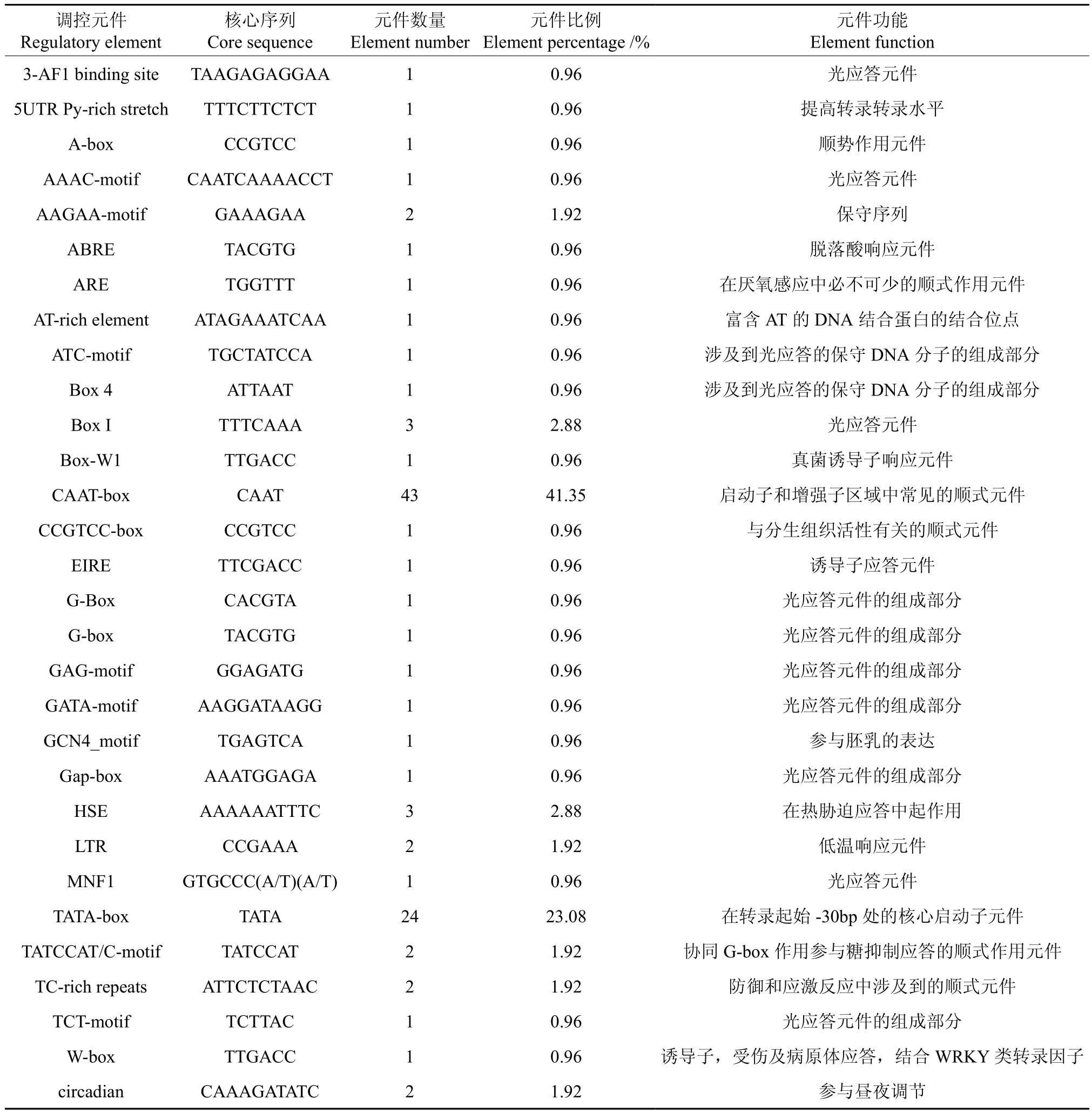
表4 EuMDS基因5′端顺式作用元件分布Table 4 Distribution of the cis-acting elements in non-coding region of EuMDS 5′end
3 结论与讨论
以杜仲转录组数据和基因组数据为基础,预测了杜仲MEP途径6条酶基因5′端非编码区中可能存在的顺式作用元件,为从分子水平上进一步研究MEP生物合成途径的调控提供了理论基础。EuDXS是MEP途径的第1个关键酶,EuDXS2基因5′端非编码区共有22种共220个顺式作用元件,其中有145个TATA-box,占总元件的65.91%,39个CAAT-box,占总元件的17.73%;EuDXR是MEP途径的第2个限速酶,EuDXR基因5′端非编码区共有21种共127个顺式作用元件,其中有27个CAAT-box,占总元件的21.26%;68个TATA-box,占总元件的53.54%;EuCMK催化MEP途径的羧基磷酸化反应,EuCMK基因5′端非编码区共有24种共134个顺式作用元件,其中有32个CAAT-box,占总元件的23.88%;68个TATA-box,占总元件的50.75%;EuMDS催化MEP途径的第5步酶促反应,EuMDS基因5′端非编码
区共有30种共104个顺式作用元件,其中有43个CAAT-box,占总元件的41.35%;23个TATA-box,占总元件的23.08%;EuHDS为MEP途径的第6个作用酶,EuHDS基因5′端非编码区共有35种共126个顺式作用元件,其中有29个CAAT-box,占总元件的23.02%;34个TATA-box,占总元件的26.98%;EuHDR是MEP途径的第3个限速酶,EuHDR基因5′端非编码区共有24种共115个顺式作用元件,其中有19个CAAT-box,占总元件的16.52%;67个TATA-box,占总元件的58.26%。此结果为用生物信息学软件参考其它植物的序列预测的数据,5′端非编码区的结构及功能,尤其是启动子部分CAAT-box和TATA-box的具体位置及功能还需要在此理论的指导下通过试验验证。

表5 EuHDS基因5′端顺式作用元件分布Table 5 Distribution of the cis-acting elements in non-coding region of EuHDS 5′end
