草原毛虫研究现状与展望
2013-03-14张棋麟袁明龙
张棋麟,袁明龙
(草地农业生态系统国家重点实验室 兰州大学草地农业科技学院,甘肃 兰州730020)
草原毛虫别名红头黑毛虫、草原毒蛾,隶属鳞翅目(Lepidoptera)毒蛾科(Lymantriidae)草原毛虫属(Gynaephora)。草原毛虫在国外没有危害和防治的报道,仅将其作为一种对极端生态环境适应的模式物种进行研究。而在我国,草原毛虫是严重危害青藏高原高寒草地的主要害虫。该虫主要取食莎草科和禾本科优良牧草,造成家畜食物短缺,改变草地植物群落结构,加剧草地退化和草地生态环境恶化。草原毛虫不仅分布范围广(超过200万hm2)、发生密度高(通常200~500头·m-2,最高虫口密度达3 000头·m-2)[1],而且还会导致家畜中毒[2-4],严重阻碍青藏高原畜牧业的健康发展。2003年,仅青海省草原毛虫的发生危害面积就超过100万hm2,直接经济损失达9 500万元之多。自20世纪60年代以来,国家每年都要投入大量的人力、物力和财力防治草原毛虫,但是还不能从根本上有效地控制该害虫的危害。特别是近年来,由于全球气候变化,该虫在青藏高原高寒草地频繁爆发成灾,危害进一步加重。尽管草原毛虫具有重要的经济意义,但目前对草原毛虫基础和应用研究还非常有限。本研究从草原毛虫种类、生物学与生态学特性、预测预报及防治等方面总结草原毛虫的研究现状,并讨论其今后应重点开展的研究方向。
1 草原毛虫的种类
草原毛虫属是毒蛾科的一个小属,由Hübner于1822年建立,属的模式种为白斑草原毛虫(G.selenitica)。中国草原毛虫属昆虫的采集和研究最早是由外国人于19世纪末对黄斑草原毛虫(G.alpherakii)的描述开始的[5]。周尧和印象初[6-7]对我国草原毛虫属4个新种进行了报道,之后严林等[6-7]对青海及周边地区的草原毛虫种类进行了细致、深入的调查和研究,报道了青海地区草原毛虫属3个新种。迄今为止,全世界文献记载的草原毛虫属昆虫共15种[8],主要分布于北半球的高山以及北极的冻土地带,尤以高原地区居多。亚洲分布13个种,欧洲分布3个种,北美1个种,北极2个种,其中亚洲草原毛虫种类集中分布在青藏高原(8个种)和帕米尔高原(4个种)。我国草原毛虫属的8个种,全部分布于青藏高原,是青藏高原的特有昆虫。其中,青海草原毛虫(G.qinghaiensis)和门源草原毛虫(G.menyuanensis)的分布范围最广,前者在青海、西藏、甘肃和四川等地从海拔3 000到5 000 m均有分布,后者分布于海拔3 000 m左右的青藏高原东北部的青海省和甘肃省;其他草原毛虫均为局部分布,如曲麻莱草原毛虫(G.qumalaiensis)主要分布于海拔4 500 m左右的青海曲麻莱和治多,久治草原毛虫(G.jiuzhiensis)主要分布于4 000 m左右的青海久治,而若尔盖草原毛虫(G.rouergensis)和小草原毛虫(G.minora)仅分布于海拔3 500 m左右的四川若尔盖草原[8]。草原毛虫的种类和分布见表1。
2 草原毛虫生物生态学特性研究现状
2.1草原毛虫的生活史 草原毛虫属全变态类昆虫,有卵、幼虫、蛹和成虫4个发育阶段。门源草原毛虫一年发生一代,以休眠的1龄幼虫在雌茧内、草根下或土中越冬[9]。门源草原毛虫雄幼虫有6个虫龄,雌幼虫有7个虫龄。以1龄幼虫形态过冬,翌年4月中旬牧草返青时,1龄幼虫开始取食,大多数幼虫到4月下旬后就进入2龄期,经过15 d左右又进入3龄期,到了4龄以后就发育缓慢,一个龄期须经20多天,到了7月下旬开始结茧化蛹,一直持续到10月初基本结束,8月中旬是化蛹盛期,从这一时期到10月上旬为羽化、交配和产卵,10月中旬1龄幼虫孵化,接着以滞育形态越冬[8]。

表1 世界上草原毛虫的种类及分布Table 1 Species and distribution of Gynaephora around world
