大通河上游高寒草甸植物群落的退化特征
2013-03-14马玉寿施建军王彦龙李世雄景美玲闵星星
刘 玉,马玉寿,施建军,王彦龙,李世雄,景美玲,闵星星
(1.青海省畜牧兽医科学院,青海 西宁 810016;2.青海大学三江源区高寒草地生态省部共建教育部重点实验室,青海 西宁 810016)
大通河流域发源于祁连山区海西州天峻县境内,东接黄土高原,西为柴达木盆地,南靠湟水谷地,北临河西走廊,由西向东流经天峻、祁连、刚察、门源、互助、天祝、永登、民和等县,与湟水汇合后注入黄河,全长554 km,总流域面积15 133 km2。由于地处青藏高原东北边缘,大通河流域气候恶劣,生态系统脆弱,人为活动对生态环境破坏严重,导致其生态环境急剧恶化,草地退化、水土流失及生物多样性不断减少等现象严重。
草地生态系统是大通河流域的生态主体,高寒草甸是大通河上游地区主要的植被类型之一。高寒草甸生态系统脆弱,抗干扰能力差,植被一旦遭受破坏,靠其自然恢复不但周期长,而且极为困难[1]。自20世纪80年代以来,大通河上游地区的高寒草甸严重退化。由于长期以来缺乏科学管理,传统落后的利用方式,掠夺式经营,加之受过度放牧、鼠虫危害和气候变化的影响,草地毒、杂草大量滋生蔓延,优良牧草比例大幅度下降,导致生产力降低,草地生态环境日趋恶化,水源涵养能力下降,草地生态服务功能丧失,这直接制约着大通河流域的社会经济可持续发展和人民生活水平提高[2-4]。为此,本研究对大通河上游地区不同退化程度高寒草甸植被群落特征进行研究,并分析不同退化程度高寒草甸的群落组成、物种多样性及生产力变化规律,以期为该区生物多样性保护和资源的持续利用提供理论依据。
1 材料与方法
1.1研究区概况 研究区位于青海省祁连县默勒镇瓦日干村的夏季牧场,地理位置37°56′ N,100°13′ E,海拔3 650 m,属高原大陆性气候,冷季长、暖季短,年均温-1.7 ℃,1月平均气温-14.8 ℃,7月平均气温9.8 ℃,气温日差较大,干湿分明,气温和降水垂直变化明显,雨热同期,年平均降水量614.8 mm。该区光能资源丰富,太阳辐射强,年蒸发量平均为1 162.3 mm,草地植被主要以高寒草甸为主,原生植被的主要优势种有垂穗披碱草(Elymusnutans)、紫花针茅(Stipapurpurea)、矮嵩草(Kobresiahumilis)、高山嵩草(K.pygmaea)等,土壤为高寒草甸土[5]。