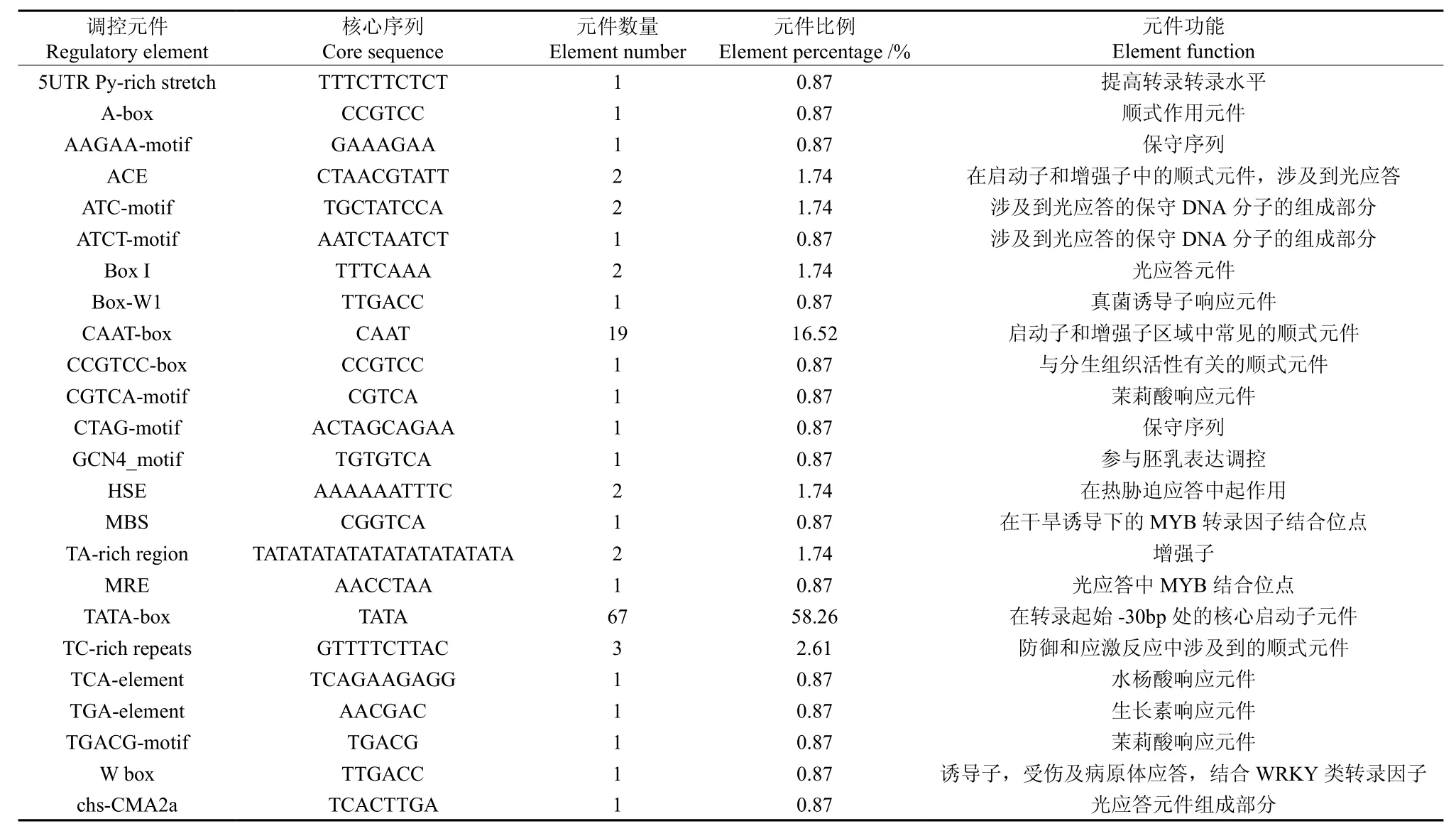
表6 EuHDR基因5′端顺式作用元件分布Table 6 Distribution of the cis-acting elements in non-coding region of EuHDR 5′end
目前,MEP途径酶基因5′端的生物信息学研究鲜见报道,大部分研究停留在转录组水平。2009年李辉亮[16]等人克隆了巴西橡胶树4-羟基-3-甲基-2-(E)-丁烯基-4-磷酸还原酶基因(HbHDR)并对其表达做了分析,得到的HbHDR长1 627 bp,编码462个氨基酸,属于LYTB家族,半定量RT-PCR结果显示,乙烯诱导胶乳HbHDR的表达。2007年马靓[17]克隆了烟草5-磷酸脱氧木酮糖还原异构酶(DXR)基因全长cDNA并对其序列做了分析,克隆得到的基因全长为1 804 bp,共编码473个氨基酸,同源序列比对发现DXR氨基酸序列和本赛姆式烟草DXR的一致性达到97%,进化树分析显示烟草DXR与番茄DXR的亲缘关系最近,且基本反应了10种不同植物DXR之间的进化关系。
文中对MEP途径系列基因5′端非编码区的顺式作用元件进行了预测,为进一步研究MEP生物合成途径中相关基因的启动子克隆、启动子结构和功能以及MEP途径产物的分子调控提供了理论依据。
参考文献:
[1] 杜红岩,胡文臻,俞 锐.杜仲产业绿皮书:中国杜仲橡胶资源与产业发展报告[M].北京:社会科学文献出版社,2013: 1-5.
[2] 薛 萍.杜仲研究现状与发展前景[J].经济林研究,1995,13(3):56-57.
[3] 王 征,金晓玲,刘雪梅,等.杜仲成熟胚器官发生途径的研究[J].中南林业科技大学学报, 2013, 33(6): 79-80.
[4] 李铁柱,杜红岩,刘慧敏,等.杜仲果实和叶片转录组数据组装及基因功能注释[J].中南林业科技大学学报, 2012, 32(11):122-123.
[5] 叶生晶,乌云塔娜,田大伦,等.杜仲MVA途径相关酶基因的鉴定及荧光定量PCR引物筛选[J].中南林业科技大学学报,2013,33(8):50-51.
[6] 杜红岩.我国的杜仲胶资源及其开发潜力与产业发展思路[J].经济林研究,2010,28(3):1-2.
[7] 杜红岩,刘昌勇,李 钦,等.杜仲叶中3种主要活性成分含量的季节变化[J].中南林业科技大学学报,2011, 31(8):6-7.
