西藏典型草地地上生物量季节变化特征
2013-03-14普布次仁德吉央宗姬秋梅
除 多,普布次仁,德吉央宗,姬秋梅,唐 洪
(1.中国气象局 成都高原气象研究所拉萨分部,西藏 拉萨 850000; 2.西藏高原大气环境科学研究所,西藏 拉萨 850000;3.西藏自治区畜牧科学研究所,西藏 拉萨 850000)
生物量是草地生态系统获取能量、固定CO2的物质载体,是生态系统结构组建的物质基础[1-3],也是生态系统最基本的数量特征之一[4]。在草地生态系统中,草地生物量是最为活跃的碳库,代表初级生产力的基本水平,决定了草地的载畜能力[5-6]。草地地上生物量的动态变化研究可以为人们了解草地生态系统的物质循环和能量流动提供基本资料。生物量的大小和季节变化直接影响人类对草地资源的利用,是实现草地持续利用和管理的重要理论根据。
近年来,学者们对青藏高原的草地地上生物量季节动态及草地生态系统碳循环开展了大量研究[7-10]。草地是西藏地区分布面积最广的生态系统类型,其天然草地面积为8 106.71×104hm2,占西藏总土地面积的71.15%,是西藏农耕地面积的232.0倍,林地面积的11.4倍,草地在各土地类型面积中所占比例位居全国各省(市、自治区)之首[11]。西藏高原独特的地理位置、复杂的气候类型和多样的环境形成了类型繁多而复杂的草地植被,既有热带、亚热带分布的草地类型,又有温带、亚寒带和寒带分布的草地类型。因此,从西藏高原复杂的草地植被类型中选取具有代表性的草地植被类型,研究其生物生产力的季节性动态变化规律,不仅能揭示高寒草原不同生态系统的结构、功能及生物生产力形成机制等生态过程,有助于了解草地生产力的时空分布特点和利用方式,还对如何有效地开发和利用草地资源,优化草地资源的管理模式和利用方式,进而实现草地资源的可持续发展具有重要的理论和生产指导意义。
本研究采用西藏高原高寒草甸、高寒草原、高寒沼泽化草甸和温性草原4种典型草地类型地上生物量定点观测数据,分析其季节动态变化特征和生长规律,揭示不同草地类型或同一类型不同区域的草地生物量在时间和空间上的差异,旨在为西藏草地资源的合理利用与管理、优化放牧结构和时间以及退化草地的恢复重建等提供基础资料和科学参考依据。
1 材料与方法
1.1采样点概况 草地生物量野外采样点设置在西藏高原中部当雄县、墨竹工卡县和拉萨市周边,共8个采样点。该地区属于高原温带半干旱季风气候区,年均气温1.5~7.8 ℃,气温由南部雅鲁藏布江河谷和拉萨河谷向北部逐渐降低,年降水量340~594 mm,降水由东向西逐渐减少。
8个采样点的草地类型、植被类型、经纬度及海拔等信息详见表1,其中草地类型和植被类型数据源自西藏自治区第一次草地资源普查成果图件。采样点当雄D和日多A属于以高山嵩草(Kobresiapygmaea)为建群种的高寒草甸,其中当雄D位于当雄谷地远离公路和人类活动影响较小的山坡上,而日多A位于其东部170 km处的墨竹工卡县日多乡东面宽阔平坦地段。当雄B和羊八井观测点是以紫花针茅(Stipapurpurea)为建群种的高寒草原,伴生有小莎草(Cyperusmicroiria),都位于当雄谷地,两者相距近80 km,其中当雄B位于当雄县城以西远离公路的地段,羊八井观测点则位于羊八井镇北面远离公路和人类活动影响小的地段,都为天然草原。拉木乡和拉萨观测点位于拉萨河谷南面相对平缓的山麓冲积扇上,属以藏白蒿(Artemisiayounghusbandii)为建群种的西藏高原中部典型温性草原类型。当雄A和日多B属于低地高寒沼泽化草甸,其中,当雄A位于当雄县城北侧500 m处,有围栏网保护,用于春季放牧,建群种为藏北嵩草(K.littledalei);日多B位于当雄A采样点东部128 km处墨竹工卡县境内谷地,没有围栏网保护,建群种为小叶金露梅(Dasiphoraparvifolia),伴有高山嵩草等。