2.2性二型、性比和成虫寿命 昆虫纲中雌雄二型现象广泛存在,至少有23个目的昆虫具有该特征,有翅亚纲的昆虫有很多种类发生性二型现象,尤其以完全变态类的鳞翅目昆虫较为普遍[10]。同种草原毛虫之间的雌、雄个体除生殖器官结构不同以外,在颜色、大小等方面也存在显著差异。例如,门源草原毛虫的雌成虫无翅、触角也不明显,而雄成虫有翅、触角呈羽毛状。在幼虫生活史方面,门源草原毛虫的雄性幼虫比雌性幼虫少发育1个虫龄(雄虫6龄,雌虫7龄),雄虫缩短发育时间有利于提高成活率,而雌虫利用多活的几十天,可以繁殖更多的后代,但由于生活时间的延长而导致死亡风险也相应增加,因此,前一优势是对后一劣势的弥补[8]。此外,雌雄蛹的颜色、大小及形态也表现出性二型。由于雄蛹要发育翅芽及飞行附属器官,所以雄蛹的发育时间较雌蛹的长,但雄蛹比雌蛹轻、小,颜色也比雌蛹浅。
门源草原毛虫雌雄虫性别变化同虫口密度变化相关,在密度低时,雌雄比为1∶0.64,而高密度时为1∶1.10[8]。该草原毛虫在种群密度低时,雌性占大多数,这样可以使繁殖率保持在较高水平;种群密度高时,雌虫相比雄虫要少,由于雌虫体型大,消耗能量多,这样就可以避免种群能量在高密度情况下产生不必要的损失。可见,草原毛虫种群的性比会随着种群密度不断变化,这样有利于能量趋向于保持在合理支出的水平,这也是该虫生长发育适应环境的一个表现。
门源草原毛虫雌成虫寿命(20~53 d)比雄成虫(3~8 d)平均长30 d左右,雌成虫寿命长使毛虫繁殖高峰期延长,把消耗的能量尽可能多地用在繁殖子代而不是自身发育上[8]。总之,性二型使草原毛虫提高了能量利用率,最大限度地把能量投入到生殖活动,提高了种群的适合度。
2.3草原毛虫的取食选择与生殖活动 草原毛虫在国外主要发生在北极冻土带,每年有11个月处在休眠期[11]。因此,关于草原毛虫对寄主植物的选择性,国内外的研究均较少。草原毛虫食性不但受遗传因素和虫口密度影响,还受栖息地食物种类、食物数量、食物组成、食物中各种营养含量和比例、毛虫性别以及虫龄等影响[12-13]。例如,北极洲和北美地区的格陵兰草原毛虫幼虫喜食桦木(Betulanana)、鹿蹄草(Pyrolagrandiflora)、北极高山柳(Salixspp.)、委陵菜(Potentillauniflora)和多瓣木(Dryasintegrifolia)等[14]。严林等[15]采用野外扣笼法发现,门源草原毛虫可取食44种植物,喜食的达到16种,喜好程度依次为二柱头藨草(Scirpusdistigmaticus)、小嵩草(Kobresiapygmaea)、垂穗披碱草(Elymusnutans)、羊茅(Festucaovina)、黄花棘豆(Oxytropisochrantha)等。而严林等[16]、万秀莲和张卫国[17]采用抽样调查法发现,青海草原毛虫喜食的植物种类仅为5种,但可食的达20余种,其中对莎草科植物最偏好,其次为禾本科,最后为杂草;喜食度依次为嵩草(Kobresiaspp.)、垂穗披碱草、剪股颖(Agrostishugoniana)、苔草(Carexkansuensis)和珠芽蓼(Polygonumviviparum)。草原毛虫不仅取食食谱很复杂,而且对于其喜食植物,还存在相应的喜食部位。有关草原毛虫取食选择的研究大多认为,植物的茎尖、叶缘和叶端由于质嫩、新鲜、易于消化等原因,是草食昆虫的最佳选择。此外,可能还存在其它因素影响草原毛虫的取食部位,例如光照因素,至于哪些因素起主导作用还是几种因素间相互协调影响等,尚需进一步研究[17]。
2.4温湿度对草原毛虫生长发育影响的研究 有关温度对草原毛虫生长发育影响的研究较少。Kukal等[18]报道了格陵兰草原毛虫的行为热调节。Kukal和Dawson[19]研究了温度对高龄幼虫取食、同化和生长率的影响。严林[8]通过设置恒温、恒温交替和室内自然变温3种条件研究门源草原毛虫幼虫发育对温度的反应,结果表明,恒温条件下,门源草原毛虫在10~33 ℃范围中,发育期随温度增加而变短,发育率上升,而在亚适温区,发育期和发育率则相反,超过40 ℃则全部死亡;恒温交替条件下,门源草原毛虫发育率和死亡率随变温的均温增加而上升,且发育适温区范围比恒温时更广,为6~39 ℃;室内自然变温下,可以用“S”型曲线描述门源草原毛虫幼虫发育速率,对同一个温度来说,自然变温条件下该虫发育速率相比恒温时要低[8]。门源草原毛虫幼虫发育起点温度为3.8 ℃,有效积温为1 462 ℃·d,41 ℃为致死温度,20 ℃为最适温[8]。