1.2样地设置与测定方法 于2012年8月在祁连县默勒镇瓦日干村夏季牧场相对平坦的滩地,根据江河源区草地退化4级梯度标准[1]选择未退化、轻度退化、中度退化、重度退化和极度退化的典型代表样地各3个,样地面积为100 m×100 m。在各样地对角线上选取能代表样地的0.5 m×0.5 m的样方10个,测定样方内的植物种类、物种数、群落总盖度、物种分盖度和自然高度。采用刈割法测定群落地上生物量,在实验室烘箱内于70 ℃下烘至恒质量恒定,分别测定干质量,作为地上生物量的指标。
1.3数据计算与分析
1.3.1植物群落重要值
重要值(IV)=(RC+RH+RB)/3。
式中,RC表示相对盖度,RH表示相对高度,RB表示相对生物量。
1.3.2物种多样性指数的测度
α多样性指数:
式中,Pi为种i的相对重要值,S为种i所在样方的物种数。
β多样性指数:
Sorenson指数(βCS) =1-CS。
式中,CS=2j(a+b),j为两个群落或样地共有种数,a和b为样方A和样方B的物种数。
以上数据采用Excel 2010数据处理软件及SPSS 17.0统计分析软件进行数据整理及统计分析。通过单因子方差分析(One-way ANOVA)比较不同退化程度草地间β多样性的变化。
2 结果与分析
2.1不同程度退化草地植物群落组成 根据样方调查数据统计,试验样地中共有草本植物24科51属60多种,其中种类最多的是菊科(Compositae),其次是禾本科(Gramineae)、莎草科(Cyperaceae)、豆科(Leguminosae)、龙胆科(Gentianaceae)、毛茛科(Ranunculaceae)、蔷薇科(Rosaceae)、玄参科(Scrophulariaceae)等。
2.1.1未退化草地 植物生长茂密,群落总盖度为97.8%,由30种植物组成,隶属16科29属(表1)。植物群落以高山嵩草为优势种,优势度为29.89%。次优势种为紫花针茅、垂穗披碱草和冷地早熟禾(Poacrymophila),优势度为21.86%。主要伴生种有黄花棘豆(Oxytropisochrocephala)、雪白委陵菜(Potentillabifurca)、卷鞘鸢尾(Irispotaninii)、洽草(Koeleriacristata)等。以科属的重要值计算,依次为莎草科(31.77)>禾本科(25.13)>菊科(8.48)>豆科(7.31)>蔷薇科(5.77)>鸢尾科(4.35)>龙胆科(4.24)>玄参科(3.91),其余8科9属重要值为9.04。