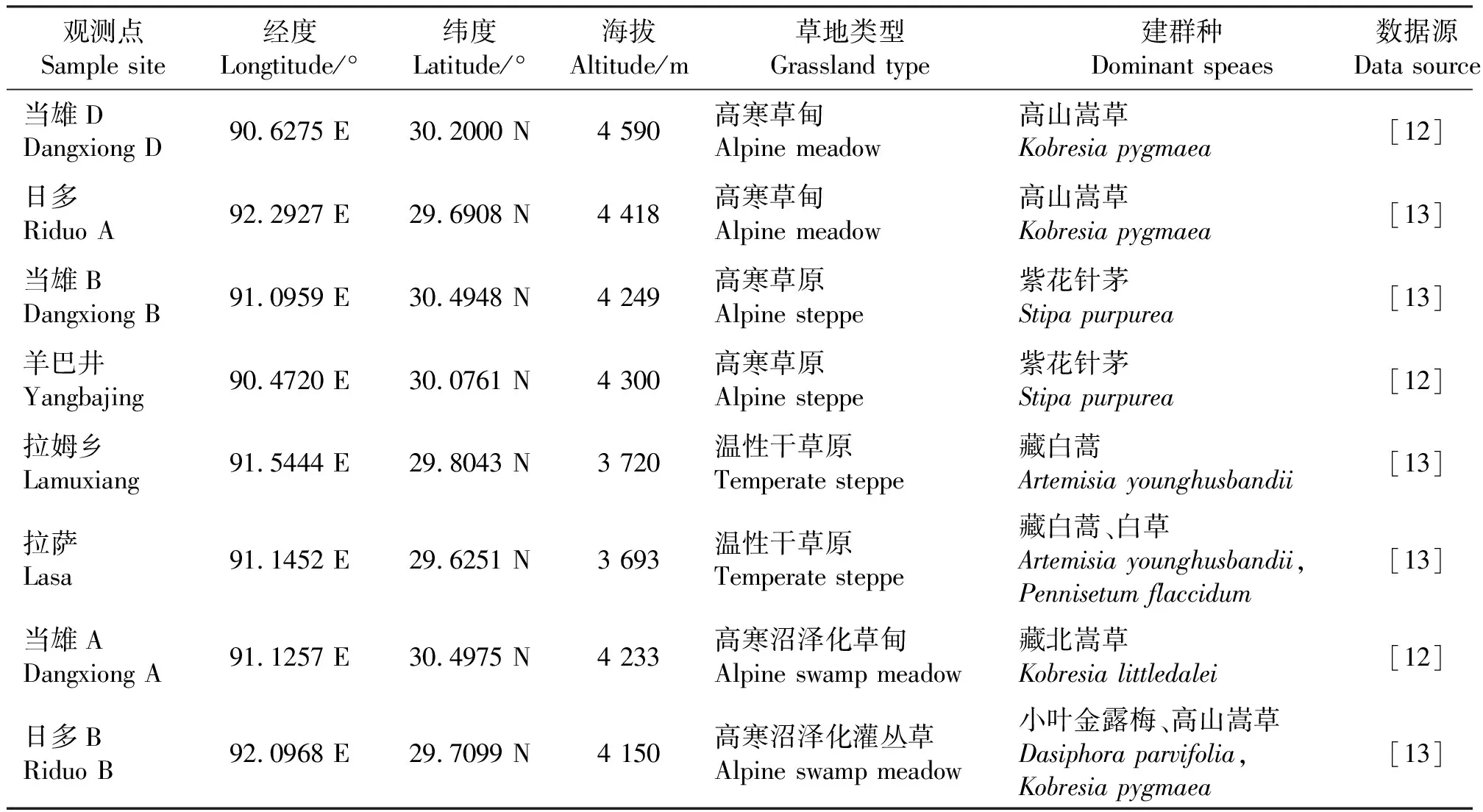
表1 草地生物量采样点信息及数据源Table 1 Grassland, vegetation types, and data sourcesof 8 sampling points
1.2研究方法
1.2.1地上生物量的测定 8个典型草地类型的采样点设在草地植被空间分布比较均一的地方。2004年1-12月分别于每月15日和30日前后3 d内对这8个采样点用收割样方称重法开展了两次的草地地上生物量(Aboveground Biomass,AGB)采样工作,样方面积为50 cm×50 cm,重复3次,同时记录观测点的GPS数据、高程和土地利用类型等。AGB观测步骤:1)面积为50 cm×50 cm正方形线圈;2)用锋利的刀片将3个样方内的草地地上部分齐地面全部刈割;3)除去黏附的土壤、砾石等杂物后装入纸袋全部带回西藏自治区畜牧科学研究所草原实验室晒干;4)在实验室对所有样品的活体和干枯部分(包括立枯物和凋落物)分别进行分拣;5)在85 ℃烘箱中烘干至质量恒定后分别称重,求3个样方内的平均质量。绿色鲜草部分的质量即为草地活体的干物质质量,以下简称活体质量,草地干枯质量包括立枯物和凋落物的质量,以下简称干枯质量,这两个部分的合计值为草地干物质总和,亦即草地地上生物量(AGB),或称为草地总地上生物量。最后都换算成单位面积地上干物质量(g·m-2)。
1.2.2生物量增长率计算 植被的绝对增长速率(Absolute Growth Rate,AGR)和相对增长速率(Relative Growth Rate,RGR)用于分析和解释单位时间内生物量的净积累值。计算公式如下[14-15]:
式中,Bi+1、Bi分别为ti+1、ti时间的地上生物量。
2 分析与结果
2.1高寒草甸地上生物量变化 采样点当雄D和日多A是典型的高寒草甸类草原。当雄D的地上生物量变化特点是1-4月有减小趋势,值也比较小,在18.60 g·m-2以下,且都为干枯草地。年内地上生物量最小值出现在4月,只有9.94 g·m-2。受低温影响,该地区草地的返青期比较晚,从5月初开始有鲜草出现,但其所占份额仍较小,仅为18.83%,5月下旬迅速上升,达64.21%;之后随着植物的生长发育、气温的回升和降水量的增加,草地生物量的累积率迅速增大,草地产量迅速增大,8月地上生物量达到年内最大值,总地上生物量和活体质量分别为49.63和47.74 g·m-2,活体质量成分在96%以上,为年内最大(图1)。从9月开始,随着气温的下降和雨季的结束,地上生物量开始下降,10月活体比重已降至23.14%,之后的11-12月只有干枯物质,一般保持在20 g·m-2左右。该点在7-8月夏季生物量都较大,处于一年中草地产量最高的阶段,产量在44 g·m-2以上,其中活体的比重在94%以上。年均地上生物量为26.24 g·m-2,其中活体和干枯物质各占一半,分别为13.47和13.03 g·m-2(图1,表2)。
同样,作为典型的高寒草甸类草地,日多A月均生物量变化特点为,1-3月和10-12月草地地上生物量都以干枯物质形式存在,其中3月处于年内草地产量最低的阶段,只有 21.77 g·m-2(图2)。随着温度的逐渐上升,4月植被开始返青,出现了活体,但所占比重在6.50%以下。5月上升至14.71%, 6月产量明显增大,活体比重达62.17%,7月达到年内最大值,为66.15 g·m-2,且大多为年内生长的活体部分,比重为92.38%。8月开始产量逐渐下降,但生物量保持在58.00 g·m-2左右,7-8月的活体比重很大,在92%以上。9月开始产量减少显著,但活体的比例还在62%左右,到了10月没有活体部分,仅为干枯物质。年均地上生物量为35.97 g·m-2,其中活体和干枯质量分别为14.16和21.89 g·m-2,干枯草地所占比例相对较大一些,在60%左右(表2、图2)。