有关草原毛虫对低温适应能力的研究主要集中于草原毛虫在北极洲和北美高寒、高山地区适应寒冷气候的宏观生态机制。如Kevan等[20]报道了在北极圈,罗丝草原毛虫幼虫通过朝向阳光生活且在迎风处隐蔽,利用表皮获得热量,使体温比周围空气温度高13 ℃,表现出该虫分布很大程度上受温度影响。Kukal等[18]以格陵兰草原毛虫为研究对象,观察到该幼虫活动期50%以上时间都在晒太阳,体温超过周围环境温度约20 ℃,能充分利用阳光辐射来生长和发育。生活史对策也是草原毛虫低温适应能力的一方面,Convey[21]研究发现,生活在北极的草原毛虫大多是A/S生活史对策,即表现出较长生活史、晚成熟、低发育率、低繁殖率、扩散能力弱和易变的种群密度[8]。Bennett等[22]还发现格陵兰草原毛虫在小生境中过冬喜选在背风、向光、温暖、潮湿的岩石附近,而不是植物丰富处。总之,在长期的进化过程中,草原毛虫在体型、行为以及生活史对策等方面均表现出对严寒极端生态环境的高度适应性[23]。
湿度对草原毛虫幼虫生长发育影响的研究也很少。草原毛虫喜湿,虫源一定时,降水量和相对湿度对毛虫有重要影响。例如,青海省河南县1991年到1995年降水量从55.8 mm升到235.0 mm,湿度也从60%增加到68%,青海草原毛虫,虫口密度就上升了近3倍,但是如果湿度过大,会造成毛虫卵腐烂[24]。草原毛虫幼虫对湿度的趋性比成虫更大,因为幼虫总在高湿、阳光充足的草地上聚集。另外,刮风、下雨气候潮湿时,成虫几乎不进行生殖活动;晴天中午、阳光直射且空气中水分含量相对较低时,毛虫很少交配;阴天时空气中湿度相对适量的时候,毛虫交配程度最强[25]。青藏高原是世界关注的生态脆弱和敏感区域,正是草原毛虫生殖活动能力的强盛,才使得草原毛虫种群密度、发生期、发生空间等产生很强的波动性和不确定性,引起大发生、大流行[26]。所以,分析和了解草原毛虫生境的温湿度范围,对于有效抑制、消灭草原毛虫具有重要的参考价值。
2.5草原毛虫的空间分布格局 对草原毛虫空间分布的研究,主要以草原毛虫的幼虫和蛹为研究对象,利用Taylor式S2=axb(式中,S2为方差,x为均数)和Iwao回归法进行聚集指标测定[27-28]。万秀莲和张卫国[17]报道了在样区尺度的同一水平面上,黄斑草原毛虫大致呈现出均匀分布,而在研究尺度上则是聚集分布,在垂直空间上,10~20和30~40 cm这两个高度范围内有明显的黄斑草原毛虫集中分布,表明黄斑草原毛虫分布总体呈聚集型,单个个体的分布也是如此[29]。可见,草原毛虫在幼虫期聚集分布明显,这样便于该种群成规模的取食,从而提高取食效率,在与其他食草动物的取食竞争中获胜,导致虫灾发生。在成虫期聚集分布的条件下,草原毛虫可以获得更多的交配机会,提高交配成功率,为翌年虫害的发生打下基础。
3 草原毛虫的防治现状研究
3.1预测预报 草地虫害预测,是科学估计草地虫害发生、蔓延和危害的根据,是治理虫害的一项重要工作,也是职能部门做出科学决策的根本依据。草原毛虫危害具有区域性,且迁移能力有限,因而对草原毛虫的预测预报,主要做本地虫源估计,异地虫量预测为次,通常不考虑迁入和迁出量。草原毛虫的预测预报最早开展于20世纪80年代,但已有的研究报道比较少,主要是根据上一年草原毛虫的基本个体数量、虫口密度、性比、死亡率、气候因素和死亡原因等来对翌年毛虫的数量进行预测[30]。其中,性比和个体数量主要调查的是上一年草原毛虫的雌成虫数量和繁殖的后代数目[31],而虫口密度可根据严林和兰景华[32]使用过的Nachman模型预测。对于最终发生数量的预测,可以用虫体基数、性比、繁殖后代数和成活率乘积来表示[30]。当然,预报某个地区虫害情况时,要首先明确草原毛虫的种类、发生区域[33]以便准确有效地开展后续工作。侯秀敏和徐秀霞[34]指出,预测预报要加强长期、系统的定位观测工作,减少监测盲区,不断改进测报手段,目的是获取连续有效、覆盖全面、科学精准的观测数据,能准确掌握虫害消长规律和发生趋势。在秋末冬初结合草原毛虫产卵量和气象部门的长期预报进行预测和综合评价,也可以在7―8月初根据草原毛虫的化蛹率、气象部门预测和取食植物生长情况,作出翌年1龄幼虫的发生趋势预测[31]。同时,要多方面协调发展,使预测预报逐步科学化、标准化,以提高对草原毛虫短、中、长期预测预报工作的时效性和准确性。除此之外,虫害的发生与同期的气象、水文条件是紧密联系的,因此预测预报应结合当地气象资料、水文条件等进行[35],提高预测预报的准确性。