2.1.2轻度退化草地 群落总盖度为85.5%,由23种植物组成,隶属11科21属。群落优势种为美丽风毛菊(Saussureapulchra),优势度为18.07%。次优势种为洽草、高山嵩草、矮嵩草和垂穗披碱草,优势度为35.68%。主要伴生种为中华羊茅(Festucasinensis)、草地早熟禾(P.pratensis)、西北黄耆(Astragalusfenzelianus)、肉果草(Lanceatibetica)、矮火绒草(Leontopodiumnanum)等。以科属的重要值计算,依次为禾本科(35.22)>菊科(22.33)>莎草科(21.58)>玄参科(5.25)>豆科(5.01),其余6科9属重要值为10.61。
2.1.3中度退化草地 群落总盖度为65.5%,由38种植物组成,隶属15科28属。群落优势种为甘肃棘豆(Oxytropiskansuensis),优势度为9.69%。次优势种为垂穗披碱草、洽草、黄花棘豆和矮火绒草,优势度为31.47%。主要伴生种为冷地早熟禾、高山嵩草、沙生风毛菊(S.arenaria)、鹅绒委陵菜(P.anserina)等。以科属的重要值计算,依次为禾本科(21.43)>豆科(18.95)>菊科(18.29)>龙胆科(15.47)>玄参科(7.65)>蔷薇科(6.92)>莎草科(6.08),其余8科10属重要值为5.21。
2.1.4重度退化草地 群落总盖度为46.0%,由18种植物组成,隶属10科17属。群落优势种为细叶亚菊(Ajaniatenuifolia),优势度为19.11%。次优势种为高山嵩草、矮嵩草、羽叶点地梅(Pomatosacefilicula)、矮火绒草、黄花棘豆和苔草(Carexspp.),优势度为35.56%。主要伴生种为多裂委陵菜(P.multifida)、二裂委陵菜(P.bifurca)、天蓝韭(Alliumcyaneum)、美丽风毛菊、肋柱花(Lomatogoniumcarinthiacum)等。以科属的重要值计算,依次为莎草科(33.20)>菊科(29.46)>报春花科(8.88)>蔷薇科(6.37)>豆科(4.72)>龙胆科(4.70),其余4科4属重要值为12.67。
2.1.5极度退化草地 群落总盖度为36.0%,由19种植物组成,隶属11科17属。群落优势种为细叶亚菊,优势度为34.12%。次优势种为沙生风毛菊、南山蒿(Artemisiananschanica)和二裂委陵菜,甘肃马先蒿(Pediculariskansuensis),优势度为30.46%。主要伴生种为肉果草、甘青大戟(Euphorbiamicractina)、马尿泡(Przewalskiatangutica)等。以科属的重要值计算,依次为菊科(52.63)>蔷薇科(9.68)>禾本科(5.82)>毛茛科(4.99),其余7科6属重要值为16.88。