由于东西部地区水热条件存在差异,即降水由东向西逐渐减少,温度由南向北逐渐递减,加之西藏的雨季由东南向西北逐步推进的季节格局,同类型草地的地上生物量在东西部存在显著的时空差异。东部墨竹工卡境内的产量明显高于西部当雄地区,西部当雄D的年均地上生物量为东部日多A的73%;东部在7月生物量达到年内最大值,而西部当雄D则在8月达到年内最大值;东部4月草地植被开始返青,西部从5月初开始有活体出现;东部从10月开始植被都已干枯,而西部地区仍有活体;东西部在草地植被的返青和生长季结束的时间上大致存在1个月的差异。在草地植被生长期地上生物量的季节变化均表现为单峰型(表2、图1、图2)。
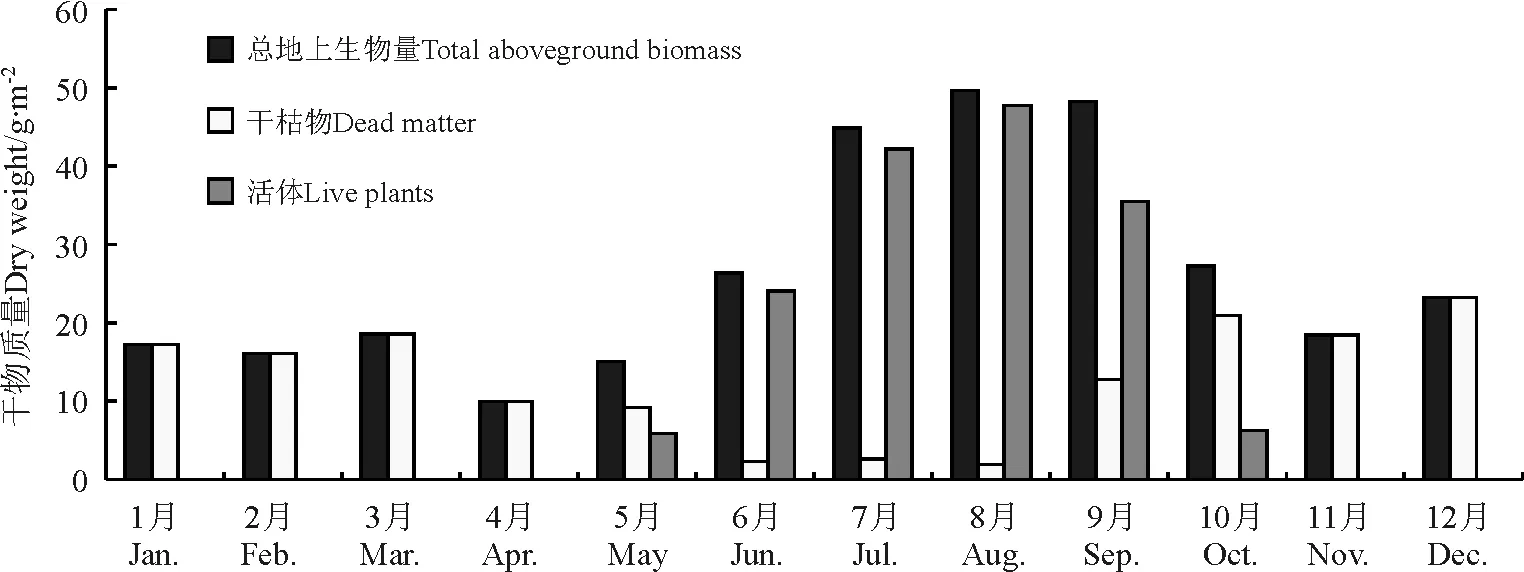
图1 2004年当雄D观测点月均地上总生物量、活体及干枯物干质量变化Fig.1 Monthly mean above ground biomass,live plants and dead dry matter of alpine meadow in Daxiong D sample site in 2004
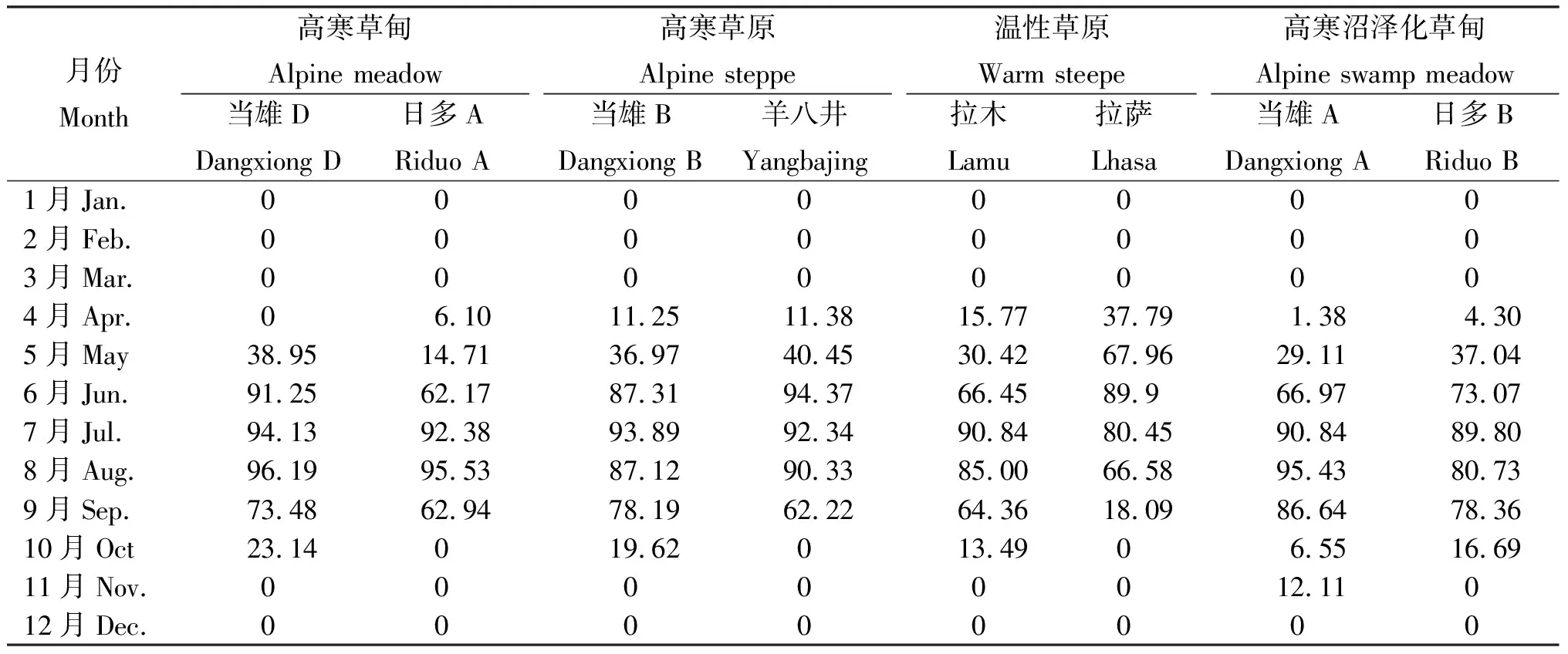
表2 2004年西藏高原典型草地类型月均活体生物量占地上总生物量的比例 Table 2 Live plants biomass to total aboveground biomass ratio of 4 typical grassland types in 8 sample sites
2.2高寒草原地上生物量变化 典型高寒草原草地类型观测点当雄B的地上生物量季节变化特点主要为,春季一般在14.80 g·m-2以下,冬季平均为18.90 g·m-2,且均为干枯草地,夏季最高,平均为42.48 g·m-2,秋季则为27.39 g·m-2。月均地上生物量变化特点是,3月最低,只有9.94 g·m-2,随着气温的上升开始返青,4月有活体但所占的比重小于11.25%。此后,随着气温的进一步上升,植被光合作用增强,草地生物量累积迅速,主要体现为活体占总生物量的比例逐渐增大,6-8月活体比重在87%以上,其中7月比重最大,为93.89%,8月地上生物量达到年内最大值,为51.10 g·m-2。10月开始随着气温的下降和雨季的结束,草地植物群落叶片开始枯黄,光合作用减弱,植物体逐渐衰老,枯落量增加,营养物质不断流失并向地下根系转移,导致地上生物量下降趋势显著,从7-9月的48 g·m-2以上下降至10月的25.69 g·m-2,且干枯物比重上升至80%以上,11-12月只有干枯部分,在24 g·m-2以下。年均地上生物量为25.18 g·m-2,其中活体和干枯物分别占54.33%和45.67%。在4-10月,植被生长期总地上生物量和活体生物量的年内变化均表现为单峰型(表2、图3)。

图2 2004年日多A观测点月均地上总生物量、活体及干枯物干质量变化Fig.2 Monthly mean above ground biomass,live plants and dead dry matter of alpine meadow in Riduo A sample site in 2004
同样为高寒草原类型的羊八井观测点的地上生物量季节变化特点是,春季生物量最低,为19.30 g·m-2,其次为冬季(19.89 g·m-2)和秋季(31.90 g·m-2),夏季草地的产量最大,达42.1 g·m-2,年平均生物量为28.30 g·m-2,其中鲜草和干枯物分别为13.01和15.29 g·m-2。月均变化特点是,1-3月地上生物量在18 g·m-2以下,其中1月为年内最低,仅为5.4 g·m-2,4月随气温的上升开始返青,但活体比重很小,仅为11.39%,5月上升至40.45%,6-8月夏季3个月在90%以上。其中,8月达到48.90 g·m-2,其鲜草也最大,为44.17 g·m-2,9月开始进入秋季后,随着气温的下降和雨季的结束,草地的光合作用减弱,草地开始枯黄,地上生物量呈下降态势,特别是活体部分的比重减少趋势尤为明显,10-2月仅为残留的干枯物,草地生长期的4-9月,地上生物量和活体生物量的变化都表现为单峰型(表2、图4)。