3.2防治现状 鉴于草原毛虫具有周期性、聚集性和毒性的危害特点,草原毛虫的防治难度巨大[36]。随着人类对草原毛虫生境的影响,草原毛虫对青藏高原高寒草地的适应性也在不断增强[37]。目前,对草原毛虫的防治,特别是当草原毛虫大面积、高密度发生时,仍主要依赖于化学防治。
1)化学防治。对草原毛虫的化学防治应推行“时间一致,用药统一”的防治思路,并在3龄期之前进行,同时要注意降低对其天敌的毒害。对草原毛虫具有良好防治效果的农药有杀灭灵[38-39]、辛硫磷[40-41]、敌百虫[42]、敌敌畏[43]、敌杀死[44],但这类有机磷农药,对家畜毒性高,目前使用较少。阿维菌素、氯氰菊脂[45-46]、有机菊酯[47-48]及精油类[49]等,这类多为抗生素、植物杀虫剂,对家畜较为安全,目前使用较为普遍。但随着生态草原、绿色肉奶工程的实施,对草原毛虫的可持续控制应坚持“绿色植保、生态植保”之理念。
2)生物防治。草原毛虫的天敌类群有寄生蜂、寄生蝇、鼠类、鸟类、寄生性细菌和寄生性病毒等[50-52]。草原毛虫病原中主要是细菌(表2)[53-54],其中研究较多的是苏云金芽孢杆菌(Bacillusthuringiensis)和类产碱假单胞菌(Alcaligensspp.)[55-56]。另外,对草原毛虫病原微生物做过专门报道的主要是从罹死草原毛虫体内分离出的草原毛虫核型多角体病毒(Gynaephora qinghaiensis Nuclear Polyhedrosis Virus,GqNPV)[53-57]。该病毒具有安全性,对人、家畜无致病性和致突变性[58-59],对草原毛虫最佳致死浓度为1.25×108PIB·mL-1,致死中时(LT50)为5.8 d,最适致死温度为20~25 ℃[60-61]。以此为基础开发的草原毛虫病毒杀虫剂是一种安全、经济、有效的生物防治手段[62]。刘世贵[63-64]在总结前人经验后提出了“综合性利用生物防治”的策略,并利用其发现的草原毛虫病毒与杆状芽胞菌相结合,研制出了更高级的病毒杀虫剂,在川青藏高原20多万hm2草地上防虫效果明显。
至今报道过的草原毛虫寄生昆虫有7种(表3),其中优势种是草原毛虫金小蜂(Pteromalusquinghaiensis)和多刺孔寄蝇(Spoggosiaechinura)两种[65],前者主要寄生草原毛虫的蛹,寄生率达33%[66],而后者对门源草原毛虫的寄生率为23.5%[65]。鸟类和西藏鼠(Ochotonathibetaea)和长尾仓鼠(Cricetuluslongiacudatus)等鼠类以草原毛虫的幼虫和蛹为食,也是草原毛虫的一种重要天敌,对毛虫数量有一定的抑制作用[9,25]。总体而言,目前对草原毛虫的生物防治研究仍停留在天敌种类的描述和寄生率调查上,尽管初步明确了草原毛虫的优势天敌种类及寄生行为特征,但在生产中采用天敌防治草原毛虫的案例还非常少。
3)其它防治方法。可以把人工防治和物理防治结合,草原毛虫成虫对阳光趋性很强,在夏秋季节阳光强烈的中午前后,集中人力对成虫进行人工捕杀;同时要注意在草原毛虫蛹期捡拾毛虫蛹,并集中焚烧和掩埋;草返青前宜用火烧虫卵,能有效抑制翌年草原毛虫种群数量[9]。
4 问题与展望
目前,对草原毛虫的研究集中在分类、生物学、生态学及防治等[1,8,67-68]方面,但基础研究仍非常薄弱,远远滞后于生产中防治之需,这与其重要的经济和生态地位极不对称。特别是对草原毛虫属的种间系统发育关系、物种形成过程、高海拔适应机制以及频繁爆发成灾的遗传学信息知之甚少。因此,今后应重点开展以下研究。