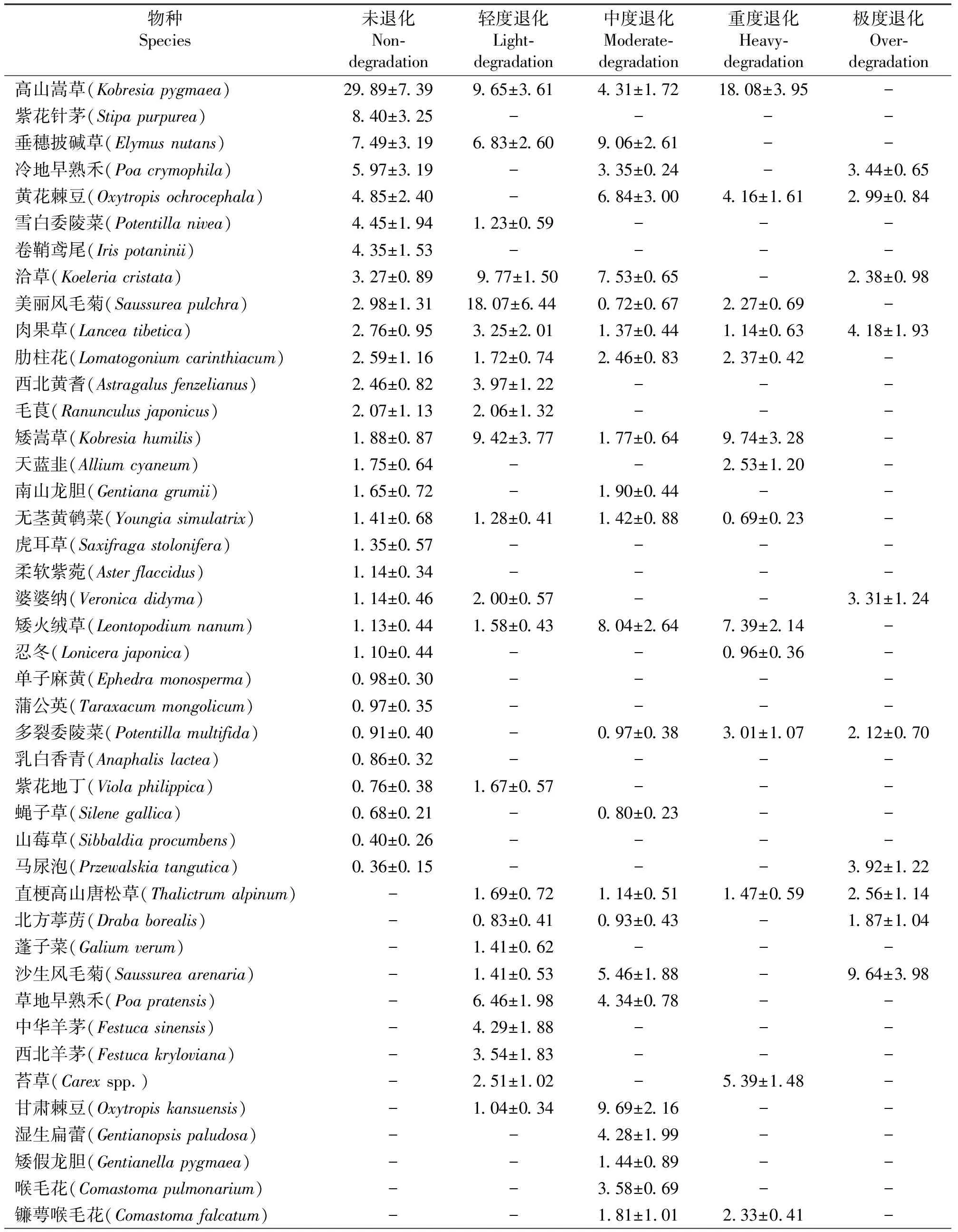
表1 不同退化程度高寒草甸植物群落组成及其重要值Table 1 Community composition and important value of different degraded grasslands
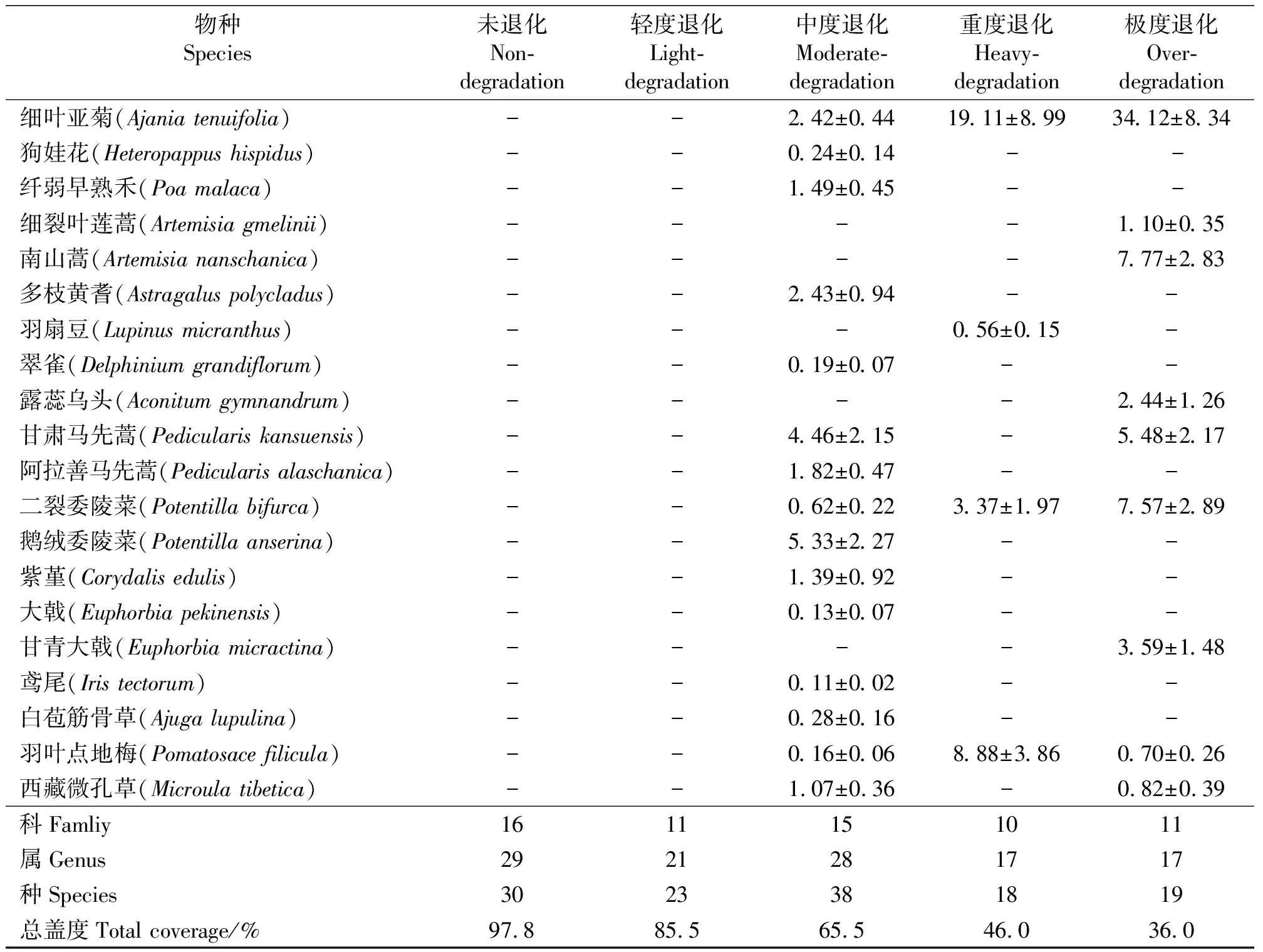
续表1
2.2植物群落物种多样性分析
2.2.1α多样性指数的变化 α多样性是一个测量群落内物种分布数量和均匀程度的指标,是生物群落在组成、结构、功能和动态方面表现出的差异,反映了各物种对环境的适应能力和对资源的利用能力[6]。
中度退化草地的Shannon-Wiener指数最高,显著高于重度退化和极度退化草地(P<0.05);轻度退化、未退化和重度退化草地显著高于极度退化草地(P<0.05),即中度退化>轻度退化>未退化>重度退化>极度退化(图1) 。
从Simpson指数看,不同退化程度草地间的变化趋势与Shannon-Wiener指数相同(图2)。中度退化草地的Simpson指数最高,与重度退化、未退化和极度退化差异显著(P<0.05),轻度退化、重度退化、未退化草地,均与极度退化草地差异显著(P<0.05),即中度退化>轻度退化>重度退化>未退化>极度退化。
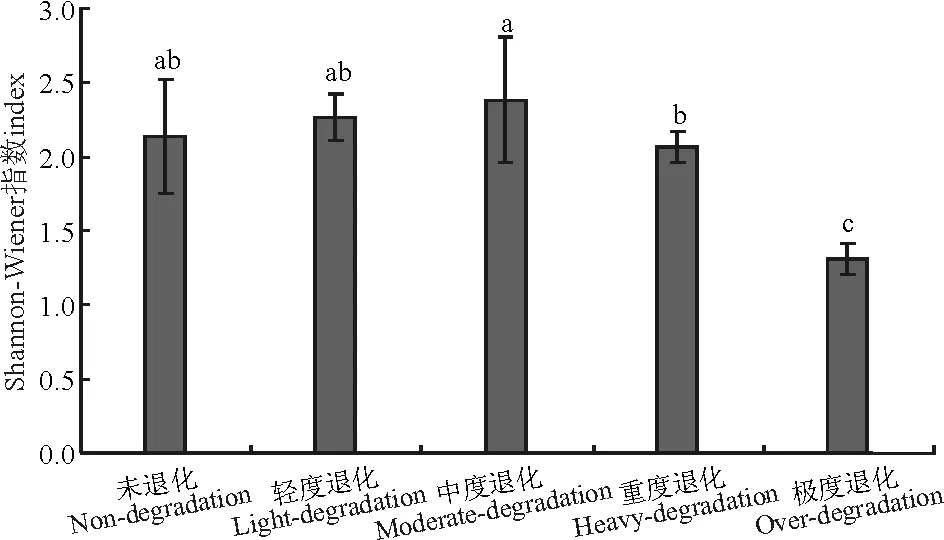
图1 不同退化程度高寒草甸群落Shannon-Wiener指数Fig.1 Shannon-Wiener index of different degraded grasslands communities
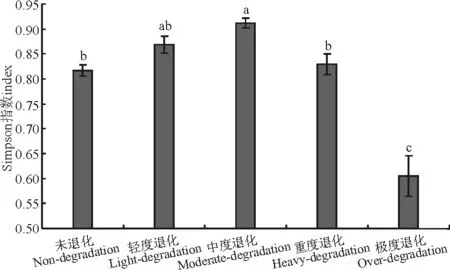
图2 不同退化程度高寒草甸群落Simpson指数Fig.2 Simpson index of different degraded grasslands communities
随退化程度的加剧,均匀度呈现轻度退化>中度退化>重度退化>未退化>极度退化的变化格局(图3),其中轻度退化和中度退化草地显著高于未退化、极度退化草地,重度退化和未退化草地显著高于极度退化草地(P<0.05)。
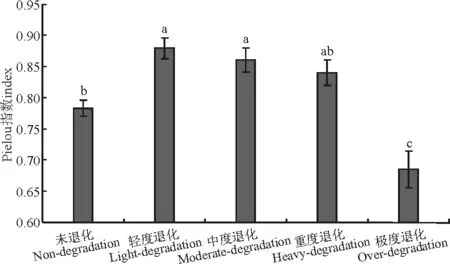