从以上两个高寒草原类型的年内变化特征来看,植被生长季节4-10月二者地上生物量和活体生物量变化趋势基本一致。主要体现在:植被从4月开始返青但活体比重较小,一般在11%左右,两个观测点都在8月达到生物量的最大值,且大小基本一致,在48~52 g·m-2,相差很小;7-9月3个月的生物量都保持相对较高的稳定状态,在46~52 g·m-2,年均生物量也都在25.18~28.30 g·m-2,相差不大;10月生物量开始下降,特别是活体比例下降尤为突出,当雄B点在20%以下,而羊八井观测点则没有活体,可能是由于该观测点海拔相对较高、地处风口且气温下降显著导致植被提前枯黄(表2、图3、图4)。
2.3温性草原地上生物量变化 与高寒草甸和草原相比,西藏中部典型温性草原草地的拉木乡观测点平均生物量较大,为46.88 g·m-2,月平均地上生物量都大于21 g·m-2,最大达71.40 g·m-2,其中,春季产量相对较低,为32.83 g·m-2,其次为秋季,47.37 g·m-2,夏季和冬季相差不大,都在53 g·m-2左右。月平均生物量变化特点是:5月生物量最低,为21.63 g·m-2,其次为4月,31.08 g·m-2;4月开始随着草地返青和生物量的累积,产量逐渐上升,特别是6月之后生物量增加尤为显著,直至8月草地产量达到71.40 g·m-2的年内月均最大值,之后进入了生物量下降阶段,但是至10月下降不显著,仍为56.56 g·m-2,但86.51%已为干枯物质形式存在。从活体生物量变化特点来看,4月开始就有活体,但所占比重较小,为15.77%,此后增加迅速,5月为30.42%,6-9月都在64%以上,其中7月活体比重达到年内最大值,为91%,10月下降至13.49%,11-3月只有干枯草地。相对于前面的高寒草甸还是高寒草原类草地,由于该采样点地处相对温暖的拉萨河谷地,草地生长期比前两者相对长一些,为4-10月(表2、图5)。
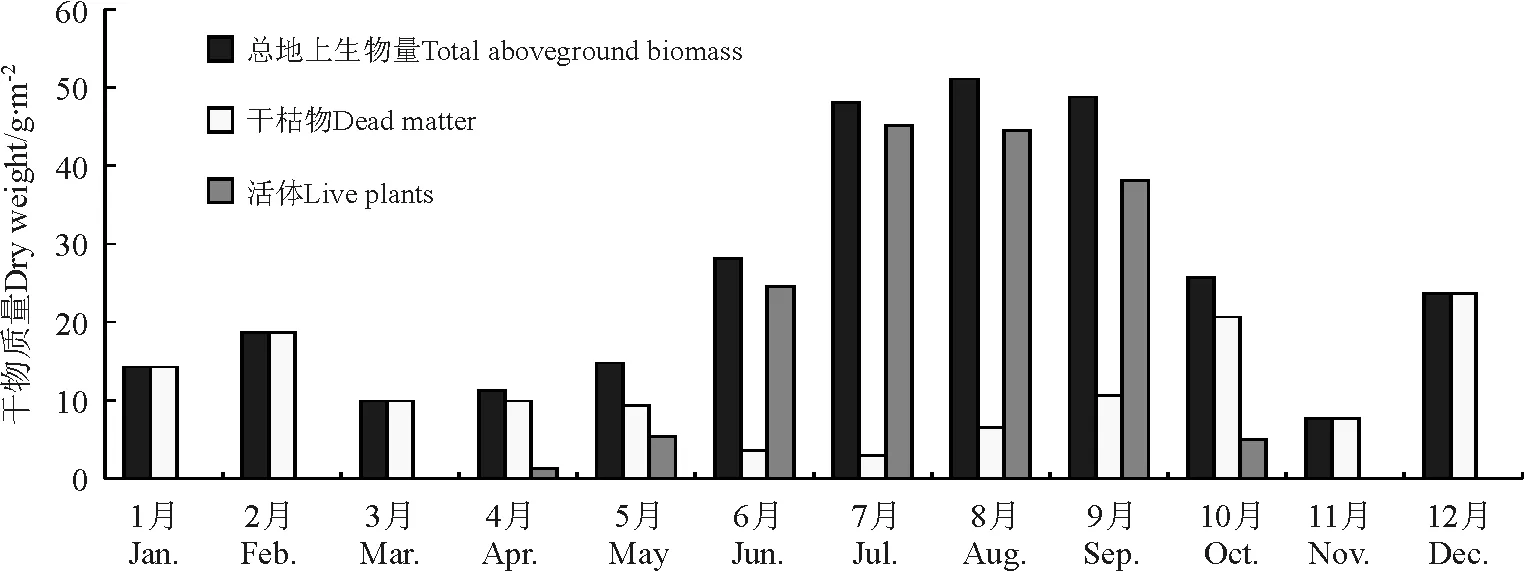
图3 2004年高寒草原草地类型当雄B观测点月均地上总生物量、活体及干枯物干质量变化Fig.3 Monthly mean above ground biomass,live plants and dead dry matter of alpine steppe in Dangxiong B sample site in 2004
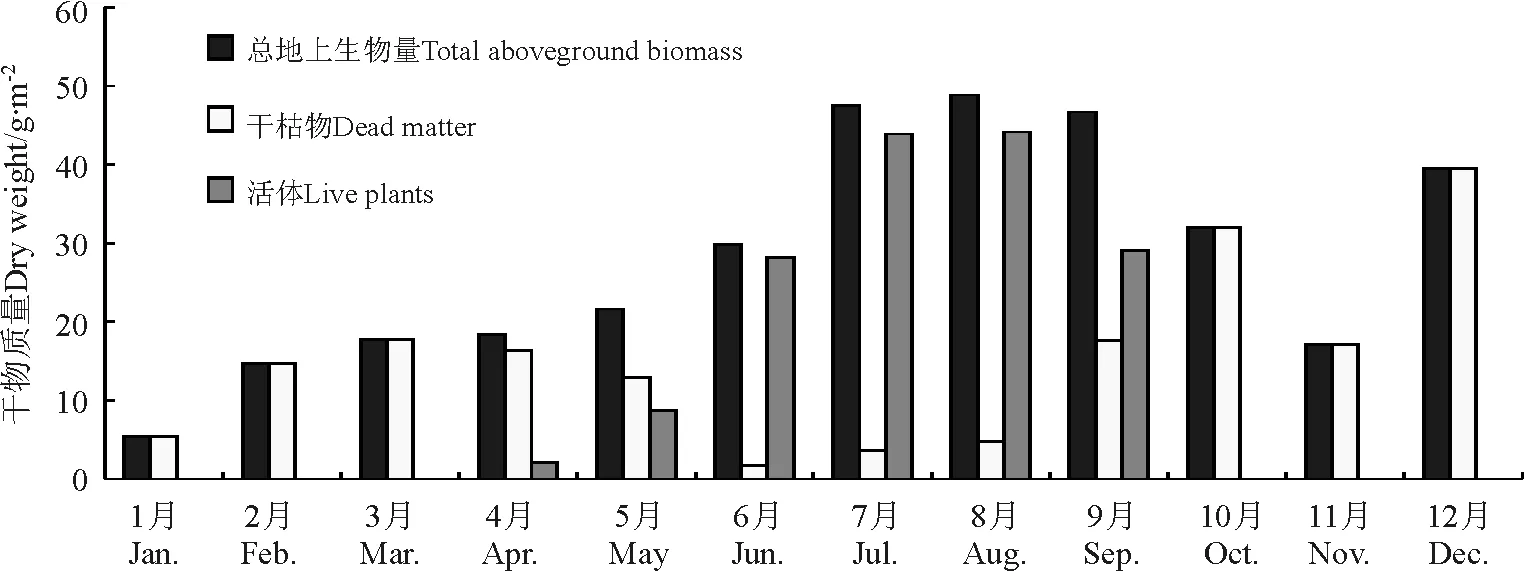
图4 2004年高寒草原草地类型羊八井观测点月均地上总生物量、活体及干枯物干质量变化Fig.4 Monthly mean above ground biomass,live plants and dead dry matter of alpine steppe in Yangbajing sample site in 2004