4.1草原毛虫的快速鉴定 DNA条形码(DNA Barcoding)最初由加拿大动物学家Hebert等[69]于2003年提出,是利用线粒体DNA细胞色素C氧化酶亚基I(cytochrome c oxidase subunit I,COI)的5’-端长约650 bp的标准基因片段,在DNA水平上进行物种鉴定。目前,基于线粒体COI基因的DNA条形码技术作为一种新的物种鉴定手段,由于其快速、准确、简便的操作流程而被广泛应用于众多生物类群的物种鉴定,包括昆虫[70-71]、鸟类[71-74]、鱼类[75-76]和线虫类[77]等。鳞翅目是昆虫中应用DNA条形码最多的,鳞翅目DNA条形码协会(All leps barcodes of life),网站为http://www.lepbarcoding.org,目前已收录了74 503个鳞翅目物种的近634 870个条形码序列。
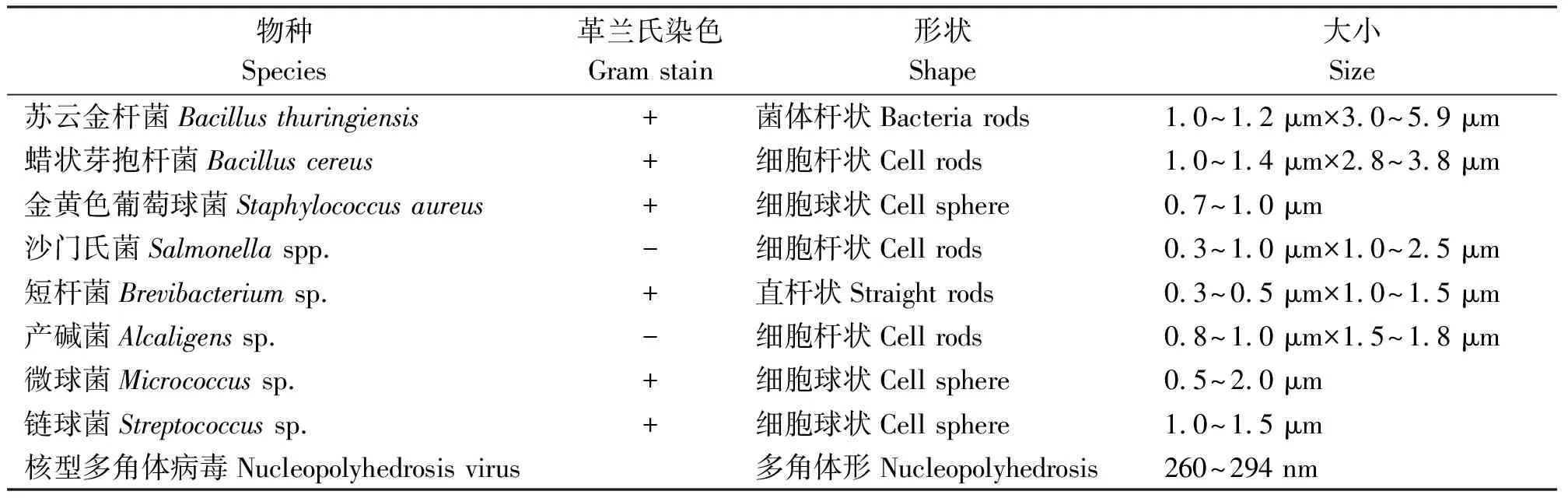
表2 草原毛虫主要病原微生物的种类和主要特征Table 2 Species and characteristics of main pathogenic microorganisms of Gynaephora
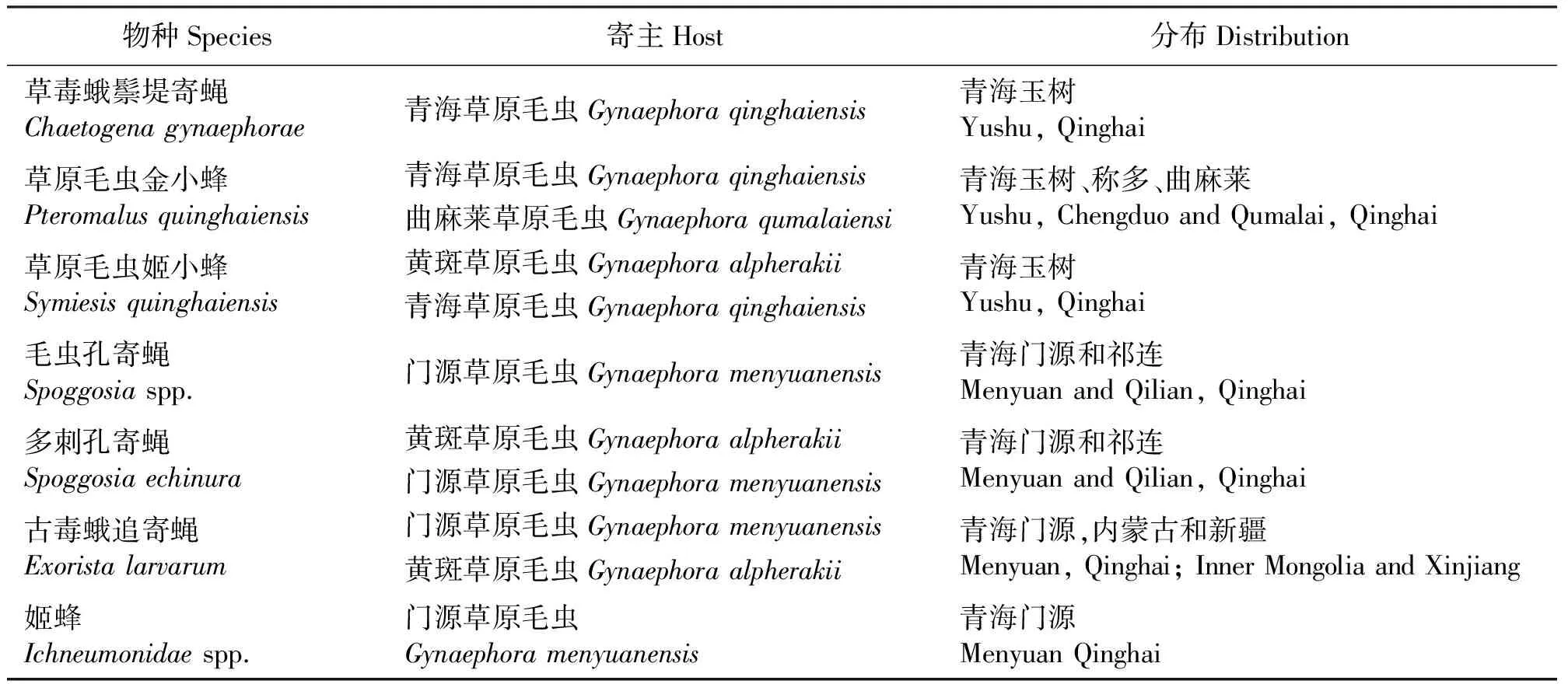
表3 草原毛虫天敌昆虫的种类及分布Table 3 Species and distribution of natural enemy insects of Gynaephora
青藏高原动植物区系中一个突出特点就是特有种占有很大比例,是物种形成的中心。我国草原毛虫属的8个种全部分布于青藏高原,占世界草原毛虫种的一半以上(8/15),因而青藏高原是名符其实的草原毛虫物种形成中心。尽管严林[8]对我国草原毛虫属8种昆虫的研究资料进行了系统总结,并建立了草原毛虫属成虫和幼虫的物种检索表,但采用外部形态特征对草原毛虫进行鉴定仍较困难,对鉴别人员的技术要求也非常高,尤其对卵、低龄幼虫和蛹。因此,有必要建立基于线粒体COI基因的草原毛虫DNA条形码技术,实现对草原毛虫不同虫态的快速而准确的鉴定。
4.2草原毛虫谱系地理学研究 草原毛虫能够在青藏高原缺氧、严寒这种极端生态环境中长期生存,并成为高寒草地的优势类群,必定具有适应于栖息地环境的种群遗传学和谱系地理结构特征,这也是剖析草原毛虫生态适应机制及爆发成灾规律的关键所在。此外,开展青藏高原特有昆虫草原毛虫的种群遗传结构及谱系地理学研究,也有助于了解在全球气候变化的背景下,青藏高原土著物种对全球气候变化的响应机制。因此,有必要研究草原毛虫的种群遗传多样性,追溯种群历史动态,揭示不同地理种群间基因流动的模式,探讨草原毛虫种间和种内的遗传分化及生态进化规律,解析青藏高原地质历史事件及气候环境的变迁对草原毛虫现今种群遗传结构及分布格局的影响,推断草原毛虫第四纪冰期时可能的避难所及冰期后的迁移扩散路线。研究结果不仅可以丰富青藏高原土著物种,特别是土著昆虫的谱系地理学研究,充实生物进化及遗传变异的研究内容,而且在实践上亦为从分子水平上了解草原毛虫的生态适应机制和成灾规律以及制定有效的控制策略提供理论依据。
4.3草原毛虫对青藏高原高海拔的生态适应机制研究 目前,高原缺氧、严寒环境导致的基因组适应性进化,在部分青藏高原土著动物中已有报道,但不同的物种对高原极端生态环境适应所涉及的基因不尽相同,且涉及的物种仅限于人类和哺乳动物[78-83]。迄今,还未见青藏高原特有昆虫等节肢动物基因组对高海拔生态环境的适应性进化研究。草原毛虫是我国青藏高原的特有种,也是严重危害高寒草地的重大害虫,该虫频繁爆发成灾,显示出其对青藏高原高寒、低氧生态环境良好的适应性。因此,草原毛虫为研究青藏高原土著物种对高海拔的适应性进化机制,提供了一个良好的模式系统。但是,目前还不清楚草原毛虫对高海拔适应性的遗传学机制,而这却是剖析草原毛虫生态适应机制及爆发成灾机制的重要内容。因此,有必要开展草原毛虫关键基因对高海拔的适应性进化,开展草原毛虫转录组学研究,旨在从分子水平上揭示草原毛虫对青藏高原的高海拔适应机制。
[1] 杨爱莲.西藏青海部分地区草原毛虫为害严重[J].草业科学,2002,19(5):73.
[2] 刘文清,河生德,尼玛.不同条件下草原毛虫引起放牧家畜口膜炎的危害性调查[J].草业与畜牧,2011(3):58-61.
[3] 尼玛,河生德,李长云.草原毛虫引起牦牛口膜炎的防治效果观察[J].草业与畜牧,2011(4):47-48.