图3 不同退化程度高寒草甸群落Pielou均匀度指数Fig.3 Pielou evenness index of different degraded grasslands communities
2.2.2β多样性指数的变化 β多样性可以定义为沿环境梯度物种的替代程度,或物种周转速率、物种替代速率和生物变化速率等,用以测度群落物种多样性沿着环境梯度的变化速率或群落间的多样性,包括不同群落间物种组成的差异[7]。不同群落或环境梯度上不同点之间共有种越少,即相似性越小,异质程度越大则β多样性值就越高。
未退化-极度退化草地与轻度退化-极度退化草地的β多样性最大,与其它草地间的β多样性差异显著(P<0.05),而未退化-轻度退化草地的β多样性最小(表2)。说明未退化与轻度退化草地群落间的生境条件较为相似,群落的物种组成相似程度较高从而导致β多样性较低,未退化和轻度退化草地与极度退化草地群落间的异质性较大,物种组成改变较多,优势种和次优势种变化明显,群落共有种少,β多样性值较高。
2.3不同退化程度草地的地上生物量分析 随退化程度的加剧,群落总的地上生物量从未退化草地到极度退化草地依次明显降低(图4)。对地上生物量进行方差分析表明,未退化草地生物量最高,与其它退化草地的群落差异显著(P<0.05)。同类植物在不同退化程度草地中生物量差异较大,优良牧草在未退化草地中生物量最高,退化草地相对未退化草地优良牧草的生物量显著降低(P<0.05),极度退化草地的优良牧草生物量最低。可食杂草的生物量依次为轻度退化>中度退化>极度退化>未退化>重度退化。中度退化草地的毒草生物量最大,与未退化及其它退化草地差异显著(P<0.05)。说明随着退化程度的加剧,禾草类、莎草类等优质牧草生物量下降,取而代之的是可食杂草和毒草,毒、杂草生物量明显升高,在群落中占据优势地位,使草地质量下降,而且草地的总生物量不断减少,草地生产力降低。

表2 不同程度退化高寒草甸群落β多样性变化Table 2 β diversity index of different degraded grasslands communities
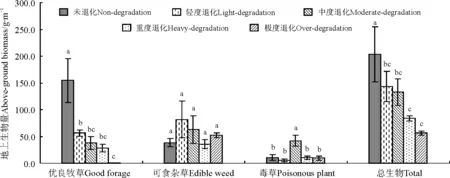
图4 不同退化程度高寒草甸地上生物量变化Fig.4 Above-ground biomass of different degraded grasslands
3 讨论