[8] 李铁柱,杜红岩,刘慧敏,等.杜仲幼果和成熟果实转录组数据组装及基因功能注释[J].中南林业科技大学学报,2012,32(10): 9-10.
[9] 严瑞芳.杜仲胶研究进展及发展前景[J].化学进展,1995, (7):65-71.
[10] 张松涛,陈红丽,崔 红,等.植物MEP途径的代谢调控机制[J].西北植物学报,2012,32(7):1500-1504.
[11] 李 恒.萜类化合物MEP生物合成途径中关键酶1-脱氧-D-木酮糖-5-磷酸合酶和1-脱氧-D木酮糖-5-磷酸还原异构化酶的研究[D].西安:西北大学,2012:5-6.
[12] 李 恒.萜类化合物MEP生物合成途径中dxr基因的克隆表达及酶活性分析[D].西安:西北大学,2009:8-9.
[13] 高 震.小麦属和山羊草属Waxy基因5′端非编码区序列变异分析[D].雅安:四川农业大学,2012:1-2.
[14] Wenjing Wu, Shuwan Zhang, Xu Li, et al.Ets-2 Regulates Cell Apoptosis via the Akt Pathway, through the Regulation of Urothelial Cancer Associated1, a Long Non-Coding RNA, in Bladder Cancer Cells[J].PLOS ONE,2013,8(9):1-2.
[15] Keunsub Lee, Xiaoqiu Huang, Chichun Yang, et al.A Genome-Wide Survey of Highly Expressed Non-Coding RNAs and Biological Validation of Selected Candidates in Agrobacterium tumefaciens[J].PLOS ONE,2013,8(8):1-2.
[16] 李辉亮,雷美玉,彭世清.巴西橡胶树4-羟基-3-甲基-2-(E)-丁烯基-4-磷酸还原酶基因(HbHDR)的克隆及表达分析[J].基因组学与应用生物学,2009,28(1):15-16.
[17] 马 靓.烟草5-磷酸脱氧木酮糖还原异构酶(DXR)基因全长cDNA的克隆及序列分析[D].武汉:华中科技大学,2007.