[4] 尖措.河南县2004年草原毛虫防治效果显著[J].青海草业,2004(4):55-11.
[5] 周尧,印象初.草原毛虫的分类研究[J].昆虫分类学报,1979,1(1):23-28.
[6] 严林,刘振魁,梅洁人,等.青海草原毛虫种类的调查研究[J].青海畜牧兽医学院学报,1994,24(1):26-28.
[7] 刘振魁,严林,霍科科,等.青海三种草原毛虫染色体的观察[J].昆虫知识,1998(2):101.
[8] 严林.草原毛虫属的分类、地理分布及门源草原毛虫生活史对策的研究[D].兰州:兰州大学,2006:1-53.
[9] 魏学红.西藏草原毛虫的发生及防治对策[J].草原与草坪,2004(2):56-57.
[10] 王孟卿,杨定.昆虫的雌雄二型现象[J].昆虫知识,2005,42(6):721-725.
[11] Morewood W,Lange P.Immature stages of high arcticGynaephoraspecies(Lymantriidae) and notes on their biology at Alexandra Fiord,Ellesmere Island,Canada[J].Journal of Research on the Lepidoptera,1997,34:119-141.
[12] Floater G.Rainfall,nitrogen and host plant condition:consequences for the processionary caterpillar,Ochrogasterlunifer[J].Ecological Entomology,1997,22(3):247-255.
[13] 朱麟,古德祥.昆虫对植物次生性物质的适应策略[J].生态学杂志,2000,19(3):36-45.
[14] Maclean S F,Jensen T S.Food plant selection by insect herbivores in Alaskan arctic tundra:the role of plant life form[J].Oikos,1985,44(1):211-221.
[15] 严林,刘振魁,梅洁人,等.野外扣笼条件下草原毛虫对食物的选择[J].草地学报,1995(4):257-268.
[16] 严林,江小蕾,王刚.门源草原毛虫幼虫发育特性的研究[J].草业学报,2005,14(2):116-120.
[17] 万秀莲,张卫国.草原毛虫幼虫的食性及其空间格局[J].草地学报,2006(1):84-88.
[18] Kukal O,Heinrich B,Duman J G.Behavioural thermoregulation in the freeze-tolerant arctic caterpillar,Gynaephoragroenlandica[J].Journal of Experimental Biology,1988,138:181-193.
[19] Kukal O,Dawson T E.Temperature and food quality influences feeding behavior,assimilation efficiency and growth rate of arctic woolly-bear caterpillars[J].Oecologia,1989,79(4):526-532.
[20] Kevan P G,Jensen T S,Shorthouse J D.Body temperatures and behavioral thermoregulation of high arctic woolly-bear caterpillars and pupae(Gynaephorarossii,Lymantriidae:Lepidoptera) and the importance of sunshine[J].Arctic and Alpine Research,1982,14(2):125.