草地植物群落的结构与组成通常以优势种和种类组成为特征,优势种的更替是植物群落演替的标志。莎草科和禾本科植物是典型高寒草甸的优势种植物,均属于密丛须根型多年生植物,是家畜喜食的优良牧草,同时也是防沙固土的优良生物材料。随着草地利用程度的加重,草地环境恶化,这些植物逐渐退化消失[8]。在未退化草地中由于没有受到放牧的干扰,禾草和莎草植物生长旺盛,在群落中属于竞争优势者;随着草地的退化演替,放牧家畜对禾本科和莎草科植物的采食频率增加,使得他们在群落中的竞争力不断降低,比例急剧下降,莎草科植物甚至消失,最终形成毒、杂草占优势的极度退化草地植被类型——“黑土滩”。
草地群落物种多样性是表述草地生态系统稳定性和持续性的一个指标,对草地物种多样性的研究可以更好地认识草地群落的组成、变化和发展,这对控制和减缓草地退化很有意义[9]。高寒草甸生态系统的退化,不但使群落物种组成发生巨大变化,而且导致物种多样性发生改变,使群落物种数目和种类之间个体分配的均匀性存在差异。在轻度退化和中度退化草地中由于放牧抑制了优势种的竞争能力,使竞争弱势物种的侵入和定居成为可能,群落内物种多样性出现一定程度的增加,导致中度退化和轻度退化草地物种多样性较大。在极度退化草地中由于原有优势种的消失,一些不受家畜采食的毒、杂草成为优势种,而这些毒、杂草在某种意义上受到了一定的“保护”,植物种分布相对单一,从而使多样性指数和均匀度指数降低。
α多样性指数表明了群落多样性量的差异,而β多样性指数表明了群落间多样性的差异,二者相互结合使用才能描绘出群落多样性的真实面目[10]。随着退化程度的加剧,群落物种数出现了变化,不同群落间共有种越少,β多样性就越高[11]。未退化-极度退化,轻度退化-极度退化,退化阶段较长,群落结构和物种组成发生很大改变,其物种的替代率高。由于过度干扰已经引起了生境的破碎化,原有优势种消失,使新的入侵种在群落中占据优势,群落间的共有种少,导致β多样性增加。未退化-轻度退化阶段是相邻阶段,由于受到干扰较小,群落间物种的变化相对较少,群落相似性高,β多样性值小。
草地退化的明显指标是植物群落种类组成和结构变化,杂类草增多以及优良牧草比例减少[12]。由于在长期过牧、鼠害等干扰下,禾草和莎草类功能群的生物量及组成比例下降,可食杂草和毒草类生物量及比例上升[13]。随着草地退化程度的加剧,地上生物量呈现下降趋势,主要是优势种植物及禾草生物量下降引起的[14]。草地顶级群落优势种群的变化随着优势种的衰退和生草层的消失。群落中动物不喜食的可食杂草或有毒植物作为次优势种或伴生种开始发育[15],并逐渐占据主导地位,导致草地质量下降,生产力降低。
4 结论