拉萨观测点也属于典型的西藏中部河谷温性草原草地,这里的平均产草量较高,达80.56 g·m-2。其显著季节性特点是,4个季节的草地产量基本保持在相对较高的状态,都在71 g·m-2以上,没有明显的季节性变化特点。秋季的产量相对较大,达94.6 g·m-2,夏季和冬季基本在78 g·m-2左右。该观测点位于拉萨市南面山地洪冲积扇上,对草地的利用强度较其它农牧区小,因而草地的产量相对保持在较高的水平,几乎为同类型草地拉木乡观测点的1倍左右。月均生物量变化不存在显著的月际波动,11月和5-6月产量相对较低,在51~60 g·m-2,其它月份都在72 g·m-2以上,10月出现最大值,达129.64 g·m-2。由于草地样品采样时的随机性和温性草原草地的局地异质性较其它天然草地显著,所以观测值无法非常准确地反映本草地类型的月均变化,但是观测值体现了该地段产量较高,草地季节性和月际波动小的特点。月均活体生物量变化特点主要表现在,5月初随着植被返青进入了草地生长阶段,但此时活体比重不到一半,为37.79%,6月开始由于气温的明显上升,植被光合作用强烈,使得生物量累积迅速,这一比例至9月都在66%以上,其中7月的活体比重达到年内月均最大值,为90.00%,其次为8月80.44%,10月已下降到18.09%,2-4月和11-12月没有活体,仅为干枯草地(图6)。对该点的野外采样工作开始于2004年2月11日,所以没有1月的数据供分析。
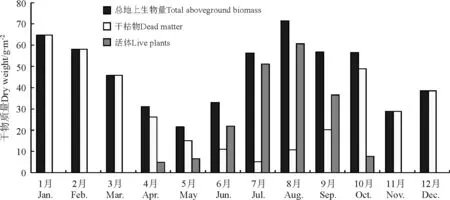
图5 2004年拉木乡观测点月均地上总生物量、活体及干枯物干质量变化Fig.5 Monthly mean above ground biomass,live plants and dead dry matter of temperate grassland in Lamu sample site in 2004
虽然上述两个采样点同属温性草原草地类,且都处在拉萨河南岸山地山麓冲积扇和缓坡上,但是观测结果存在以下的差异:拉萨观测点的年平均生物量明显大于拉木乡观测点,两者相差1倍左右,而且拉萨观测点没有明显的季节性变化,且基本保持在71 g·m-2以上。可见,城市化进程使得拉萨市周边山地草地资源的利用强度低,草地生物量的季节变化不明显,而远离拉萨市的拉姆乡附近对草地资源的利用强度比城市周边高,放牧食草导致了该地区草地产量较低,且出现了明显的季节变化。植被长期地上生物量和活体生物量变化都表现为单峰型(表2、图5、图6)。

图6 2004年拉萨观测点月均地上总生物量、活体及干枯物干质量变化Fig.6 Monthly mean above ground biomass,live plants and dead dry matter of temperate grassland in Lhasa sample site in 2004
2.4高寒沼泽化草甸地上生物量变化 当雄A位于当雄县城北侧,属于低地高寒沼泽化草甸,有围栏网保护,用于春季放牧。该观测点的平均生物量达384.45 g·m-2,在所有观测点中产量最高。从季节变化来看,春季因放牧生物量最低,为191.36 g·m-2,冬季次之,为375.81 g·m-2,夏季达406.28 g·m-2,草地产量最大值出现在秋季,达564.38 g·m-2。月均生物量变化特点为,从1月开始逐渐降低,4月达到年内最低,为182.14 g·m-2,5月比4月略高,为 182.77 g·m-2,6月开始,随着气温的上升,光合作用显著增加,绿色植物生长强劲,产量呈现显著上升态势,直至9月草地产量达到年内最大值,为711.52 g·m-2。10月开始逐渐减少,直至春季达到最低值。活体生物量月均变化特点为,4月开始出现活体,但其比重仅为1.38%,5月上升至30%,6月为66.97%,7-9月在86%以上,其中8月这一比重最大,达95.43%。10月活体出现了锐减现象,仅为6.55%,而1-3月和12月没有活体,仅为残存的干枯草地(表2、图7)。