[21] Convey P.How are the life history strategies of antarctic terrestrial invertebrates influenced by extreme environmental conditions[J].Journal of Thermal Biology,1997,22(6):429-440.
[22] Bennett V A,Lee R E,Nauman J S,etal.Selection of overwintering microhabitats used by the arctic woollybear caterpillar,Gynaephoragroenlandica[J].Cryo Letters,2003,24(3):191-200.
[23] Strathdee A T,Bale J S.Life on the edge:Insect ecology in arctic environments[J].Annual Review of Entomology,1998,43:85-106.
[24] 侯建香.草原黑毛虫的生长发育与气象条件的关系[J].青海气象,1999(3):29-30.
[25] 陈安杰.草原毛虫生活习性调查初报[J].四川草原,1984(1):51-56.
[26] 戴虎德,方应春.果洛地区气候变化对草原毛虫影响分析[J].青海气象,2010(2):34-38.
[27] 严林,袁兴宁,鲍学文.青海草原毛虫幼虫的空间分布型及抽样技术研究初报[J].青海畜牧兽医学院学报,1988(1):20-24.
[28] 丁岩钦.昆虫种群生态学原理与应用[M].北京:科学出版社,1980:80-124.
[29] 严林.门源草原毛虫空间分布型及抽样技术研究[J].昆虫知识,1996(3):164-167.
[30] 沈南英,刘伯良,曾璐,等.草原毛虫消长规律及预测预报的研究[J].中国草原,1983(4):57-61.
[31] 侯秀敏,拉毛,潘桂兰.青海省草地虫害调查与预测[J].青海草业,2004(1):49-51.
[32] 严林,兰景华.Nachman模型在门源草原毛虫蛹种群简易估计抽样中的应用[J].青海畜牧兽医学院学报,1994(2):20-23.
[33] 史国菊,吉汉忠.海北州2010年草地虫害危害趋势预测分析[J].青海草业,2010(1):31-32.
[34] 侯秀敏,徐秀霞.青海省草地鼠虫害预测预报工作展望[J].青海草业,2006(3):29-30.
[35] 黄登宇,马恩波.东亚飞蝗Locustamigratoriamanilensis(Meyen)预测预报研究进展[J].动物学报,2001,47(专刊):37-41.
[36] 张勤文,莫重辉,沈明华,等.食入草原毛虫导致放牧羊口腔黏膜溃烂的病理学诊断[J].动物医学进展,2011(12):126-129.
[37] 黎怀鸿.影响草原毛虫活动的主要因素[J].四川草原,1995(4):35-37.
[38] 陈永尧,张生合.杀灭灵灭治草原毛虫及蝗虫的效果[J].养殖与饲料,2008(9):56-59.
[39] 王朝华.杀灭灵、辛硫磷、高效顺反氯氰菊酯防治草原毛虫的药效试验[J].青海畜牧兽医杂志,2000(1):21-22.
[40] 蒋湘,张生合,骆海强.净叶宝、虫必净等五种农药防治草原毛虫药效试验报告[J].青海草业,1999(3):13-14.
[41] 杨帆.9种杀虫剂对草原毛虫的室内效果比较研究[J].青海畜牧兽医杂志,2005(4):5-7.
[42] 邓武烈,陈雨,陈安杰.25%敌百虫油剂防治草原毛虫的药效试验报告[J].四川草原,1982(2):62-66.
[43] 沈世英,韩民新.用敌敌畏等七种农药喷雾防治草原毛虫试验报告[J].畜牧兽医科技通讯,1980(4):5-10.
[44] 任程.锐劲特、敌杀死、快杀灵防治草原毛虫田间试验报告[J].黑龙江畜牧兽医,2003(5):10-12.
[45] 高原.草原毛虫灭治效果试验[J].畜牧兽医科技信息,1997(12):6.
[46] 贺发科,张顺林.溴氰菊酯灭治草原毛虫最佳用药量试验简报[J].青海畜牧兽医杂志,1988(6):44-45.
[47] 沈世英.拟除虫菊酯类杀虫剂对草原毛虫、蝗虫、叶蝉的试验及防治[J].草与畜杂志,1986(3):13-14.
[48] 沈世英,郭树栋,王延宁.杀灭菊酯、除虫菊酯对草原毛虫的毒效测定及防治试验[J].青海畜牧兽医杂志,1986(5):15-19.
[49] 严林,胡凤祖,吴静,等.烈香杜鹃精油和牛尾蒿精油对门源草原毛虫的生物活性[J].西北农业学报,2009(5):58-63.
[50] 王志远,高晓春,袁兴宁,等.病毒杀虫剂防治草原毛虫的应用研究[J].青海草业,1997(2):1-6.
[51] 刘世贵,任大胜,刘德明,等.草原毛虫(GynaephoraruoergensisChou et yin)病毒的发现和鉴定[J].四川大学学报(自然科学版),1984(1):104-108.
[52] 刁治民.核型多角体病毒对草原毛虫的毒力测定[J].草业科学,1996,13(3):50-54.
[53] 刁治民,何长芳.青海草原毛虫核型多角体病毒的初步研究[J].青海畜牧兽医杂志,1993(2):3-6.
[54] 刁治民.草原毛虫病原微生物的初步研究[J].草业科学,1996,13(1):38-40.
[55] 朱文,杨志荣,葛绍荣,等.苏云金杆菌防治草地蝗虫的研究[J].西南农业学报,1995(2):61-64.