随着退化程度的加剧,高寒草甸草地植物群落的物种组成发生明显变化,群落优势种由优良牧草逐步过渡为毒、杂草;α多样性指数呈现低-高-低的变化格局,未退化草地与轻度退化草地群落相似性较其它退化草地高;群落中次优势种或伴生种随退化加剧逐步占据优势地位,草地质量降低。
[1] 马玉寿,郎百宁,李青云,等.江河源区高寒草甸退化草地恢复与重建技术研究[J].草业科学,2002,19(9):5-9.
[2] 尚占环,龙瑞军,马玉寿.青藏高原江河源区生态环境安全问题分析与探讨[J].草业科学,2007,24(3):1-7.
[3] 张目,朱国亮.青藏高原高寒草地生态系统严重退化[J].草业科学,2004,21(2):56.
[4] 孙达飞,龙瑞军,蒋文兰,等.三江源区不同鼠洞密度下高寒草甸植物群落生物量和土壤容重特性研究[J].草业学报,2008,17(5):111-116.
[5] 吴玉虎.大通河流域植物区系[J].云南植物研究,2004,26(4):355-372.
[6] 李博,杨持,林鹏.生态学[M].北京:高等教育出版社,2004.
[7] 邱波,任青吉,罗燕江,等.高寒草甸不同生境类型植物群落的α及β多样性研究[J].西北植物学报,2004,24(4):655-661.
[8] 石德军,李希来,杨力军,等.不同退化程度“黑土滩”草地群落特征的变化及恢复对策[J].草业科学,2006,23(7):1-3.
[9] 杨元合,饶胜,胡会峰,等.青藏高原高寒草地植物物种丰富度及其与环境因子和生物量的关系[J].生物多样性,2004,12(1):200-205.
[10] 柳小妮,张德罡,孙九林,等.东祁连山杜鹃灌丛草地物种多样性特征及干扰因子[J].草地学报,2007,15(1):13-19.
[11] 柳小妮,孙九林,张德罡,等.东祁连山不同退化阶段高寒草甸群落结构与植物多样性特征研究[J].草业学报,2008,17(4):1-11.
[12] 李海英,彭红春,王启基.高寒矮嵩草草甸不同退化演替阶段植物群落地上生物量分析[J].草业学报,2004,13(5):26-32.
[13] 陈全功.江河源区草地退化与生态环境的综合治理[J].草业学报,2007,16(1):10-15.
[14] 刘伟,周华坤,周立.不同程度退化草地生物量的分布[J].中国草地学报,2005,27(2):9-15.
[15] 马玉寿,尚占环,施建军,等.黄河源区“黑土滩”退化群落类型多样性及其群落结构研究[J].草业科学,2006,23(12):6-11.