日多B观测点位于研究区东部墨竹工卡县境内谷地,属于低地高寒沼泽化草甸,没有围栏网保护。由于其丰富的地下水资源,草地生物量较大,达222.35 g·m-2。草地地上生物量季节变化主要表现在,春季最低,为188.67 g·m-2,其次为冬季(213.24 g·m-2),最大值出现在夏季,达256.71 g·m-2,秋季一般在230.76 g·m-2。月均变化的特点是月际波动较小。4月的地上生物量较低,为167.79 g·m-2,其次为3月(168.63 g·m-2),其余月份都在174.58 g·m-2以上,其中6-9月均在244.00 g·m-2,9月达到了月均最大产量,为297.12 g·m-2。活体生物量月均变化特点为4月返青后,植被生长开始,活体比重仅为4.30%,之后温度的上升使得光合作用加强,生物量累积迅速,活体增加显著,5月达到37.04%,6-9月在73%以上,其中7月达到近90%的月均最大比重。10月开始,随着气温的下降和雨季的结束,植物体衰老,植物叶片枯黄,枯落量明显增加,导致活体比重下降极为迅速,只有16.69%。1-3月及11-12月只有干枯部分(表2、图8)。
由于地理位置低洼、排水不畅、常年积水及土壤湿度常年处于饱和或超饱和状态,使得低地高寒沼泽化草甸地上生物量都很高。其中有围栏网的当雄观测点年均地上生物量达384.45 g·m-2,为温性草原草地的6倍,高寒草甸和高寒草原草地的12~14倍。无围网的日多B观测点年均地上生物量为222.35 g·m-2,比有网围的明显低,仅为其58%左右,但是与其它类型草地相比高出许多,是温性草原的3.5倍,高寒草甸和高寒草原生物量的6~8倍。可见,在相近气候和环境条件下,局地土壤水分含量的大小直接决定了草地地上生物量的大小。与自由放牧相比,围栏禁牧措施可以明显提高草原地上生物量,是改良退化草地最普遍和有效的措施之一(表2、图1-8)。

图7 2004年当雄A观测点月均地上总生物量、活体及干枯物干质量变化Fig.7 Monthly mean above ground biomass,live plants and dead dry matter of swamp meadow in Dangxiong A sample site in 2004
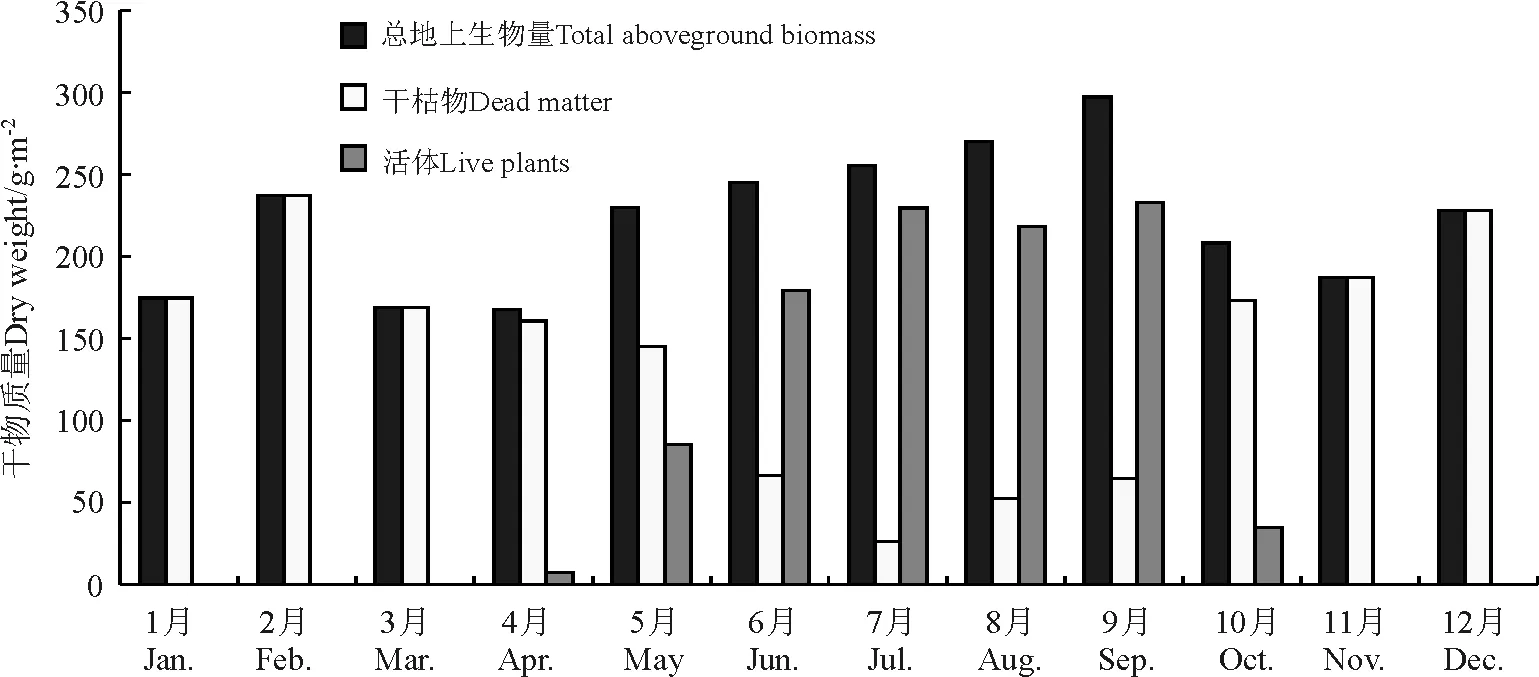
图8 2004年日多B观测点月均地上总生物量、活体及干枯物干质量变化Fig.8 Monthly mean above ground biomass,live plants and dead dry matter of swamp meadow in Riduo B sample site in 2004
2.5地上生物量的增长规律 本研究中的地上生物量不仅包括年内生长的绿色鲜草部分,还包括立枯物和凋落物等干枯部分,其中绿色鲜草部分是年内植被生长期通过植物光合作用所产生的生物量积累部分,呈典型的单峰型。因此,下面分析中仅考虑了活体生物量的绝对增长率(AGR)和相对增长率(RGR)。
不同草地类型地上生物量的活体部分AGR和RGR季节动态均在生物量达到高峰期前是正增长,之后为负增长,且同一观测点的AGR和RGR在季节动态变化趋势上大致相同,但是不同草地类型或同一类型不同区域的草地最快生长期出现的时间存在差异(表3)。
对典型的高寒草甸草原类型来讲,当雄D在4-8月为草地地上生物量累积阶段,累积率在0.184~0.608 g·m-2·d-1,其中,5-6月和6-7月生物量累积率基本一致,达年内达到最大,大约在0.6 g·m-2·d-1,8月之后AGR和RGR都呈现负值。虽属同一植被类型,但由于东西两地水热条件的差异,日多A草地活体生物量AGR的最大值出现在6-7月,日增加率是1.381 g·m-2,日增长率也较当雄D要高出1倍多,其次是5-6月,为0.516 g·m-2·d-1,但增长率出现负值的时间要比当雄D早,表明东部草地提前进入成熟和枯黄阶段(表3)。