[56] 杨志荣,朱文,葛绍荣,等.类产碱假单胞菌防治草地蝗虫的研究[J].中国生物防治,1996(2):55-57.
[57] 刘世贵,任大胜,刘德明,等.草原毛虫核型多角体病毒的首次发现[J].四川草原,1984(2):50-52.
[58] 杨志荣,刘世贵,伍铁桥,等.草原毛虫核型多角体病毒的安全性研究Ⅰ.草原毛虫核型多角体病毒的致病性研究[J].四川大学学报(自然科学版),1990(2):232-238.
[59] 杨志荣,刘世贵,伍铁桥,等.草原毛虫核型多角体病毒的安全性研究Ⅱ.用Ames法检测核型多角体病毒的致突变性[J].四川大学学报(自然科学版),1991,28(4):532-536.
[60] 刁治民,赵风英,刘连荣.青海草原毛虫核型多角体病毒增殖的研究[J].青海师范大学学报(自然科学版),1990(4):32-37.
[61] 刁治民,刘连荣,林建国.草原毛虫核型多角体病毒致死中浓度及致死中时的测定[J].青海畜牧兽医学院学报,1990(Z1):29-33.
[62] 任程.草原毛虫病毒杀虫剂田间药效试验报告[J].青海草业,1996(4):31-32.
[63] 刘世贵,任大胜,杨志荣,等.草原毛虫核型多角体病毒杀虫剂配伍组分筛选[J].四川草原,1988(4):37-42.
[64] 刘世贵,杨志荣,伍铁桥,等.草原毛虫病毒杀虫剂的研制及其大面积应用[J].草业学报,1993,2(4):47-50.
[65] 严林.草原毛虫蛹期寄生天敌种类初步观察[J].青海畜牧兽医杂志,1994,24(6):15-16.
[66] 沈南英,方伋,晋德馨,等.草原毛虫金小蜂生物学特性的初步研究[J].中国草原,1980(1):52-57.
[67] 魏学红,臧建成,马少军,等.西藏那曲地区草原毛虫发生为害情况调查及药剂防治试验[J].中国植保导刊,2009,(11):27-28.
[68] Yan L,Wang G,Liu C Z.Number of instars and stadium duration ofGynaephoramenyuanensis(Lepidoptera:Lymantriidae) from Qinghai-Tibetan Plateau in China[J].Annals of the Entomological Society of America,2006,99(6):1012-1018.
[69] Hebert P D N,Ratnasingham S,de Waard J R.Barcoding animal life:Cytochrome c oxidase subunit 1 divergences among closely related species[J].Proceedings of the Royal Society of London Series B:Biological Sciences,2003,270:96-99.
[70] Hajibabaei M,Janzen D H,Burns J M,etal.DNA barcodes distinguish species of tropical Lepidoptera[J].Proceedings of the National Academy of Sciences of the United States of America,2006,103(4):968-971.
[71] Hebert P D N,Stoeckle M Y,Zemlak T S,etal.Identification of birds through DNA barcodes[J].PLOS Biology,2004,2(10):1657-1663.
[72] Kerr K C R,Stoeckle M Y,Dove C J,etal.Comprehensive DNA barcode coverage of North American birds[J].Molecular Ecology Notes,2007,7(4):535-543.
[73] Tavares E S,Baker A J.Single mitochondrial gene barcodes reliably identify sister-species in diverse clades of birds[J].BMC Evolutionary Biology,2008,8:81
[74] Yoo H S,Eah J Y,Kim J S,etal.DNA barcoding Korean birds[J].Molecules and Cells,2006,22(3):323-327.
[75] Savolainen V,Cowan R S,Vogler A P,etal.Towards writing the encyclopaedia of life:an introduction to DNA barcoding[J].Philosophical Transactions of the Royal Society B:Biological Sciences,2005,360(1462):1805-1811.
[76] Ward R D,Zemlak T S,Innes B H,etal.DNA barcoding Australia’s fish species[J].Philosophical Transactions of the Royal Society B:Biological Sciences,2005,360(1462):1847-1857.
[77] Elsasser S C,Floyd R,Hebert P D N,etal.Species identification of North American guinea worms(Nematoda:Dracunculus) with DNA barcoding[J].Molecular Ecology Resources,2009,9(3):707-712.
[78] Luo Y,Gao W,Gao Y,etal.Mitochondrial genome analysis ofOchotonacurzoniaeand implication of cytochrome c oxidase in hypoxic adaptation[J].Mitochondrion,2008,8(5-6):352-357.
[79] Xu S Q,Yang Y Z,Zhou J,etal.A mitochondrial genome sequence of the Tibetan antelope(Pantholopshodgsonii)[J].Genomics Proteomics Bioinformatics,2005,3(1):5-17.
[80] Elson J,Lightowlers R.Mitochondrial DNA clonality in the dock:Can surveillance swing the case[J].Trends in Genetics,2006,22(11):603-607.
[81] Gissi C,Iannelli F,Pesole G.Evolution of the mitochondrial genome of Metazoa as exemplified by comparison of congeneric species[J].Heredity,2008,101(4):301-320.
[82] Avise J C.Phylogeography:retrospect and prospect[J].Journal of Biogeography,2009,36:3-15.
[83] Boore J L.The use of genome-level characters for phylogenetic reconstruction[J].Trends in Ecology and Evolution,2006,21(8):439-446.