表3 2004年草地生长季活体干物质量增长速率动态变化Table 3 AGR and RGR of live plants dry matter in 8 sample sites during the growing seasons in 2004
高寒草原草地类型当雄B的AGR在0.140~0.686 g·m-2·d-1,其中6-7月的日增长率最大,达0.686 g·m-2,RGR最大也出现这一时段,7月鲜草产量达到峰值后植被净累积率出现减少趋势;而羊八井观测点活体生物量的AGR和RGR,在5-6月出现了最大值,分别为0.646 g·m-2·d-1和0.099 g·d-1·d-1,最低出现在7-8月,8月达到最大值后,生物量累积率开始出现减少趋势。位于拉萨河谷地冲积扇上的温性草原的最大累积速率在6-7月,其中拉木乡观测的日累积率为0.973 g·m-2,而拉萨观测点的累积率大于同类型拉木观测点,为1.222 g·m-2·d-1(表3)。
低地高寒沼泽化草甸的AGR、RGR和日累积率都较其它类型大,其中当雄A在植被生长阶段日累积率最大出现在6-7月,高达10.395 g·m-2,其它月份差异不大,一般在1.6~2.7 g·m-2·d-1,9月活体生物量达到最大值,之后的9-10月出现了19.345 g·m-2的减少速率。日多B的AGR峰值要比当雄A提前1月,出现在5-6月,且大小远小于当雄A,为3.131 g·m-2·d-1,9月活体生物量达到峰值后,随着气温的下降和植物的衰老、枯黄,10月绝对增长速率AGR出现负值,且其值达-6.603 g·m-2·d-1(表3)。
2.6草地地上生物量与气候要素的关系 影响草地植被生长的气候因子是多方面的,其中水热条件是影响植被生长和生物量累积的主要因素。为了分析研究区不同草地类型地上生物量与水热条件之间的关系,分别建立了以地上总生物量和活体生物量作为因变量,降水与气温作为自变量的线性回归模型。总体上,草地地上生物量与降水之间显著相关,除草地空间异质性强的温性草原草地拉木乡、拉萨及有人工围网的低地沼泽化草甸当雄A观测点之外,都与降水极显著相关(P<0.01),与气温的关系显著(P<0.05)。对于所有草地类型来说,草地植被生长季节植物光合作用所产生的生物量积累部分即活体生物量与降水之间的线性相关程度比总地上生物量更为密切(P<0.001)。此时,温度与鲜草生物量之间的线性关系要明显高于与总地上生物量之间(P<0.01)的相关程度(表4)。可见,作为温带半干旱季风气候区,降水是影响研究区草地地上生物量的主要气候因素,而与温度关系较弱;活体生物量作为植被生长期通过植物光合作用所产生的生物量积累部分共同受到降水和气温的影响,其中降水是最主要的限制因子。其它研究表明,气候是草地地上生物量的重要影响因素,我国和北美、南美等温带草地地上生物量的变化主要受降水控制,在我国北方,降水是温带草地生态系统生产力(地上生物量) 的主要限制因子之一[16]。
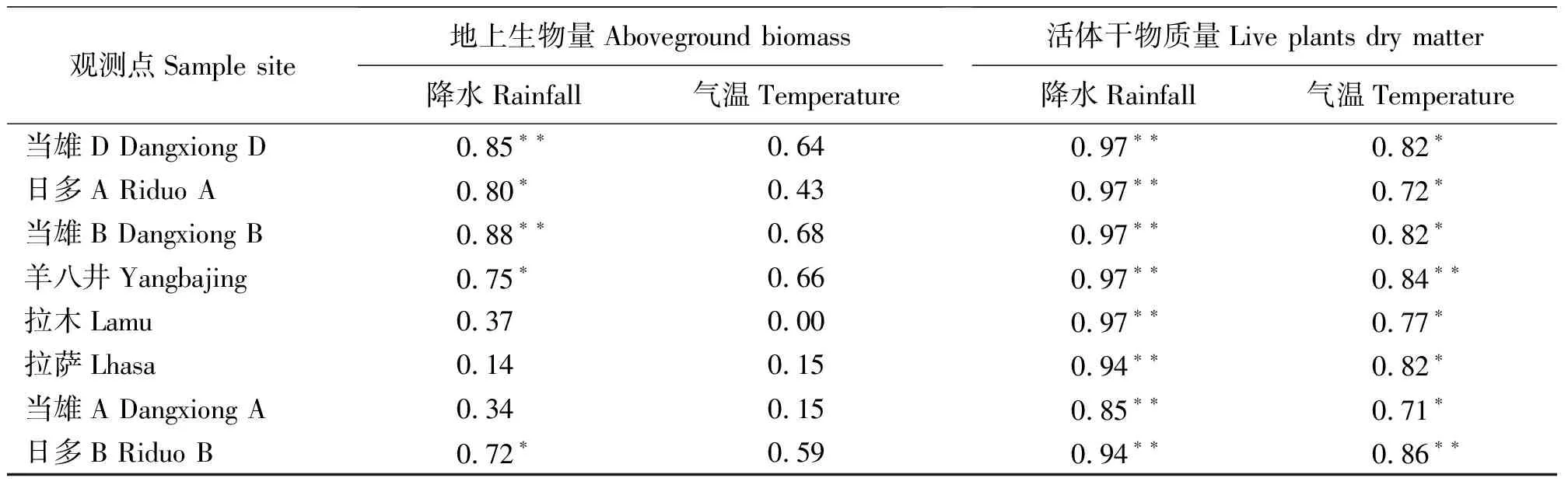
表4 草地地上生物量与气候要素之间的线性相关系数Table 4 The correlation between aboveground biomass and climate variables
3 结论
低地高寒沼泽化草甸地上生物量最高,其中有围网的年均生物量达384.45 g·m-2,为温性草原类草地生物量的6倍,高寒草甸和草原类草地的11~14倍,无围网的地段年均地上生物量为222.35 g·m-2,比有网围的明显低,仅为其58%。与自由放牧相比,围栏禁牧措施可以明显提高草原地上生物量,是改良退化草地最普遍和有效的措施之一。
温性草原是在半干旱气候条件下西藏高原中部雅鲁藏部江及其支流拉萨河和年楚河河谷的典型草地类型,其草地生产力大于高寒草原和高寒草甸。拉萨河谷地两个典型温性草原观测点的草地生物量大小对比表明,城市周边山地草地生物量显著大于远离城市的地区,城市化进程使得城市周边山地草地的放牧强度降低,除了温带半干旱气候区草地所固有的植被生长季节变化之外,草地的总地上生物量不存在明显的季节变化。在远离城市的地区,放牧等人类活动对草地资源的利用强度高,特别是冬春放牧啃食使得该地区草地总产量较低,且出现明显的季节变化。
无论是高寒草甸、高寒草原、高寒沼泽化草甸还是温性草原,草地地上生物量的AGR 和RGR 季节动态均在生物量达到高峰期前是正增长,之后为负增长,并且同一观测点的AGR和RGR在季节动态变化趋势上大致相同,但是不同草地类型或同一类型不同区域的草地最快生长期出现的时间存在先后,主要取决于区域水热条件差异及其季节性变化特征。
降水是影响研究区草地地上生物量主要的气候因素之一,在草地植被生长期光合作用而产生的鲜草生物量累积共同受到降水和气温的影响,其中降水是最主要的限制因子。
[1] Ni J.Carbon storage in grasslands of China[J].Journal of Arid Environments,2002,50:205-218.
[2] Scurlock J M O,Johnson K,Olson R J.Estimating net primary productivity from grassland biomass dynamics measurements[J].Global Change Biology,2002,8(8):736-753.
[3] 高添,徐斌,杨秀春,等.青藏高原草地生态系统生物量碳库研究进展[J].地理科学进展,2012,31(12):1724-1731.
[4] 李凯辉,王万林,胡玉昆,等.不同海拔梯度高寒草地地下生物量与环境因子的关系[J].应用生态学报, 2008,19(11):2364-2368.
[5] Soussana J F,Loiseau P,Vuichard N,etal.Carbon cycling and sequestration opportunities in temperate grasslands[J].Soil Use and Management,2004,20(2):219-230.
[6] 赵同谦,欧阳志云,贾良清,等.中国草地生态系统服务功能间接价值评价[J].生态学报,2004,24(6):1101-1110.
[7] 朱宝文,周华坤,徐有绪,等.青海湖北岸草甸草原牧草生物量季节动态研究[J].草业科学,2008,25(12):62-65.
[8] 梁天刚,崔霞,冯琦胜,等.2001-2008年甘南牧区草地地上生物量与载畜量遥感动态监测[J].草业学报,2009,18(6):12-22.
[9] 乔春连,李婧梅,王基恒,等.青藏高原高寒草甸生态系统CO2通量研究进展[J].草业科学,2012,29(2):204-210.
[10] 杨兆平,欧阳华,宋明华,等.青藏高原多年冻土区高寒植被物种多样性和地上生物量[J].生态学杂志,2010,29(4):617-623.
[11] 西藏自治区土地管理局,西藏自治区畜牧局.西藏自治区草地资源[M].北京:科学出版社,1994.
[12] 西藏自治区土地管理局和畜牧局编制.1∶200万西藏自治区草地类型图[Z].1991.
[13] 西藏自治区畜牧局和西藏自治区土地管理局编制.1∶20万西藏自治区一江两河中部地区草地类型图[Z].1991.
[14] 姜恕.草地生态研究方法[M].北京:农业出版社,1988:67-83.
[15] 杨永兴,王世岩,何太蓉,等.三江平原典型湿地生态系统生物量及其季节动态研究[J].中国草地,2002,24(1):1-7.
[16] 马文红,杨元合,贺金生,等.内蒙古温带草地生物量及其与环境因子的关系[J].中国科学C辑:生命科学,2008,38(1):84-92.
